Состояние популяции Juniperus communis subsp. oblonga bieb. в зависимости от распространения полупаразита Arceuthobium oxycedri (DC.) Bieb. и совместного произрастания c Pinus kochiana Klotzsch в условиях Внутреннегорного Дагестана
 Bieb. и совместного произрастания c Pinus kochiana Klotzsch в условиях Внутреннегорного Дагестана")
Автор: Асадулаев Загирбег Магомедович, Маллалиев Максим Маллалиевич, Садыкова Гульнара Алиловна, Рамазанова Зулфира Рамазановна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Общая биология
Статья в выпуске: 2-3 т.19, 2017 года.
Бесплатный доступ
Обсуждаются причины ухудшения состояния популяции Juniperus communis subsp. oblonga на оползневом склоне хребта Чакулабек Внутреннегорного Дагестана в связи с массовым распространением полупаразита Arceuthobium oxicedri и Pinus кochiana. Показано, что A. oxicedri не является первоначальной причиной деградации популяции J. communis subsp. oblonga. Выдвинуто предположение о том, что сообщества с доминированием J. communis subsp. oblonga на северных склонах гор в аридных условиях Внутреннегорного Дагестана сформировались после вырубки здесь сосновых лесов, которые в настоящее время восстанавливаются.
Полупаразит, внутреннегорный дагестан, куст, произрастание, экотоп, популяция
Короткий адрес: https://sciup.org/148205154
IDR: 148205154 | УДК: 58.02:58.072
State of the Juniperus oblonga bieb. Population depending on the spread semiparasite arceuthobium oxycedri (DC.) bieb. and joint growth with Pinus kochiana Klotzsch under the conditions of Innermountain Dagestan
The reasons of deterioration the state of Juniperus communis subsp. oblonga population on the degraded slope ridge in Chakulabeck at Intermountain Dagestan in connection with the mass spread of semiparasite Arceuthobium polyparasitic oxicedri and the introduction into the community the new dominant species Pinus кochiana were dicussed. It is shown that A. oxicedri is not the primary reason of degradation of J. communis subsp. oblonga population. It was suggested that communities with dominance of J. communis subsp. oblonga on the Northern slopes of mountains in arid conditions of Intermountain Dagestan was formed after the felling of the pine forest and is currently replaced by secondary pine forests.
Текст научной статьи Состояние популяции Juniperus communis subsp. oblonga bieb. в зависимости от распространения полупаразита Arceuthobium oxycedri (DC.) Bieb. и совместного произрастания c Pinus kochiana Klotzsch в условиях Внутреннегорного Дагестана
поражены арцеутобиумом меньше [7]. В мировой литературе распространению и вредоносности A. oxicedri уделено достаточно большое внимание. Прежде всего, это фундаментальная сводка Ciesla et al., [8], где приводится информация об обширной географии этого вида: Африка, Западная Европа, Балканы, Россия и другие бывшие советские республики, Ближний Восток, Индия и западный Китай. A. оxycedri паразитирует здесь на 17 видах можжевельника, 5 видах кипариса, плосковеточнике, кипарисовике [9] и признан потенциально опасным полупаразитным растением [1, 2], который недостающие вещества компенсирует за счет растения-хозяина [3].
Вредоносность A. oxycedri на видах Juniperus обстоятельно изучена во флоре арчовников в бассейне р. Искандер в условиях Таджикистана [10, 11], в заповедном природном комплексе Чилтер в западном Крыму и на мысе Мартьян в южном Крыму ([12]. В арчовых лесах провинции Белуджистан Пакистана [13] проведено полномасштабное изучение биологии этого паразита для разработки лесозащитных мероприятий. Здесь же изучено не только распространение и способность A. оxycedri поражать представителей семейства Cupressaceae , но и детально раскрыты вопросы его репродуктивной биологии [14]. В условиях севера Ирана A. oxycedri также признан опасным полупаразитическим растением, способным к повреждению можжевельника, снижая его устойчивость к неблагоприятным климатическим условиям. Оценено влияние гербицидов (ра-ундап и базагран) на снижение пораженности можжевельников [15]. В охраняемой зоне регионального парка дель Альто Мансанарес (Мадрид) изучена степень заражения можжевельника колючего A. оxycedri с целью отслеживания их распространения под воздействием лесных пожаров и человека [16].
Паразитарная направленность системы Juniperus-Arceuthobium подтверждена также с помощью рентгеновского микроанализа. Проведено сравнительное исследование минеральных элементов, содержащихся в тканях можжевельника и арце-утобиума, где показано, что эндофитные ткани паразита по содержанию S, P и, главным образом, K богаче, чем соседние ткани хозяев, и, следовательно, отношение K/Са выше в паразите [17]. Проведенный выше обзор литературных источников показывает, что A. oxicedri считается основной причиной деградации популяций видов Juniperus. Между тем прямых доказательств вредоносности A. oxicedri не приводится.
Цель работы: изучить структуры популяции и состояния кустов J. oblonga в зависимости от пораженности A. oxycedri , условий микроэкопотов и совместного произрастания P. kochiana на оползневом известняковом склоне хребта Чакулабек Внутреннегорного Дагестана.
Методика исследований. Работа проводились в 2013-2016 гг. на северо-восточном склоне хребта Чакулабек в окрестности с. Цудахар Левашинского района Внутреннегорного Дагестана на высотах от 1100 до 1400 м над уровнем моря. Общая площадь исследованного массива J. oblonga составила около 500 га. Склон представляет собой местами оголенный вследствие оползней массив известняковой плиты крутизной 30-40о, с фрагментарной фриганоидной растительностью и маломощным почвенным слоем в зависимости от условий экото-па. В последние десятилетия на этом участке наблюдается массовое расселение P. kochiana при одновременной деградации популяций J. oblonga . Почва горно-долинная лугово-степная намытая среднесуглинистая на древне-аллювиальных карбонатных суглинках. Исследованный склон имеет сложный микрорельеф. Для получения максимально объективной информации о зависимости состоянии растений J. oblonga и A. oxycedri от условий места произрастания выделены следующие экото-пы: оползневый, природниковый, оголенная известняковая плита, ложбина временного водостока, площадка террасы, шлейф из щебня, межложбинный гребень. Выделенные участки отличаются по мощности и влажности почвы, составу материнской породы, крутизне.
При закладке пробных площадей (ПП) и их описании руководствовались основными положениями, изложенными в работе «Методы изучения лесных систем» [19]. ПП по 400 м2 в количестве 20 шт. заложены на разных высотных уровнях склона: у основания, в середине и на вершине (в пределах 500 м) с учетом всего разнообразия экотопов и элементов рельефа.
Жизненность особей видов определена на основе подходов, изложенных в работах Уранова [20] и Злобина [21], по 5-ти бальной шкале на основе таких характеристик как хлоротичность, сморщенность побегов, наличие сухих побегов, паразитов и вреди- телей, мощность развития, длина годичного прироста, высота и диаметр кустов, наличие шишек. Возрастные группы и их характеристики даны по Т.А. Работнову [22] с учетом биологических особенностей изучаемых видов. Статистическая обработка результатов проведена с использованием дисперсионного, регрессионного и кластерного анализов с помощью программы Statistica v.5.5. По результатам дисперсионного анализа установлена степень зависимости (r2) и доля влияния (h2) факторов среды на показатели. Латинские названия растений даны по С.К. Черепанову [23].
Результаты и обсуждение . На основании полного учета кустов J. oblonga, произрастающих на площади около 500 га исследованного склона, проведена их группировка по возрасту, полу и состоянию, определена доля каждой из этих групп у основания, в середине и в верхней части склона (табл.1). Обнаружено, что с высотой по склону доля мужских кустов, кустов молодого возраста и здоровых кустов снижается, при одновременном значительном возрастании доли ослабленных кустов, у которых пол не определяется, и сухих кустов. Пораженность можжевельника A. oxicedri при этом вверх по склону явно усиливается при значительном возрастании участия P. kochiana в сообществе.
Таблица 1. Группировка кустов Juniperus oblonga и их пораженность полупаразитом Arceuthobium oxicedri на северо-восточном склоне хребта Чакулабек, в %
|
Группировки кустов |
Участки по склону |
||
|
нижний |
средний |
верхний |
|
|
всех возрастов |
37 |
48 |
15 |
|
молодые |
52 |
41 |
7 |
|
мужские |
22 |
14 |
7 |
|
женские |
25 |
23 |
21 |
|
пол не определяется |
53 |
63 |
72 |
|
здоровые |
27 |
19 |
6 |
|
полусухие |
45 |
48 |
43 |
|
сухие |
28 |
33 |
51 |
|
пораженные арцеутобиумом |
63 |
77 |
92 |
|
участие сосны |
1 |
16 |
83 |
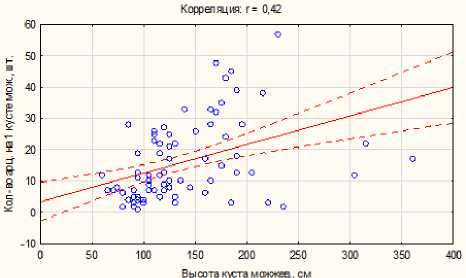
Рис. 1. Численность Arceuthobium oxicedri в зависимости от размеров кустов Juniperus oblonga
В целом на склоне преобладают особи J. oblonga генеративного состояния (52,9%) с существенным снижением доли молодых особей в верхней части склона. При этом на участках, где совместно с можжевельником произрастает сосна, молодые кусты первого имеют угнетенное состояние и встречаются лишь единично. Независимо от места произрастания на склоне доля кустов можжевельника с меньшими размерами, пораженных арцеутобиу-мом, больше, чем доля пораженных кустов с крупными размерами. Степень пораженности кустов можжевельника арцеутобиумом имеет линейную положительную зависимость от размеров кустов первого и не связана с условиями произрастания (рис. 1).
С высотой над уровнем моря по склону уменьшаются не только биометрические показатели кустов J. oblonga , но и A. oxicedri и эта тенденция подтверждена статистически (табл. 2). Кроме того, арцеутобиум слабее развит (6,3 см) на угнетенных (2,8 балла) кустах можжевельника, что объясняется полупаразитарным характером отношений между этими двумя видами. При этом различия между особями арцеутобиума с разных высот меньше (h2, 6,0%), чем между особями можжевельника с тех же высот (h2, 19,6%). Это и понятно, потому что состояние кустов можжевельника определяет развитие арцеутобиума, а не наоборот.
Таблица 2. Биоморфологические и виталитетные показатели Juniperus oblonga и Arceuthobium oxicedri и их статистическая оценка
|
Признаки |
Местопроизрастания по склону |
Общее |
F |
h2, % |
t-крит. (низ.-верх. склона) |
||||||
|
низ |
середина |
верх |
|||||||||
|
X |
CV, % |
X |
CV, % |
X |
CV, % |
X |
CV, % |
||||
|
A.oxycedri |
|||||||||||
|
длина, см |
7,5 |
35,8 |
6,7 |
21,9 |
6,3 |
20,1 |
6,8 |
28,5 |
2,9* |
6,0 |
2,22* |
|
ширина, см |
6,2 |
38,1 |
5,9 |
22,1 |
5,1 |
22,1 |
5,8 |
30,2 |
3,4* |
7,3 |
2,29* |
|
состояние, балл |
4,4 |
16,5 |
4,1 |
16,5 |
3,8 |
15,1 |
4,1 |
17,2 |
6,9** |
16,5 |
3,66*** |
|
число на 1 куст, шт. |
17,9 |
86,6 |
14,5 |
74,7 |
14,8 |
64,6 |
15,7 |
77,5 |
0,7 |
0,0 |
0,93 |
|
J. oblonga |
|||||||||||
|
высота куста, см |
165,7 |
42,5 |
124,5 |
32,8 |
114,8 |
30,9 |
135,0 |
41,0 |
8,3** |
19,6 |
3,54*** |
|
состояние, балл |
3,5 |
24,6 |
3,2 |
23,6 |
2,8 |
20,5 |
3,1 |
25,0 |
7,5** |
17,8 |
3,68*** |
Примечание: общее число изученных кустов = 90; здесь и далее уровни значимости – *-Р<0,05; **-Р<0,01; ***-Р<0,001
Видно, что степень угнетенности можжевеловых кустов не зависит напрямую от пораженности арцеутобиумом, а является следствием других причин, предварительно приведших к его ослаблению. Если бы состояние кустов можжевельника напрямую определялось степенью поражения арце-утобиумом, то угнетенность первых должна была быть выше при более сильном и массовом развитии арцеутобиума. Такое не наблюдается. Например, наиболее благополучные кусты можжевельника у основания склона (состояние 3,5 баллов и высота 165,7 см) сильнее поражены (17,9 на 1 куст). Кроме того, при увеличении плотности арцеутобиума на куст, их жизненное состояние внутри кустов можжевельника снижается (рис. 2).
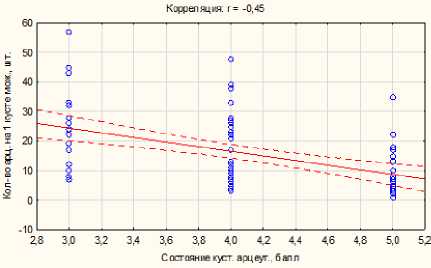
Рис. 2. Зависимость состояния Arceuthobium oxicedri от плотности их размещения на кустах Juniperus oblonga
Что касается коэффициентов вариации биометрических показателей, то они с высотой по склону снижаются, что может быть связано с ухудшением условий произрастания и общим снижением амплитуды показателей J. oblonga и A. oxicedri .
Как было отмечено выше, исследованный склон имеет не только высотное различие условий в пределах 500 метров, но и сложный микрорельеф с экотопами, различающимися по мощности почвы, влажности составу грунта. Зависимость (r2) показателей признаков можжевельника и арцеутобиума от условий экотопов составляет от 13,3 до 29,8%, за исключением признака «число арцеутобиума на 1 куст». Последнее, как было отмечено выше, зависит от размеров кустов J. oblonga, а не от условий произрастания последнего (табл. 3). Различие между участками, видимо, связано, прежде всего, с почвенными условиями, на что, на наш взгляд, указывает развитость кустов можжевельника. Кусты можжевельника, произрастающие в природниковых участках в три раза выше кустов, на щебнистых участках. Такая же, хотя и менее значительная разница наблюдается и в развитии арцеутобиума, что еще раз подтверждает зависимость размеров арцеуто-биума от размеров кустов можжевельника, но не наоборот. Кусты J. oblonga, произрастающие в условиях природникового луга, также имеют хорошие биометрические показатели, несмотря на более сильную пораженность арцеутобиумом. С ухудшением состояния кустов можжевельника (от 3,7 до 2,9 баллов) ухудшается и состояние паразита (от 4,5 до 3,9 баллов) и эта зависимость также статистически доказана. Т.е. не арцеутобиум является первопричиной угнетенного состояния кустов можжевельника.
Арцетобиум поселяется на ослабленных кустах можжевельника, и размеры кустов первого вида в последующем напрямую определяются первоначальным состоянием (до поражения) и размерами растения-хозяина.
Таблица 3. Биоморфологические и виталитетные показатели Arceuthobium oxicedri и Juniperus oblonga в зависимости от условий экотопов склона
|
Экотопы |
A. oxycedri |
J. oblonga |
||||
|
высота, см |
ширина, см |
состояние, балл |
число на 1 кусте, шт. |
высо та, см |
состояние, балл |
|
|
природниковый луг (n=11) |
9,5 |
7,7 |
4,5 |
24,2 |
235,9 |
3,7 |
|
площадка террасы (n=26) |
6,6 |
5,6 |
4,6 |
12,3 |
133,5 |
3,7 |
|
межложбинный гребень (n=11) |
7,6 |
6,5 |
3,9 |
12,9 |
123,2 |
2,9 |
|
оползневый участок (n=17) |
6,2 |
5,4 |
3,9 |
14,6 |
125,0 |
2,9 |
|
известняковая плита (n=10) |
6,4 |
5,5 |
3,7 |
21,9 |
122,0 |
2,7 |
|
ложбина водостока (n=8) |
5,9 |
5,0 |
3,3 |
16,0 |
97,5 |
2,3 |
|
шлейф из щебня (n=7) |
5,3 |
4,6 |
3,9 |
13,0 |
86,4 |
2,9 |
|
F-критерий |
7,8*** |
4,5*** |
8,5*** |
1,9 |
15,7*** |
8,4*** |
|
h2, % |
35,5*** |
22,3*** |
37,6*** |
7,1 |
54,4*** |
37,3*** |
|
г2, % |
19,2*** |
13,3*** |
28,0*** |
0,1 |
29,8* |
27,6* |
Примечание: n - число изученных кустов J. oblonga
Что касается корреляционной взаимосвязи признаков A. oxicedri и J. oblonga, то, прежде всего, отметим функциональную зависимость (г - 0,93) высоты и ширины кустов первого (табл.4). Это и понятно, так как арцеутобиум имеет почти шарообразную форму, у которого всякие биометрические изменения кроны происходят синхронно. На высоком уровне значимости подтверждена корреляционная зависимость и между признаками арцеуто-биума и можжевельника. Положительная зависимость биометрических показателей арцеутобиума с таковыми можжевельника мы объясняем поражением кустов последнего в раннем возрасте. В силу этого рост и развитие арцеутобиума происходит одновременно с ростом и развитием кроны можжевельника. Единовременное и массовое поражение еще молодых кустов можжевельника, как было отмечено выше, можно объяснить, общим снижением защитных механизмов, в т.ч. слабым развитием покровных тканей или наличием микротрещин на коре ослабленных кустов и т.д. И, наконец, увеличение числа A. oxycedri приводит как к дополнительному ослаблению на кустов можжевельника (гху -0,47; -0,45 соответственно), так и к общему снижению виталитета полупаразитарного комплекса A. oxycedri - J. oblonga.
Таблица 4. Корреляционные взаимосвязи признаков Arceuthobium oxicedri и Juniperus oblonga , произрастающих на разных экотопах
|
Характеристика |
A. oxycedri |
J. oblonga |
|||
|
длина , см |
ширина, см |
число на 1 кусте, шт. |
высота куста, см |
состояние куста, балл |
|
|
ширина арцеут., см |
0,93*** |
||||
|
число арц. на 1 кусте, шт. |
0,38*** |
0,33** |
|||
|
высота куста можжев., см |
0,65*** |
0,65*** |
0,42*** |
||
|
состояние куста можжев., балл |
-0,03 |
0,00 |
-0,47*** |
0,18 |
|
|
состояние куст. арцеут., балл |
0,04 |
0,07 |
-0,45*** |
0,17 |
0,97*** |
Примечание : n=90.
Для оценки зависимости показателей арце-утобиума и можжевельника от условий произрастания проведен кластерный анализ (рис. 3). Участки при этом на дендрограмме сгруппировались по близости почвенных условий, т.е. с более благоприятными и менее неблагоприятными условиями разместились на противоположных сторонах. Межложбинные гребни и оползневые обрывистые участки, составляющие один кластер, видимо, одинаково засушливы, хотя по составу грунта между участками разница значительная.
Выше было отмечено, что наш интерес к изучению популяции J. oblonga на оползневом известняковом склоне хребта Чакулабек Внутреннегорного Дагестана был вызван выявленным нами ранее [7] угнетенным состоянием кустов можжевельника на фоне хорошего состояния других древесных, кустарниковых и травянистых растений на этом склоне. Фактором, изменившим здесь структуру популяции можжевельника в неблагоприятном направлении, мог быть чрезмерный выпас скота. В литературе по этому вопросу мнения расходятся. Например, Б.Н. Шамшиев [24) указывает на устойчивость арчевников Киргизии к выпасу, а В.М. Джанаева [25] и К.Ф. Мухамедшин [26] считают, что выпас скота на популяции можжевельника оказывает негативное влияние. На наш взгляд, этот фактор на изученном склоне не является решающим, так как здесь встречаются отдельные особи разных возрастов с высокой жизненностью. Кроме того, в последние десятилетия на этой территории наблюдается значительное снижение пастбищной нагрузки, что, однако, не привело к улучшению состояния особей J. oblonga.
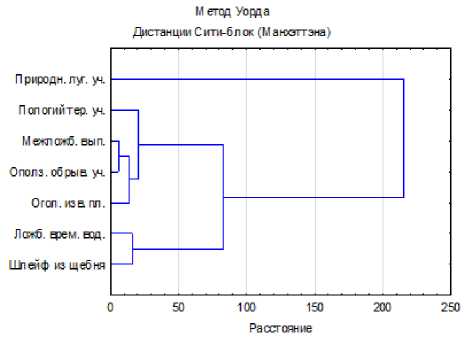
Рис. 3. Дендрограмма близости экотопов северовосточного склона хребта Чакулабек по совокупности признаков растений арцеутобиума и можжевельника
В аридных условиях Внутреннегорного Дагестана сообщества с доминированием J. oblonga распространены только на северных, западных и, частично, восточных склонах. На больших высотных уровнях Среднегорного и Высокогорного Дагестана можжевельник произрастает и на южных склонах. По нашим наблюдениям на северных и смежных с ним склонах на определенном этапе в можжевеловых редколесьях массово начинает распространяться Pinus kochiana, постепенно вытесняя J. oblonga . И нам представляется, что в такой ситуации P. kochiana, как будущий доминант, может оказывать определенное отрицательное влияние на растения J. oblonga .
Как известно, существует три варианта реагирования ценопопуляций растений на внешние воздействия: изменение жизненного состояния, численности и типа возрастного спектра [27]. В нашем случае в популяции можжевельника на хребте Чакулабек явно проявляются все три варианта последствий, но только в негативную сторону. Первоначально наблюдается снижение жизненности кустов можжевельника в связи с массовым распространением и совместным произрастанием сосны Коха. Затем на ослабленных под влиянием сосны кустах можжевельника поселяется полупаразит арцеуто-биум, увеличение плотности которого приводит к уменьшению скорости роста и снижению численности можжевельника. В конечном счете снижение жизненного состояния приводит к значительному смещению возрастного спектра, и ослаблению влияния можжевельника на ценоз [20, 28], происходит смена растительного сообщества – можжевеловых редколесий сосновыми лесами. Такова, на наш взгляд, динамика развития полупаразитарного комплекса – арцеутобиум-можжевельник и демута-ционного процесса на оползневом склоне хребта Чакулабек Внутреннегорного Дагестана при массовом распространении P. кochiana.
Список литературы Состояние популяции Juniperus communis subsp. oblonga bieb. в зависимости от распространения полупаразита Arceuthobium oxycedri (DC.) Bieb. и совместного произрастания c Pinus kochiana Klotzsch в условиях Внутреннегорного Дагестана
- Афанасьева, Н.Б. Введение в экологию растений: учебное пособие/Н.Б. Афанасьева, Н.А. Березина.-М.: Изд-во МГУ, 2011. 800 с.
- Онипченко, В.Г. Функциональная фитоценология: Снэкология. Учебное пособие. -М., 2014. 576 с.
- Press, M., Barker, M., Scholes, J. (eds) Physiological plant ecology. British Ecological Society Symposium. -Oxford: Blackwell Science, 1999. № 39. 480 p.
- Львов, П.Л. Фрагменты арчевого редколесья в предгорьях Дагестана//Научные доклады высшей школы. Биол. науки. 1963. №1. С. 120-124.
- Львов, П.Л. Сохраним Талгинские редколесья//Первая дагестанская республиканская конференция по охране природы. -Махачкала, 1968. С. 14-16.
- Асадулаев, З.М. Некоторые подходы к анализу структурных особенностей популяции Juniperus oblonga на территории памятника природа Дагестана «Сосновка»/З.М. Асадулаев, М.Д. Залибеков//Вестник ДНЦ РАН. 2011. № 41. С. 30-40.
- Асадулаев, З.М. Структурная и ресурсная оценка природных популяций Juniperus oblonga Bieb. в Дагестане/З.М. Асадулаев, Г.А. Садыкова. -Махачкала, 2011. 216 с.
- Ciesla, W.M. Hosts and Geographic Distribution of Arceuthobium oxycedri/W.M. Ciesla, B.W. Geils, R.P.Аdams//2002. РМРС-RN-11WWW: http://www.fs.fed.us/rm/pubs/rmrs_rn11/.
- Adams, R.P. Systematics of multi seeded eastern hemisphere Juniperus based on leaf essential oils and RAPD DNA fingerprinting//Biochem Syst Ecol. 1999. Vol. 27. 709-725.
- Садиков, Х.Х. О распространении Arceuthobium oxycedri (ДС). М.В. в бассейне реки Искандер/Х.Х. Садиков, М. Дарвозиев//Вестник Таджикского национального университета. 2 (66), 2011. Душанбе: СИНО. С. 37-40.
- Садиков, Х.Х. Арчевники бассейна реки Искандер. Автореферат дисс. …к.б.н. -Новосибирск, 2012. 16 с.
- Сухарева, А.О. Экологическое состояние и перспективы заповедования природного комплекса Чилтер (западный Крым)/А.О. Сухарева, О.И. Оскольская//Экосистемы, их оптимизация и охрана. 2009. Вып. 20. С. 212-223.
- Sarangzai, A.M. A. New spread of dwarf mistletoe (Аrceuthobium oxycedri) in Juniper forests/A.M. Sarangzai., N. Khan, M. Wahab, A. Kakar//Pakistan. Journal of botany. 2010/42(6). 3709-3714.
- Wahid, H.A. Dwarf mistletoe Arceuthobium oxycedri and damage caused by dwarf mistletoeto family Cupressaceae/H.A. Wahid, M.Y.K. Barosai, M. Din//Pure Appl. Bio. 2015. 4 (1). P.15-23.
- Kavosi, M.R. Effects of foliar application herbecides to control semi-parasitic plant Arceuthobium oxycedri/M.R. Kavosi, F. Faridi, G. Hajizaden//Bioscience. 2012. Vol. 4. No. 2. P. 76-80.
- Zavala, I. Global Positioning System as a tool for ecosystem studies at the landscape level: an application in the Spanish Mediterranean/I. Zavala, M.A. Zavala//Landscape and Urban Planning. 1993. Vol. 24, Is. 1-4. P. 95-104.
- Rey, L. Trophic Relations of the Dwarf Mistletoe Arceuthobium oxycedri with its Host Juniperus excelsa/L. Rey, A. Sadik., A. Fer, S. Renaudin//Journal of Plant Physiology. 1991. Vol. 138, Is. 4, August. P. 411-416.
- Садыкова, Г.А. Структурная и ресурсная оценка природных популяций можжевельника продолговатого (Juniperus oblonga Bieb.) в Дагестане//Диссер…. к.б.н. -Махачкала, 2010. 187 с.
- Методы изучения лесных сообществ. -СПб.: НИИ Химии СПбГУ, 2002. 240 с.
- Уранов, А.А. Жизненное состояние вида в растительном сообществе//Бюллетень МОИП. 1960. Т. 65. № 3. С. 77-91.
- Злобин, Ю.А. Ценопопуляционная диагностика экотопа//Экология. 1980б. № 2. С. 22-30.
- Работнов, Т.А. Вопросы изучения состава популяции для целей фитоценологии//Проблемы ботаники. 1950 б. Вып. 1. С. 465-483.
- Черепанов, С.К. Сосудистые растения России и сопредельных государств. -СПб., 1995. 992 с.
- Шамшиев, Б.Н. Влияние выпаса скота и режима заповедования на лесные культуры арчи в Киргизии//Лесной журнал. 1995. № 2-3. С. 192-195.
- Джанаева, В.М. Естественное возобновление арчи в урочище Киргиз-Ата//Труды КиргизЛОС. -Фрунзе, 1958. Вып 1. С. 43-56.
- Мухамедшин, К.Ф. Арчовые леса и редколесья Южной Киргизии//Труды КиргЛОС. -Фрунзе: Кыргызстан, 1967. Вып. 5. С. 247.
- Заугольнова, Л.Б. Погодичная динамика ценопопуляций некоторых видов песчаных степей Наурзумского заповедника//Ценопопуляции растений: развитие и взаимоотношения. -М.: Наука. 1977. С. 36-50.
- Уранов, А.А. Возрастной спектр фитоценопопуляции как функция времени и энергетических волновых процессов//Научные доклады высшей школы. Биол. науки. 1975. №2. С. 7-33.
