Современное состояние флоры хребта Малый Балхан (Туркменистан)
")
Автор: Павленко А.В., Лактионов А.П., Мурзаханов Р.
Журнал: Фиторазнообразие Восточной Европы @phytodiveuro
Статья в выпуске: 1 т.20, 2026 года.
Бесплатный доступ
Приводятся данные о современном состоянии флоры Малого Балхана – небольшого горного хребта, расположенного в Западном Туркменистане. Представлены данные о жизненных формах растений, проведен экологический, хорологический и эколого-фитоценотический анализы, сравнительная характеристика с сопредельными районами. Составлен аннотированный конспект флоры, включающий 401 вид растений из 56 семейств. Приведены сведения об узколокальных эндемичных видах.
Растительность, жизненные формы, эндемик, Ирано-Туран, мезофиты, Красная книга Туркменистана, Каракумы, Большой Балхан
Короткий адрес: https://sciup.org/148332997
IDR: 148332997 | УДК: 581.9 | DOI: 10.24412/2072-8816-2026-20-1-64-99
Current state of the flora of the Lesser Balkhan ridge (Turkmenistan)
The article presents data on the current state of flora of the Lesser Balkhan, a small mountain range located in Western Turkmenistan. Data on plant life forms are presented, ecological, chorological and ecological-phytocenotic analyses are conducted, and a comparative description with adjacent areas is provided. An annotated flora list has been compiled, including 401 plant species from 56 families. Information on narrow-local endemic species is provided.
Текст научной статьи Современное состояние флоры хребта Малый Балхан (Туркменистан)
Малый Балхан – небольшой горный хребет, занимающий относительно небольшую территорию, представленную весьма интересной уникальной флорой. Регион не является труднодосягаемым: вокруг расположен целый ряд населённых пунктов. С севера к хребту вплотную пролегает железная дорога, а с юга автотрасса Ашхабад – Туркменбаши (Красноводск) (рис. 1). Тем не менее, флора и растительность этих гор остается малоизученной.
Первым исследователем, посетившим Малый Балхан, был А.А. Антонов, который в 1889 г. в течение одного дня обследовал северные предгорья, назвав их «каменистой степью» (Antonov, 1890). В начале июня 1931 г. небольшой гербарный материал из южной подгорной равнины и предгорий Малого Балхана был собран Н. С. Нардиной. Основательно данный район был изучен в послевоенные годы в конце 1930-х – начале 1950-х гг. выдающимся исследователем Р.П. Тарасовым. В ходе работы были изучены особенности рельефа, геологии, составлены геоботанические описания растительности, картирование всего Малого Балхана, а также аннотированный флористический список (Tarasov, 1954), который намного позже был незначительно дополнен некоторыми видами (Kurbanov, 1988).
Материалы и методика
Обследования Малого Балхана впервые проведены в мае 2018 г. Исследования продолжались в 2019 (апрель), 2022 (апрель – июнь) и в 2023 (июль). На основной хребет подъем осуществлялся по мотоциклетному маршруту на северном склоне, а также пешим ходом с южной стороны. Подъём пешим ходом во многом облегчают многочисленные тропы скота, выпасаемого здесь круглый год.
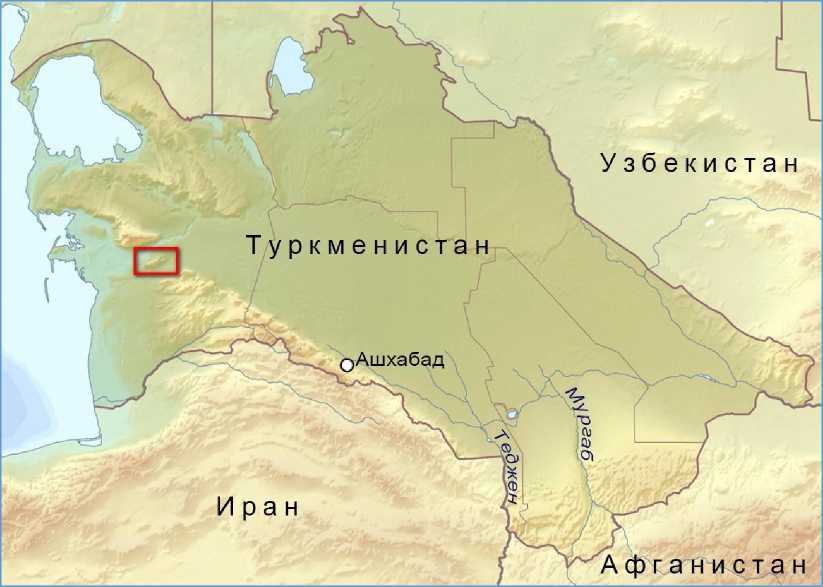
Рис. 1. Расположение Малого Балхана в Туркменистане
Fig. 1. Location of Lesser Balkhan in Turkmenistan
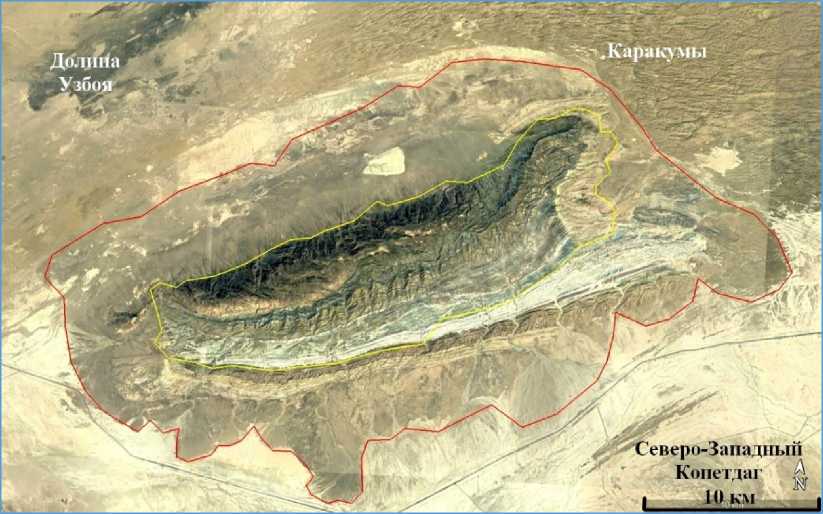
Рис. 2. Спутниковая карта Малого Балхана: желтой линией очерчен главный хребет, красной – предгорья (Google Earth Pro, 2025)
Fig. 2. Satellite map of Lesser Balkhan: the yellow line outlines the main ridge, the red line – the foothills (Google Earth Pro, 2025)
Исследования флоры осуществлялось маршрутным методом с параллельным составлением геоботанических описаний. Все местонахождения фиксировались GPS-приемником, встроенном в фотокамеру Canon Mark II. Проводился сбор гербарных образцов наиболее интересных видов. Собрано около 250 листов. Ежедневно закладывались пешие маршруты по 4–10 км в зависимости от сложности рельефа. Площадь популяций наиболее интересных видов оценивалась глазомерно. Ряд наблюдений фиксировали фотосъемкой.
Для определения видов мы руководствовались региональными «Флорами» и «Определителями» (Nikitin, Geldikhanov, 1988; Opredelitel rasteniy, 1968–1993).
Нами проведён анализ флористического состава флоры обоих хребтов и их предгорных частей. Составлен конспект флоры. При анализе и работе над конспектом использованы результаты собственных гербарных сборов и фотографический материал, обработка коллекций Национального гербарного фонда Туркменистана (ASH), просмотренны виртуальные образцы Цифрового гербария МГУ им. М. В. Ломоносова (MW) (URL: , некоторые оцифрованные образцы Ташкентского института ботаники (TASH), а также использованы литературные данные прошлых исследований (Nikitin, Gelgikhanov, 1988; Kurbanov, 1988).
Научные названия растений приводятся в соответствие с базами данных IPNI (URL: и POWO (URL: . Для определения площадей некоторых участков использована программа Google Earth Pro (2025).
Гербарный материал хранится в Национальном гербарном фонде Туркменистана (ASH).
Обследованная территория
Малый Балхан по сравнению с прочими горными регионами страны отличается небольшой площадью – около 500 км2, ограничивается широтами 39º11´ и 39º24´ и долготами 54º44´ и 55º9´ (рис. 2). Максимальная высота отмечается в западной части хребта (784 м, 39º17´6.17″ N, 54º50´52.38″ E). Крайняя северная точка расположена на окраине ж/д ст. Перевал (39º 24´ 22.80″ N, 55º0´22.43″ E). Южная точка упирается в автотрассу Ашхабад–Туркменбаши (Красноводск) (39º11´30.74″ N, 54º55´4.85″ E). Восточный край в 3,5 км южнее ж/д ст. Ахчакуйма (39º18´57.04″ N, 55º9´58.62″ E), а западная точка в 13 км восточнее г. Кумдаг (39º15´36.44″ N, 54º44´56.58″ E). Расстояние от северной до южной точки 25 км, а от восточной до западной – 37 км. Малый Балхан представляет собой целый комплекс холмов с центральным главным хребтом, верх которого также напоминает плато площадью около 60 км2. Южные склоны достаточно пологие, а северные крутые, скалистые, труднодоступные.
Согласно ботанико-географическому районированию Малый Балхан в составе Закаспийской провинции Переднеазиатской группы Азиатской пустынной области Кюрендагского округа, вместе с низкогорьями северо-западного Копетдага (Prozorovskiy, Maleyev, 1947). В более поздних источниках хребет включают в состав Южнотуранской провинции Ирано-Туранской подобласти Сахаро-Гобийской области (Botanical geography, 2003). Что касается флористического районирования, то весь Балханский район более близок ко флоре прочих горных фитохорий, в частности, ко флоре Копетдаг-Хорасанской провинции Переднеазиатской области подцарства Древнего Средиземноморья (Kamelin, 2018), чем ко флоре пустынь Южного Турана.
Климат
Климат Малого Балхана резко континентальный аридный, исключительно сухой – испаряемость во много раз превышает количество осадков. Ближайшая к Балханам метеостанция расположена на равнине в городе Берекет (бывш. Казанджик) (высота 35 м над ур. м., 39.2500º N, 55.5167º E) (Arkhiv pogody, 2025). Средняя годовая температура воздуха для этого пункта 17.1ºС (1973–2024 гг). Анализ данных показывает стабильный рост среднегодовых температур каждое десятилетие (рис. 3). За 50 лет в регионе потеплело на 1.6 ºС. Абсолютный минимум зафиксирован 06 февраля 2012 г. (–23.7 ºС). Большое значение имеет ветровой режим. В описываемом регионе он сравнительно высокий. Ветра отмечаются не менее 250 дней в году, нередки пыльные бури, а порывы достигают 30–40 м/с. Среднегодовая скорость ветра за последние 10 лет составляет 3.3 м/с. Осадков за год в среднем выпадает 147 мм (2020–2024 гг).
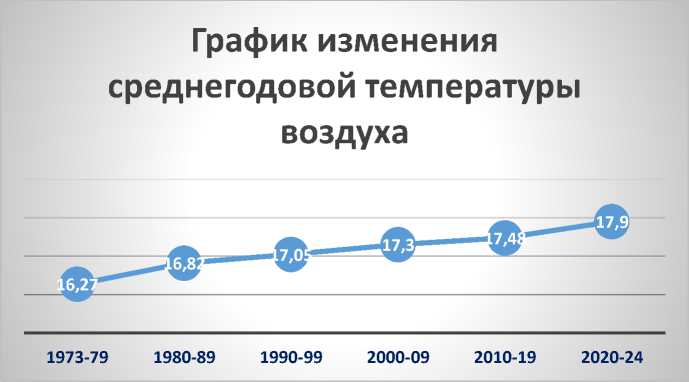
Рис. 3. Изменение среднегодовых температур (ºС) (1973–2024 гг)
Fig. 3. Change in average annual temperatures (ºС) (1973–2024)
Геология
Для Малого Балхана характерны обнажения плотных пород (скалы), слагаемые из плотных известняков и известняково-песчаниковых пород, глинистых и мергелистых известняков, а также плотных песчаников (Tarasov, 1954).
В северо-восточной части главного хребта обнажаются пласты рыхлых мергелей. В центральной части данной горной системы отмечается довольно широкая продольная (с востока на запад) депрессия составленная, по большей части, рыхлыми глинами. В предгорьях повсеместны холмы, сложенные палеогеновыми пестроцветными глинами. Крупные участки заняты холмогорьями с выходом гипсоносных пород.
Результаты и обсуждения
Растительность
Растительность хребта Р. П. Тарасов рассматривал по приуроченности к различным геологическим обнажениям: обнажения плотных пород (скалы) с растительностью по трещинам и щелям скал; обнажения относительно неплотных пород с явно выраженной ассоциированностью растительности; обнажения неплотных глинистых пород, подвергающихся сильной эрозии и представленная единичными, далеко отстоящими друг от друга растениями и прочие биотопы (Tarasov, 1954).
Южная предгорная часть представляет собой сильно всхолмленную равнину. Холмогорья, сложенные пестроцветными палеогеновыми глинами с весьма скудной растительностью – бедленды (рис. 4), представители которой чаще всего гипсофиты и галофиты, занимают всю южную часть и подходят вплотную к главному хребту, в нижней части склонов которого также отмечаются выходы палеогеновых глин (рис. 5). Склон достаточно пологий и эдификатором сообществ является Zygophyllum atriplicoides с небольшим набором травянистых монокарпиков. Вблизи немногих родников (ур. Чалсу и Торонгылы) отмечены небольшие рощицы Populus euphratica Olivier и единичные угнетённые особи Salix excelsa S. G. Gmel. Северные склоны главного хребта Малого Балхана довольно крутые и труднодоступные. Скалистые склоны изрезаны глубокими ущельями, поднимающимися до платообразной верхушки хребта (рис. 8).
Подгорная равнина, селевые русла покрыты крупными валунами. Подгорная и предгорная часть представлена формацией Haloxylon ammodendron с субэдификаторами Caroxylon orientale (S.G. Gmel.) Tzvelev и Xylosalsola arbuscula (Pall.) Tzvelev. На состав трав сильное влияние оказывает тесное соседство Малого Балхана с песчаной пустыней (рис. 7). Среди травянистой растительности подгорной и предгорий северной части хребта значительную долю составляют псаммофиты. Крутые скалистые склоны практически лишены хазмофитов. Пояс шибляка на Малом Балхане не выражен. Самая верхняя платообразная часть главного хребта характеризуется полустепной растительностью, преимущественно злаково-разнотравнополынной (рис. 6). Арчи практически нет (всего 2 экземпляра на весь Малый Балхан).
Растительность пестроцветных обнажений плато крайне скудна и представлена единичными особями растений.

Рис. 4. Бедленды южных предгорий Малого Балхана
-
Fig. 4. Badlands of the southern foothills of the Lesser Balkhan

Рис. 5. Пестроцветные выходы палеогеновых глин
-
Fig. 5. Variegated outcrops of Paleogene clays

Рис. 6. Разнотравно-полынные сообщества на гребне главного хребта
Fig. 6. Forb-wormwood communities on the crest of the main ridge

Рис. 7. Полоса контакта Малого Балхана и Каракумов
Fig. 7. Contact strip of the Lesser Balkhan and the Karakum Desert

Рис. 8. Скалистый склон северной экспозиции
Fig. 8. Rocky slope with northern exposure
Флора
Флора Малого Балхана представлена 3 отделами: Equisetophyta (1 род, 1 вид), Pinophyta (2 рода, 3 вида), Magnoliophyta (397 видов). Соотношение однодольных и двудольных видов выражается пропорциональной зависимостью 1:6.5. Распределение таксонов исследуемой флоры по крупным систематическим группам (табл.1) характерен для флор аридных районов умеренного климата.
Таблица 1. Распределение видов по классам сосудистых растений
Table 1. Distribution of species by classes of vascular plants
|
Таксоны |
Малый Балхан |
|
|
Число таксонов |
Доля от общего числа, % |
|
|
Сосудистые споровые: Equisetopsida |
1 |
0.25 |
|
1 |
0.25 |
|
|
Голосеменные: Gnetopsida Pinopsida |
3 |
0.75 |
|
2 |
0.5 |
|
|
1 |
0.25 |
|
|
Покрытосеменные: Liliopsida Magnoliopsida |
397 |
99.0 |
|
53 |
13.2 |
|
|
344 |
85.8 |
|
|
Всего: |
401 |
- |
Доля споровых сосудистых растений в МБ очень мала (0,3%).
Ранее для флоры данного региона отмечалось 313 видов высших растений (Tarasov, 1954). По результатам исследований и обработки более поздних литературных материалов нами приводится для МБ 401 вид, относящихся к 56 семействам (табл. 2).
Таблица 2 . Распределение видов и родов по семействам флоры Балханов (APG-IV)
Table 2 . Distribution of species and genera by families of the flora of Balkhan (APG-IV)
|
№ |
Семейство |
Малый Балхан |
||
|
родов |
видов |
% видов |
||
|
1. |
Equisetaceae |
1 |
1 |
0,25 |
|
2. |
Cupressaceae |
1 |
1 |
0,25 |
|
3. |
Ephedraceae |
1 |
2 |
0,5 |
|
4. |
Melanthiaceae |
1 |
1 |
0,25 |
|
5. |
Liliaceae |
3 |
6 |
1,5 |
|
6. |
Amaryllidaceae |
1 |
5 |
1,2 |
|
7. |
Asparagaceae |
1 |
3 |
0,7 |
|
8. |
Asphodelaceae |
1 |
1 |
0,25 |
|
9. |
Iridaceae |
1 |
3 |
0,7 |
|
10. |
Ixioliriaceae |
1 |
1 |
0,25 |
|
11. |
Cyperaceae |
2 |
4 |
1,0 |
|
12. |
Juncaceae |
1 |
3 |
0,7 |
|
13. |
Poaceae |
18 |
26 |
6,5 |
|
14. |
Berberidaceae |
2 |
2 |
0,5 |
|
15. |
Papaveraceae |
5 |
11 |
2,7 |
|
16. |
Ranunculaceae |
4 |
4 |
1,0 |
|
17. |
Geraniaceae |
2 |
5 |
1,2 |
|
18. |
Biebersteiniaceae |
1 |
1 |
0,25 |
|
19. |
Nitrariaceae |
4 |
4 |
1,0 |
|
20. |
Rutaceae |
1 |
2 |
0,5 |
|
21. |
Thymelaeaceae |
1 |
2 |
0,5 |
|
22. |
Brassicaceae |
28 |
38 |
9,5 |
|
23. |
Capparaceae |
1 |
1 |
0,25 |
|
24. |
Cleomaceae |
1 |
2 |
0,5 |
|
25. |
Fabaceae |
8 |
28 |
7,0 |
|
26. |
Elaeagnaceae |
1 |
1 |
0,25 |
|
27. |
Rhamnaceae |
1 |
1 |
0,25 |
|
28. |
Rosaceae |
3 |
3 |
0,7 |
|
29. |
Urticaceae |
1 |
1 |
0,25 |
|
30. |
Euphorbiaceae |
2 |
13 |
3,2 |
|
31. |
Hypericaceae |
1 |
1 |
0,25 |
|
32. |
Linaceae |
1 |
1 |
0,25 |
|
33. |
Phyllanthaceae |
1 |
1 |
0,25 |
|
34. |
Salicaceae |
1 |
1 |
0,25 |
|
35. |
Zygophyllaceae |
2 |
7 |
1,7 |
|
36. |
Amaranthaceae |
27 |
44 |
11,0 |
|
37. |
Caryophyllaceae |
7 |
11 |
2,7 |
|
38. |
Frankeniaceae |
1 |
1 |
0,25 |
|
39. |
Plumbaginaceae |
2 |
3 |
0,8 |
|
40. |
Polygonaceae |
5 |
11 |
2,7 |
|
41. |
Tamaricaceae |
2 |
6 |
1,5 |
|
42. |
Primulaceae |
1 |
1 |
0,25 |
|
43. |
Apocynaceae |
3 |
3 |
0,7 |
|
44. |
Gentianaceae |
1 |
1 |
0,25 |
|
45. |
Rubiaceae |
5 |
9 |
2,2 |
|
46. |
Boraginaceae |
14 |
26 |
6,5 |
|
47. |
Convolvulaceae |
2 |
4 |
1,0 |
|
48. |
Solanaceae |
1 |
2 |
0,5 |
|
49. |
Lamiaceae |
11 |
13 |
3,1 |
|
50. |
Mazaceae |
1 |
1 |
0,25 |
|
№ |
Семейство |
Малый Балхан |
||
|
родов |
видов |
% видов |
||
|
51. |
Orobanchaceae |
2 |
4 |
1,0 |
|
52. |
Plantaginaceae |
3 |
3 |
0,7 |
|
53. |
Scrophulariaceae |
1 |
1 |
0,25 |
|
54. |
Asteraceae |
28 |
49 |
12,2 |
|
55. |
Caprifoliaceae |
2 |
4 |
1,0 |
|
56. |
Apiaceae |
12 |
17 |
4,2 |
|
Всего |
236 |
401 |
100 |
|
Весьма интересно сопоставление флор двух соседствующих регионов. Для Большого и Малого Балханов нами составлены «логические ряды» ведущих 10-ти семейств в виде прогрессии, убывающей по числу видов в семействах (рис. 9).
|
Большой Балхан |
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
|
Малый Балхан |
2 1 |
4 1 |
3 1 |
5 1 |
1 |
8 1 |
7 1 |
1 11 1 |
1 9 1 |
6 1 |
|
Каракумы |
4 1 |
2 1 |
3 1 |
5 1 |
1 |
8 1 |
12 1 |
7 1 |
6 1 |
13 1 |
Рис. 9. Логические ряды систематических структур Балханов и Каракумов.
Обозначения: 1 – Poaceae; 2 – Asteraceae; 3 – Brassicaceae; 4 – Amaranthaceae; 5 – Fabaceae; 6 – Caryophyllaceae; 7 – Apiaceae; 8 – Boraginaceae; 9 – Lamiaceae; 10 – Caprifoliaceae; 11 – Euphorbiaceae; 12 – Polygonaceae; 13 – Tamaricaceae
Fig. 9. Logical series of systematic structures of the Balkhan and Karakum deserts.
Designations: 1 – Poaceae; 2 – Asteraceae; 3 – Brassicaceae; 4 – Amaranthaceae; 5 – Fabaceae; 6 – Caryophyllaceae; 7 – Apiaceae; 8 – Boraginaceae; 9 – Lamiaceae; 10 – Caprifoliaceae; 11 – Euphorbiaceae; 12 – Polygonaceae; 13 – Tamaricaceae
Из рисунка видно, что логический ряд первых 3-х семейств флоры Большого Балхана составили Poaceae-Asteraceae-Brassicaceae, что соответствует флорам других горных регионов Туркменистана, где есть средний горный пояс растительности (Копетдаг и Кугитанг), но в последних в числе доминирующей тройки участвует Fabaceae. Во флоре Малого Балхана ведущую тройку составил ряд Asteraceae-Amaranthaceae-Poaceae, причем выдвижение на 2-ю позицию Amaranthaceae свидетельствует о тесном контакте с пустынями Турана. Это подтверждается сходством тройки ведущих семейств Каракумов и Малого Балхана. Во флоре последнего ксерофиты Asteraceae и Amaranthaceae доминируют во флоре, что является следствием более суровых климатических условий, отсутствия водных источников и относительно низким уровнем высот.
Наиболее многочисленными родами во флоре исследуемого региона являются Astragalus (20 видов), Euphorbia (12), Cousinia (8), Zygophyllum , Calligonum и Ferula (по 6), Allium (5), Artemisia, Erodium и Strigosella (по 4), что указывает на древнесредиземноморское происхождение описываемой флоры.
Флоры Малого и Большого Балханов соседствуют в географическом плане, но достаточно отличимы по составу, на что указывают низкие показатели сходства флор. Согласно коэффициента Жаккара ( K j ) и Сёренсена-Чекановского ( K sc ), степень сходства ББ и МБ составляет 0.39 и 0.56 (для сравнения: соответствующие коэффициенты для Юго-Западного и Центрального Копетдага соответственно 0.64 и 0.78). Общих для обоих Балханов – 280 видов (рис. 10).
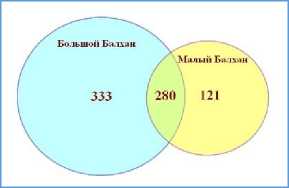
Рис. 10. Соотношения числа видов на Балханах
Fig. 10. Species abundance ratios in the Balkhans
Для более детальной характеристики флоры исследуемой территории мы провели анализ жизненных форм по самым популярным системам (Serebryakov, 1964; Varming, 1902; Raunkiaer, 1934).
Согласно системе И. Г. Серебрякова на Малом Балхане отмечается 27 жизненных форм растений (табл. 3, рис. 11).
Деревья представлены 2-мя группами. Первая группа – вечнозелёные, куда относится единственный вид – Juniperus turcomanica , в настоящее время представленный лишь двумя угнетенными экземплярами . К группе листопадных относятся обитатель песчаных наносов в предгорьях Ammodendron conollyi Bunge ex Boiss., Populus euphratica Olivier, образующий небольшую популяцию (около 20 экземпляров) в ущелье Торонгылы на южном склоне хребта и Elaeagnus orientalis L., произрастающий в числе 4-х особей у родника Чалсу.
Несуккулентные кустарники представлены группой из 14 видов, куда относятся Lycium depressum Stocks, L. ruthenicum Murray, Nitraria schoberi L., Zygophyllum atriplicoides , Haloxylon ammodendron и др. Из суккулентных кустарников выявлен единственный Halostachys caspica (M. Bieb.) C.A. Mey. К группе вечнозелёных относится род Ephedra с 2-мя видами.
Несуккулентные кустарнички представлены 10-ю видами: краснокнижным Malacocarpus crithmifolius (Retz.) C.A. Mey., оригинальным красивоцветущим Caragana grandiflora , эндемичным Cerasus turcomanica Pojark. и др. Подушковидный – весьма колючий кустарничек – Astragalus verus Olivier. Флора региона представлена одним несуккулентным полукустарником – Astragalus ammodendron Bunge. Таких же полукустарничков по Малому Балхану 28 видов (3 вида рода Reaumuria , Acanthophyllum elatius Bunge, редкий довольно обычный Andrachne telephioides L. и много др.). Всего 9 видов полукустарничков-суккулентов: Anabasis eriopoda (Schrenk) Benth. ex Volkens, Kalidium caspicum (L.) Ung.-Sternb., Kaviria aucheri (Moq.) Akhani, Salsola arbusculiformis Drobow, Xylosalsola arbuscula (Pall.) Tzvelev и др.). Все суккуленты относятся к Amaranthaceae.
Из травянистых многолетников наиболее представительна группа стержнекорневых поликарпиков (57 видов), куда относятся род Hypericum , Hedysarum micropterum Bunge ex Boiss., 10 видов рода Astragalus и др.).
К кистекорневым относится сорняк Plantago lanceolata L.
Всего 4 вида короткокорневищных трав: хазмофит Parietaria judaica L ., Vincetoxicum pumilum Decaisne, Achillea arabica Kotschy и A. wilhelmsii K. Koch.
Длиннокорневищные поликарпики представлены 31 видом: Equisetum ramosissimum Desf., 3 вида рода Asparagus , 3 вида Juncus и др.
К дерновинным многолетникам относятся Alopecurus arundinaceus Poir., Cenchrus orientalis (Rich.) Morrone, Poa bulbosa L., Stipa caucasica Schmalh., Stipagrostis karelinii (Trin. et Rupr.) Tzvelev и др. В этой группе 9 видов.
Клубнеобразующих 9 видов: Biebersteinia multifida DC., узколокальный краснокнижный Euphorbia oidorhiza Pojark. и Bongardia chrysogonum (L.) Spach. Луковичные травы представлены 11 видами ( Fritillaria karelinii (Fisch. ex D. Don) Baker, рода Gagea , Tulipa , Allium и др.), а к клубнелуковичным относятся Colchicum robustum (Bunge) Stef. и Ixiolirion tataricum (Pall.) Roem., Schult. et Schult. f. К клубнекорневищным относятся Eremurus kopetdaghensis Popov ex B. Fedtsch. и 3 вида рода Iris. К клубнелуковичнокорневищным относится только Bolboschoenus maritimus (L.) Palla.
Корнеотпрысковых 5 видов: сорные Alhagi pseudalhagi subsp. persarum , Convolvulus arvensis L., Rhaponticum repens (L.) Hidalgo, а также Karelinia caspia (Pall.) Less. и Mentha longifolia (L.) Huds.
Многолетников-паразитов 4 вида – все виды родов Cistanche и Orobanche .
К монокарпическим травам мы относим группы однолетних, двулетних и многолетних растений, плодоношение которых за весь жизненный цикл происходит всего раз. Сюда относится самая крупная группа растений – несуккулентные однолетние монокарпики (189 видов), куда мы отнесли как коротковегетирующие травы (эфемеры), так и однолетники длительной вегетации (летне-осенние). К первой группе относится целый ряд видов Poaceae, Brassicaceae, Boraginaceae, Lamiaceae, а ко второй группе, в основном, ряд видов-ксерофитов из Amaranthaceae и Asteraceae. Двулетние монокарпики включают 7 видов: Lappula barbata (M. Bieb.) Gurke, 4 вида рода Cousinia , Apium graveolens L. и Artemisia scoparia Waldst. et Kit. Суккулентами у монокарпиков являются Bienertia cycloptera Bunge и Tetradiclis tenella (Ehrenb.) Litv .
Единственным однолетним паразитом является Cuscuta brevistyla A. Braun ex A. Rich.
Многолетними монокарпиками являются 4 представителя рода Ferula .
Таблица 3. Спектр жизненных форм флоры согласно системе И. Г. Серебрякова
Table 3 . Spectrum of life forms of flora according to the system of I. G. Serebryakov
|
Жизненные формы |
Малый Балхан |
|
|
2 « п о 2 ^ s s « |
о ° й д Цо ® |
|
|
I . Деревья |
4 |
1.0 |
|
1. Вечнозелёные |
1 |
0.25 |
|
2. Листопадные |
3 |
0.7 |
|
II. Кустарники |
17 |
4.2 |
|
1. Несуккулентные |
14 |
3.5 |
|
2. Суккулентные |
1 |
0.25 |
|
3. Безлистные вечнозелёные кустарники |
2 |
0.5 |
|
III. Кустарнички |
11 |
2.7 |
|
1. Несуккулентные |
10 |
2.5 |
|
2. Подушковидные |
1 |
0.25 |
|
IV. Полукустарники и полукустарнички |
41 |
10.2 |
|
1. Несуккулентные |
28 |
7.0 |
|
2. Суккулентные |
9 |
2.2 |
|
3. Подушковидные несуккулентные |
3 |
0.7 |
|
4. Полукустарник несуккулентный |
1 |
0.25 |
|
V. Травянистые поликарпики |
125 |
31.2 |
|
1. Несуккулентные и в их числе: |
121 |
30.2 |
|
стержнекорневые, включая подушковидные |
57 |
14.2 |
|
кистекорневые |
1 |
0.25 |
|
короткокорневищные |
4 |
1.0 |
|
длиннокорневищные |
18 |
4.5 |
|
дерновинообразующие |
9 |
2.2 |
|
клубнеобразующие и клубнекорневые |
9 |
2.2 |
|
луковичные и клубнелуковичные |
13 |
3.2 |
|
клубнекорневищные |
4 |
1.0 |
|
клубнелуковично-корневищные |
1 |
0.25 |
|
корнеотпрысковые |
5 |
1.2 |
|
2. Паразитические травянистые поликарпики |
4 |
1.0 |
|
VI. Травянистые монокарпики |
203 |
50.6 |
|
1. Несуккулентные |
196 |
48.9 |
|
однолетние |
189 |
47.1 |
|
двулетние |
7 |
1.7 |
|
2. Суккулентные однолетние |
2 |
0.5 |
|
3. Паразитические однолетние монокарпики |
1 |
0.25 |
|
4. Многолетние клубнеобразующие |
4 |
1.0 |
|
Всего |
401 |
100 |
Таким образом, более половины флоры составляют однолетние растения, как способ переживания самого неблагоприятного периода (лето-осень-зима) в виде семян. В целом, процентные показатели различных групп растений как для Малого, так и для Большого Балхана схожи. Лишь доля полудревесной растительности Малого Балхана почти в 2 раза превышает этот показатель на Большом, что свидетельствует о более суровых условиях, практически близких к условиям туранских пустынь умеренного пояса (рис. 11, 12).
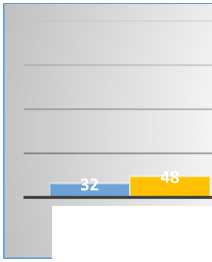
ДРЕВЕСНЫЕ
■ Малый Балхан ■ Большой Балхан
ПОЛУДРЕВЕСНЫЕ

ПОЛИКАРПИЧЕСКИЕ
ТРАВЫ
МОНОКАРПИЧЕСКИЕ
ТРАВЫ
Рис. 11. Спектр жизненных форм растений Малого и Большого Балханов
Fig. 11 . Spectrum of life forms of plants of the Lesser and Big Balkhans
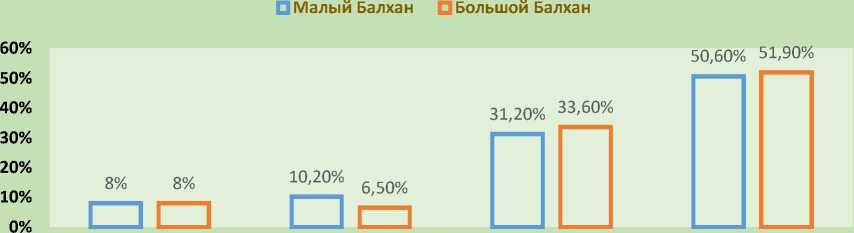
Древесные Полудревесные Поликарпические травы Монокарпические травы
Рис. 12. Соотношения жизненных форм Малого и Большого Балханов
-
Fig. 12. Relationships between life forms in the Small and Large Balkhans
Более упрощённая система жизненных форм К. Раункиера основана на расположении почек возобновления относительно почвогрунтов (рис. 13). Система наглядно характеризует флористический состав описываемой флоры.
К фанерофитам мы относим все деревья и кустарники; к хамефитам – полудревесные (полукустарники и полукустарнички); гемикриптофиты – многолетние травы без видоизменённой корневой системы; криптофиты – виды, у которых почки возобновления находятся на видоизменениях корней и стеблей, а также водные растения; терофиты – все однолетние травы.
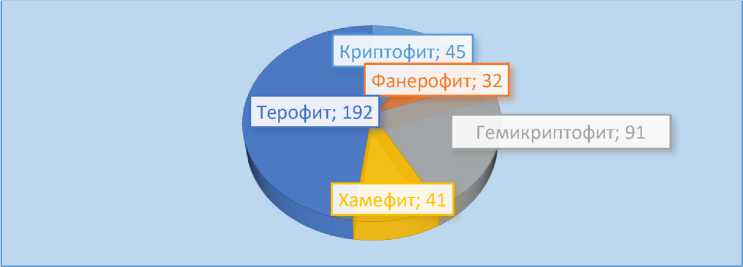
Рис. 13. Спектр жизненных форм растений Малого Балхана по системе Раункиера (Raunkiaer, 1934)
-
Fig. 13. Spectrum of life forms of plants of the Lesser Balkhan according to the Raunkiaer system (Raunkiaer, 1934)
По отношению к влагообеспечению мы распределили все виды по классификации Е. Варминга (рис. 14).
Из рис. 13 следует, что на исследуемой территории большинство видов относятся к мезофитам (57.9%), т.е. растениям, предпочитающим достаточное увлажнение. Обеспеченность влагой на Малом Балхане значительно меньше, чем, к примеру, на Большом. Вследствие этого доля мезофитов на Малом Балхане меньше (57.9% к 67.7%). Дополнительным фактом, подтверждающим вышесказанное, является более высокая доля ксеромезофитов и ксерофитов на Малом – 38.4%, тогда как на Большом Балхане она составляет 23.4%.
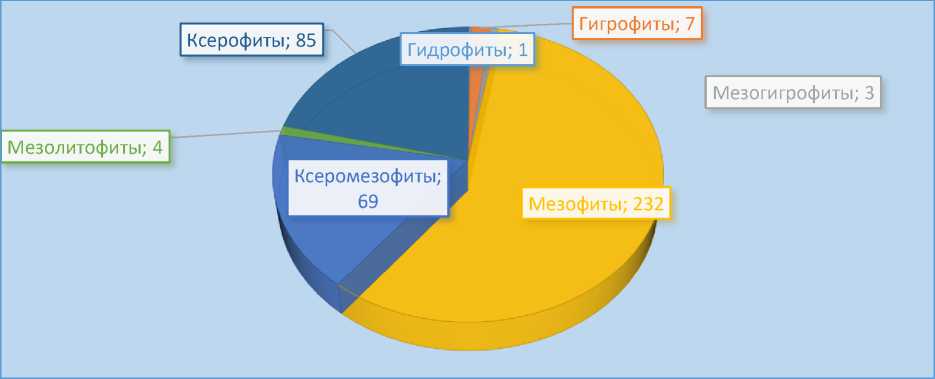
Рис. 14. Спектр жизненных форм по отношению к влагобеспечению на Малом Балхане по системе Варминга (1902)
-
Fig. 14. Spectrum of life forms in relation to moisture supply on the Lesser Balkhan according to the Warming system (1902)
Единственным гидрофитом является Equisetum ramosissimum Desf. К мезолитофитам относятся 4 вида: Juniperus turcomanica , Parietaria judaica L ., Vincetoxicum pumilum Decaisne и эндемичный Asperula balchanica Bobrov. К мезоксерофитам относятся Rubia florida Boiss., Argusia sogdiana (Bunge) Czerep., Diarthron vesiculosum (Fisch. et C.A. Mey. ex Kar. et Kir.) C.A. Mey., Tetracme recurvata Bunge и многие др. Вторая по величине группа – ксерофиты – включает представителей из 14-ти семейств и самые многочисленные по ксерофитам – это Asteraceae и Amaranthaceae, а семейства Ephedraceae и Asparagaceae представлены исключительно ксерофитами. Мезофиты – самая многочисленная группа и наибольшее число таких видов относятся к Poaceae, Brassicaceae и Fabaceae.
По отношению и устойчивости к засолению почв все виды нами разделены на 4 группы: гликофиты, галогликофиты, галофиты, а также особая группа растений-гипсофитов, местообитания которых связаны с меловыми холмами и пестроцветными обнажениями гипсоносных глин (рис. 15).
Гликофиты – растения незасоленных местообитаний, представлены 289 видами и составляют около 70,3% от всей флоры. Галогликофиты распространены как на незасоленных, так и на умеренно минерализованных почвогрунтах и на исследуемой территории отмечается 48 видов. К ним относятся Populus euphratica , Caroxylon dendroides (Pall.) Tzvelev, Asparagus brachyphyllus Turcz., Limonium reniforme (Girard) Lincz. и многие др.
Галофиты в числе 19 видов: Frankenia hirsuta L., Aeluropus lagopoides (L.) Thwaites, Nitraria schoberi L., Reaumuria fruticosa Bunge и др.
Из 45 гипсофитов примерами могут служить Plocama bruguierei (A. Rich. ex DC.) M. Backlund et Thulin, Biebersteinia multifida DC., Crambe edentula Fisch. et C.A. Mey. ex Korsh., Cleome turkmena Bobr. и многие др.
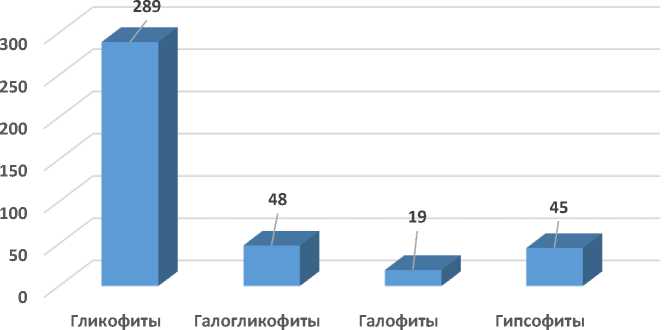
Рис. 15. Спектр видов по отношению к засолению почв
-
Fig. 15. Species spectrum in relation to soil salinity
Относительно высокая доля галофитов и гипсофитов на Малом Балхане указывает на более широкую распространенность участков, связанных с избыточным засолением и обнажением гипсоносных пород.
Для выявления центров многообразия и генезиса аборигенной флоры Малого Балхана нами проведено разделение всех видов на 4 класса (генетические элементы) с общим происхождением. Анализ данного дробления позволяет оценить положение в пределах Переднеазиатской области Древнесредиземноморья. В основе лежит региональный принцип выделения географического элемента для гор Средней Азии (Kamelin, 1971, 1973).
При проведении географического (хорологического) анализа аборигенной флоры Малого Балхана нами выявлен 42 элемента из 9 групп, относящихся к 4 классам (табл. 4).
В дополнительной 10-ой группе два представителя адвентивной (заносной) флоры ( Amaranthus albus L. и A. blitoides S. Watson), присутствующих в фитоценозах исследуемой территории.
Плюрирегиональная группа одноимённого класса включает широкораспространённые по всему Земному шару виды: Bolboschoenus maritimus, Juncus articulatus, J. bufonius, Phragmites australis, Centaurea solstitialis и др. Обычно это сорные рудеральные растения.
К голарктическим элементам относятся Echinops ritro L., Equisetum ramosissimum, Chenopodium album L., Polygonum aviculare L. и Galium spurium L. К палеарктическим видам мы отнесли Asperugo procumbens L., Lactuca tatarica (L.) C.A. Mey., Descurainia sophia (L.) Webb ex Prantl и пр. Голарктические и палеарктические элементы чаще всего относятся к сорным растениям. В целом, виды первых 3-х групп составили 5.2% от всей флоры.
Исходя из таблицы можно утверждать, что виды древнесредиземноморского генезиса во флоре исследуемого района занимают достаточно высокое положение (141 вид; 35.2%). К древнесредиземноморскому классу ареалов мы относим не только древнесредиземноморские виды, но и восточносредиземноморские, среди которых имеются виды, проникающие в северное Причерноморье, Восточную Европу и Южную Сибирь.
В составе флоры Малого Балхана явно доминирует Ирано-Туранский класс (237 видов; 59.1%). Самым крупным географическим элементом является туранская группа, у которой происхождение видов связано с холодными пустынями Туранской низменности. Вместе с ирано-туранской группой эти виды являются ядром ксерофильной флоры изученного района. В составе класса копетдаг-хорасанская группа, охватывающая весь Копетдаг и северную часть Иранского нагорья, представлена как копетдаг-хорасанским, так копетдагским, западнокопетдагским и балханским элементами, объединяющими собственно эндемичные виды. Наиболее крупной эндемичной группой являются копетдаг-хорасанские эндемики, которые на территории Туркменистана встречаются как на Балханах, так и в Копетдаге, а за пределами страны – на севере Ирана. Наиболее редкими из них являются Anthochlamys turcomanica Iljin, Jurinea antonowii C. Winkl. и Heliotropium litvinovii Popov, причем первые два вида на территорию Большого Балхана не заходят. В этой же группе занесенный в последнее издание Красной книги Туркменистана Juniperus turcomanica. В группе копетдагских эндемиков обычные виды: Astragalus curvipes Trautv., Elwendia longipes (Freyn) Pimenov et Kljuykov и Paracaryum turcomanicum Bornm. et Sint. Западнокопетдагскими являются 6 видов, из которых два занесены в Красную книгу: Eremurus kopetdaghensis и Cleome turkmena. Помимо них в данную группу входят Diarthron lessertii (Wikstr.) Kit Tan, Astragalus velatus Trautv., Cousinia glandulosa Kult. и Ferula karakalensis Korovin. Только на территории Балханов встречается 8 видов: Astragalus balchanensis Boriss. (рис. 17-a), Asperula balchanica Bobrov (рис. 17-b), Acantholimon balchanicum Korovin (рис. 17-c), Cerasus turcomanica Pojark., а такие виды как Astragalus basilii Kamelin et Kovalevsk., Jurinea lydiae (рис. 17-d), Cousinia balchanica Tscherneva (рис. 17-e) и Euphorbia oidorhiza Pojark. (рис. 17-f) являются узколокальными эндемиками Малого Балхана.
Таблица 4. Соотношения типов ареалов флоры Малого Балхана
Table 4. Ratios of flora habitat types in Lesser Balkhan
|
№ |
Географические элементы |
Малый Балхан |
|
|
число видов |
% от общего числа |
||
|
I. Плюрирегиональный класс |
|||
|
1. Плюрирегиональная группа |
|||
|
I.1 |
Плюрирегиональный |
8 |
2.0 |
|
Всего |
8 |
2.0 |
|
|
II. Голарктический класс |
|||
|
2. Голарктическая группа |
|||
|
II.1 |
Голарктический |
5 |
1.2 |
|
II.2 |
Палеарктический |
8 |
2.0 |
|
Всего |
13 |
3.2 |
|
|
III. Древнесредиземноморский класс |
|||
|
3. Древнесредиземноморская группа |
|||
|
III.1 |
Древнесредиземноморский |
41 |
10.2 |
|
III.2 |
Восточносредиземноморский |
59 |
14.7 |
|
III.3 |
Понтическо-древнесредиземноморский |
9 |
2.2 |
|
III.4 |
Понтическо-восточносредиземноморский |
14 |
3.5 |
|
III.5 |
Европейско-восточносредиземноморский |
3 |
0.7 |
|
III.6 |
Восточносредиземноморско-джунгарский |
1 |
0.2 |
|
III.7 |
Европейско-древнесредиземноморский |
13 |
3.2 |
|
III.8 |
Кавказско-восточносредиземноморский |
1 |
0.2 |
|
Всего |
141 |
35.2 |
|
|
IV. Ирано-туранский класс |
|||
|
4. Пригималайская группа |
|||
|
IV.1 |
Иран-пригималайский |
1 |
0.2 |
|
Всего |
1 |
0.2 |
|
|
5. Ирано-туранская группа |
|||
|
IV.2 |
Ирано-туранский |
73 |
18.2 |
|
IV.3 |
Джунгаро-ирано-туранский |
3 |
0.7 |
|
IV.4 |
Памироалай-ирано-туранский |
3 |
0.7 |
|
IV.5 |
Понтическо-ирано-туранский |
4 |
1.0 |
|
IV.6 |
Иран-южнотуранский |
1 |
0.2 |
|
IV.7 |
Кавказо-ирано-туранский |
1 |
0.2 |
|
Всего |
85 |
21.2 |
|
|
6. Иранская группа |
|||
|
IV.8 |
Иранский |
21 |
5.2 |
|
IV.9 |
Памироалай-иранский |
1 |
0.2 |
|
IV.10 |
Ирано-тянь-шаньский |
1 |
0.2 |
|
IV.11 |
Кавказско-иранский |
3 |
0.7 |
|
IV.12 |
Центральноазиатский |
4 |
1.0 |
|
IV.13 |
Иран-центральноазиатский |
2 |
0.5 |
|
32 |
8.0 |
||
|
№ |
Географические элементы |
Малый Балхан |
|
|
число видов |
% от общего числа |
||
|
Всего |
|||
|
7. Туранская группа |
|||
|
IV.14 |
Туранский |
39 |
9.7 |
|
IV.15 |
Копетдаг-южнотуранский |
3 |
0.7 |
|
IV.16 |
Понтическо-турано-джунгарский |
2 |
0.5 |
|
IV.17 |
Турано-джунгарский |
4 |
1.0 |
|
IV.18 |
Южнотуранский |
16 |
4.0 |
|
IV.19 |
Восточногирканский |
2 |
0.5 |
|
IV.20 |
Циркумгирканский |
2 |
0.5 |
|
Всего |
68 |
17.0 |
|
|
8. Горносреднеазиатская группа |
|||
|
IV.21 |
Горносреднеазиатский |
1 |
0.2 |
|
IV.22 |
Южнотурано-горносреднеазиатский |
1 |
0.2 |
|
IV.23 |
Иран-горносреднеазиатский |
8 |
2.0 |
|
IV.24 |
Копетдаг-горносреднеазиатский |
2 |
0.5 |
|
IV.25 |
Копетдаг-памироалайский |
1 |
0.2 |
|
IV.26 |
Гиссаро-копетдагский |
1 |
0.2 |
|
IV.27 |
Копетдаг-восточногирканский |
1 |
0.2 |
|
Всего |
15 |
3.7 |
|
|
9. Копетдаг-хорасанская группа |
|||
|
IV.28 |
Копетдаг-хорасанский |
19 |
4.7 |
|
IV.29 |
Западнокопетдагский |
6 |
1.5 |
|
IV.30 |
Копетдагский |
3 |
0.7 |
|
IV.31 |
Балханский |
8 |
2.0 |
|
Всего |
36 |
9.0 |
|
|
Итого |
399 |
99.5 |
|
|
10. Дополнительно |
|||
|
V. \ |
Адвентивный |
2 |
0.5 |
|
Всего |
2 |
0.5 |
|
|
Итого |
401 |
100 |
|
Что касается Cousinia balchanica , то этот вид был впервые собран Р.П. Тарасовым в апреле 1942 года и по собранным экземплярам вид был описан в 1996 году (Cherneva, 1996). Согласно этикетке, растения собраны в 9 км юго-восточнее ж/д станции Айдин. Нам удалось обнаружить два растения в 2021 году, что является второй находкой этого редчайшего вида.
Гербарный образец: «Малый Балхан, 10 км юго-западнее ст. Перевал, скалистый склон северной экспозиции, N 39°19.265́, E 54°55.387́, 09.V.2021, А.В. Павленко (ASH)».
Второй малобалханский эндемик Euphorbia oidorhiza довольно широко распростренен в восточной части хребта и произрастает как на глинистых почвах, так и на пестроцветных обнажениях.
Третий вид – Astragalus basilii – собран в 1930 году в юго-восточных предгорьях описывамого района. Собрано лишь 2 экземпляра на 1 гербарный лист, по которому и был описан новый вид (Kamelin, Kovalevskaya, 1981). Тщательные многократные поиски по указанной в этикетке точке и на смежных участках положительных результатов не принесли.
Изученный виртуальный образец: «Западная Туркмения. Долина между Копет-Дагом и Мал[ым] Балханом, сухое русло, IV.1930, О. Ясмир (LE1274557)».
Во флоре Малого Балхана мы выделили 5 эколого-фитоценотических групп: низкотравные полусаванны, местами переходящие в крупнотравные, бедленды и пестроцветы, ущелья и каменистые скалистые склоны, предгорную полосу контакта с песчаными массивами, прибрежно-водную эвригалинную.
В первой группе низкотравные полынники, занимающие верхнюю платообразную часть главного хребта. Местами мозаично отмечаются участки крупнотравных сообществ. Только в этой группе встречается 38 видов: Eremurus kopetdaghensis Popov ex B. Fedtsch., Medicago radiata L., Leymus multicaulis (Kar. et Kir.) Tzvelev, Gentiana olivieri Griseb. и др. С полусаваннами связано еще 110 видов: Echinops ritro , Hypericum scabrum L., Asperugo procumbens L. и пр.
Бедленды и пестроцветы, подпирающие основной хребет с востока, юга и запада, представлены 47-ю видами, произрастающими только на этих участках: Asparagus brachyphyllus Turcz., Crambe edentula Fisch. et C.A. Mey. ex Korsh., Cleome quinquenervia DC., Scrophularia scoparia Pennell и многие др. Растений, распространенных и в других биотопах 111 видов: Prosopis farcta (Banks et Sol.) J.F.Macbr., Halocharis hispida (C.A. Mey.) Bunge, Cuminum setifolium (Boiss.) Koso-Pol. и др.
Каменистые ущелья и скалистые склоны расположены в основном на севере главного хребта. Исключительно в данных биотопах произрастает 35 видов: краснокнижный Malacocarpus crithmifolius (Retz.) C.A. Mey., Cousinia balchanica, Euphorbia monostyla, Marrubium anisodon K. Koch и др. Еще 92 вида встречаются как в ущельях, так и на других участках Малого Балхана: Bromus danthoniae Trin., Zygophyllum atriplicoides, Androsace maxima L., Microcephala lamellata (Bunge) Pobed. и др.
Предгорная полоса контакта с песками объединяет 108 видов, встречающихся исключительно в данной эколого-фитоценотической группе: Ephedra strobilacea Bunge, Delphinium camptocarpum Fisch. et C.A. Mey., Chrozophora gracilis Fisch. et C.A. Mey. ex Ledeb., Nonea caspica (Willd.) G. Don и др. Кроме этого с предгорьями связан еще 71 вид, произрастающий в других фитоценозах: Buglossoides tenuiflora (L. f.) I. M. Johnst., Amaranthus albus L., Arnebia decumbens (Vent.) Coss. et Kralik и др.
В последней группе растения, произрастающие по берегам немногих горных источников воды. Всего 21 вид: Alopecurus arundinaceus , Elaeagnus orientalis L., Cichorium ihtybus L. и др. Помимо этих видов имеются растения, связанные не только с прибрежными участками, но и другими фитоценотическими группами. Таких видов 12, к примеру, Fumaria parviflora Lam. связана с низкотравными полусаваннами, Tribulus terrestris L. связан с ущельями и предгорьями.
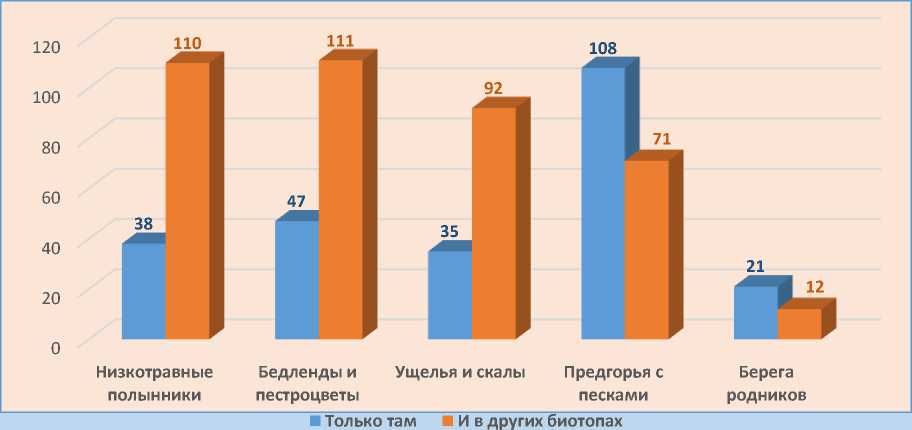
Рис. 16. Рсапределение видов по эколого-фитоценотическим группам
-
Fig. 16. Distribution of species by ecological-phytocenotic groups
Исходя из диаграммы (рис. 16), наибольшее число видов – 179 – связано с предгорной полосой тесного контакта с песками Каракумы на севере и северо-востоке Малого Балхана, что в очередной раз доказывает схожесть флор этих двух регионов. В северных предгорьях отмечаются как песчаные наносы-островки, так и непосредственный контакт с грядовобугристыми песчаными массивами.

Рис. 17. a – Astragalus balchanensis ; b – Asperula balchanica ; c – Acantholimon balchanicum ; d – Jurinea lydiae ; e – Cousinia balchanica ; f – Euphorbia oidorhiza
Fig. 17 . a – Astragalus balchanensis ; b – Asperula balchanica ; c – Acantholimon balchanicum ; d – Jurinea lydiae ; e – Cousinia balchanica ; f – Euphorbia oidorhiza
Таблица 5. Конспект флоры Малого Балхана
Table 5. Summary of the flora of the Lesser Balkhan
|
№ |
Семейства и виды |
о о |
Красная книга и эндемизм |
Жизненные формы по… |
Ареал |
н Д' |
|||
|
Серебрякову |
Раункиеру |
Вармингу |
отношению к засолению |
||||||
|
1. Equisetaceae C. Rich. ex DC. |
|||||||||
|
1. |
Equisetum ramosissimum Desf. |
ОР |
ДТП |
Геофит |
Гидрофит |
Гликофит |
II.1 |
5 |
|
|
2. Cupressaceae Bartl. |
|||||||||
|
2. \ |
Juniperus turcomanica B. Fedtsch. |
Ч |
ВД |
Фанерофит |
Мезолитофит |
Гликофит |
IV.28 |
1 |
|
|
3. Ephedraceae Dumort. |
|||||||||
|
3. |
Ephedra intermedia Schrenk et C.A. Mey. |
Ч |
БВК |
Фанерофит |
Ксерофит |
Гликофит |
IV.21 |
1-3 |
|
|
4. |
E. strobilacea Bunge |
Р |
БВК |
Фанерофит |
Ксерофит |
Гликофит |
IV.18 |
4 |
|
|
4. Colchicaceae DC. |
|||||||||
|
5. |
Colchicum robustum (Bunge) Stef. |
Р |
КлП |
Геофит |
Мезофит |
Гликофит |
IV.24 |
2 |
|
|
5. Liliaceae Juss. |
|||||||||
|
6. |
Fritillaria karelinii (Fisch. ex D. Don) Baker |
Ч |
ЛП |
Геофит |
Мезофит |
Гликофит |
IV.14 |
2 |
|
|
7. |
Gagea olgae Regel |
Р |
ЛП |
Геофит |
Мезофит |
Гликофит |
IV.8 |
1 |
|
|
8. |
G. reticulata (Pall.) Schult. et Schult. fil. |
НР |
ЛП |
Геофит |
Мезофит |
Гликофит |
III.2 |
1-4 |
|
|
9. |
G. tenuifolia (Boiss.) Fomin |
Ч |
ЛП |
Геофит |
Мезофит |
Гликофит |
III.2 |
1 |
|
|
10. |
Tulipa biflora Pall. |
ОР |
ЛП |
Геофит |
Мезофит |
Гликофит |
IV.2 |
2,4 |
|
|
11. |
T. undulatifolia var. micheliana (Hoog) Wilford |
НР |
III (VU) |
ЛП |
Геофит |
Мезофит |
Гликофит |
IV.25 |
1-2 |
|
6. Amaryllidaceae J.St.-Hil. |
|||||||||
|
12. |
Allium borszczowii Regel |
Р |
ЛП |
Геофит |
Мезофит |
Гликофит |
IV.14 |
2,4 |
|
|
13. |
A. caspium (Pall.) M. Bieb. |
Р |
ЛП |
Геофит |
Мезофит |
Гликофит |
IV.14 |
2 |
|
|
14. |
A. fibrosum Regel |
Р |
ЛП |
Геофит |
Мезофит |
Гликофит |
IV.14 |
4 |
|
|
15. |
A. rubellum M. Bieb. |
НР |
ЛП |
Геофит |
Мезофит |
Гликофит |
IV.28 |
1-4 |
|
|
16. |
A. sabulosum Steven ex Bunge |
Р |
ЛП |
Геофит |
Мезофит |
Гликофит |
IV.2 |
4 |
|
|
7. Asparagaceae Juss. |
|||||||||
|
17. |
Asparagus brachyphyllus Turcz. |
Р |
ДТП |
Геофит |
Мезоксерофит |
Галогликофит |
IV.8 |
2 |
|
|
18. |
A. breslerianus Schult. f. |
Р |
ДТП |
Геофит |
Мезоксерофит |
Галогликофит |
IV.2 |
2 |
|
|
19. |
A. persicus Baker |
ОР |
ДТП |
Геофит |
Мезоксерофит |
Гликофит |
IV.8 |
3 |
|
|
8. Asphodelaceae Juss. |
|||||||||
|
20. |
Eremurus kopetdaghensis Popov ex B. Fedtsch. |
НР |
II (EN) |
КкП |
Геофит |
Мезофит |
Гипсофит |
IV.29 |
1 |
|
9. Iridaceae Juss. |
|||||||||
|
№ |
Семейства и виды |
из н о о « |
Красная книга и эндемизм |
Жизненные формы по… |
Ареал |
н Д' |
|||
|
Серебрякову |
Раункиеру |
Вармингу |
отношению к засолению |
||||||
|
21. |
Iris drepanophylla Aitch. et Baker |
НР |
КкП |
Геофит |
Мезофит |
Гликофит |
IV.28 |
1 |
|
|
22. |
I. falcifolia Bunge |
Р |
КкП |
Геофит |
Мезофит |
Гликофит |
IV.14 |
4 |
|
|
23. |
I. longiscapa Ledeb. |
И |
КкП |
Геофит |
Мезофит |
Гликофит |
IV.14 |
1,2,4 |
|
|
10. Ixioliriaceae Nakai |
|||||||||
|
24. |
Ixiolirion tataricum (Pall.) Roem., Schult. et Schult. f. |
НР |
КлП |
Геофит |
Мезофит |
Гликофит |
III.2 |
1-2 |
|
|
11. Cyperaceae Juss. |
|||||||||
|
25. |
Bolboschoenus maritimus (L.) Palla |
ОР |
КлкП |
Гелофит |
Гигрофит |
Галогликофит |
I.1 |
5 |
|
|
26. |
Carex diluta M. Bieb. |
НР |
ДТП |
Геофит |
Гигрофит |
Гликофит |
III.2 |
5 |
|
|
27. |
C. pachystylis J. Gay |
Р |
ДТП |
Геофит |
Мезофит |
Гликофит |
III.2 |
1-2 |
|
|
28. |
C. physodes M. Bieb. |
Р |
ДТП |
Геофит |
Мезоксерофит |
Гликофит |
IV.2 |
4 |
|
|
12. Juncaceae Juss. |
|||||||||
|
29. |
Juncus articulatus L. |
Р |
ДТП |
Геофит |
Гигрофит |
Галогликофит |
I.1 |
5 |
|
|
30. |
J. bufonius L. |
Р |
ОМ |
Терофит |
Гигрофит |
Галогликофит |
I.1 |
5 |
|
|
31. |
J. maritimus Lam. |
Р |
ДТП |
Геофит |
Гигрофит |
Галофит |
III.7 |
5 |
|
|
13. Poaceae Barnhart |
|||||||||
|
32. |
Aeluropus lagopoides (L.) Thwaites |
НР |
ДТП |
Геофит |
Мезофит |
Галофит |
III.3 |
4-5 |
|
|
33. |
A. littoralis (Gouan) Parl. |
НР |
ДТП |
Геофит |
Мезофит |
Галофит |
III.3 |
4-5 |
|
|
34. |
A. pungens (M. Bieb.) K. Koch |
НР |
ДТП |
Геофит |
Мезофит |
Галофит |
III.2 |
4 |
|
|
35. |
Agropyron fragile (Roth) Candargy |
НР |
ДрТП |
Гемикриптофит |
Мезофит |
Гликофит |
IV.16 |
1 |
|
|
36. |
Alopecurus arundinaceus Poir. |
НР |
ДрТП |
Гемикриптофит |
Мезогигрофит |
Гликофит |
II.2. |
5 |
|
|
37. |
Avena barbata Pott ex Link |
НР |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.13 |
1-2 |
|
|
38. |
Bromus danthoniae Trin. |
НР |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.1 |
1-3 |
|
|
39. |
B. rubens (L.) Nevski |
НР |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.1 |
1-4 |
|
|
40. |
B. tectorum (L.) Nevski |
ОЧ |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.7 |
1-4 |
|
|
41. |
Cenchrus orientalis (Rich.) Morrone |
НР |
ДрТП |
Гемикриптофит |
Мезофит |
Гликофит |
III.2 |
1 |
|
|
42. |
Cutandia memphitica (Spreng.) Benth. |
НР |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.1 |
4 |
|
|
43. |
Cynodon dactylon (L.) Pers. |
НР |
ДТП |
Геофит |
Мезогигрофит |
Гликофит |
III.1 |
5 |
|
|
44. |
Eremopyrum bonaepartis (Spreng.) Nevski |
НР |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.2 |
1-4 |
|
|
45. |
E. distans (K. Koch) Nevski |
НР |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.1 |
1-4 |
|
|
46. |
E. orientale (L.) Jaub. et Spach |
Ч |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.3 |
1-4 |
|
|
47. |
Hordeum leporinum Link |
Р |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.3 |
5 |
|
|
48. |
Leymus multicaulis (Kar. et Kir.) Tzvelev |
НР |
ДрТП |
Гемикриптофит |
Мезофит |
Гликофит |
IV.16 |
1 |
|
|
49. |
Phragmites australis (Cav.) Trin. ex Steud. |
НР |
ДТП |
Геофит |
Гигрофит |
Галогликофит |
I.1 |
5 |
|
|
50. |
Poa bulbosa L. |
ОЧ |
ДрТП |
Гемикриптофит |
Мезофит |
Гликофит |
III.3 |
1-3 |
|
|
№ |
Семейства и виды |
н о о « рц |
Красная книга и эндемизм |
Жизненные формы по… |
Ареал |
н Д' |
|||
|
Серебрякову |
Раункиеру |
Вармингу |
отношению к засолению |
||||||
|
51. |
Schismus arabicus Nees |
НР |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.2 |
4 |
|
|
52. |
Stipa caucasica Schmalh. |
НР |
ДрТП |
Гемикриптофит |
Мезофит |
Гликофит |
III.2 |
1 |
|
|
53. |
Stipagrostis karelinii (Trin. et Rupr.) Tzvelev |
Р |
ДрТП |
Гемикриптофит |
Мезофит |
Гликофит |
IV.2 |
4 |
|
|
54. |
S. pennata (Trin.) De Winter |
НР |
ДрТП |
Гемикриптофит |
Мезофит |
Гликофит |
IV.2 |
4 |
|
|
55. |
S. plumosa ( L.) Munro ex T.Anderson |
Р |
ДрТП |
Гемикриптофит |
Мезофит |
Гликофит |
III.2 |
4 |
|
|
56. |
Trisetaria loeflingiana (L.) Paunero |
НР |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.7 |
4 |
|
|
57. |
Vulpia persica (Boiss. et Buhse) V.I. Krecz. et Bobrov |
Р |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.8 |
1,3 |
|
|
14. Berberidaceae Juss. |
|||||||||
|
58. |
Bongardia chrysogonum (L.) Spach |
Ч |
КлбП |
Геофит |
Мезофит |
Гликофит |
III.2 |
1-2 |
|
|
59. |
Leontice ewersmannii Bunge |
НР |
КлбП |
Геофит |
Мезофит |
Гликофит |
IV.8 |
1-2 |
|
|
15. Papaveraceae Juss. |
|||||||||
|
60. |
Fumaria parviflora Lam. |
Р |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.1 |
1,5 |
|
|
61. |
F. vaillantii Loisel. |
Р |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.1 |
1,5 |
|
|
62. |
Glaucium elegans Fisch. et C.A. Mey. |
НР |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.8 |
2 |
|
|
63. |
G. oxylobum Boiss. et Buhse |
НР |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.28 |
2 |
|
|
64. |
Hypecoum parviflorum Kar. et Kir. |
НР |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.14 |
1,2,4 |
|
|
65. |
H. pendulum L. |
НР |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.1 |
1,2,4 |
|
|
66. |
H. trilobum Trautv. |
Ч |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.12 |
1 |
|
|
67. |
Papaver litwinowii Fedde ex Popov |
НР |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.23 |
1,3 |
|
|
68. |
P. pavoninum Schrenk |
Ч |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.8 |
1-4 |
|
|
69. |
Roemeria hybrida (L.) DC. |
Ч |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.2 |
1-4 |
|
|
70. |
R. refracta DC. |
ОЧ |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.8 |
1-4 |
|
|
16. Ranunculaceae Juss. |
|||||||||
|
71. |
Adonis bienertii Butkov |
Р |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.11 |
1 |
|
|
72. |
Anemone petiolulosa Juz. |
Ч |
КлбП |
Геофит |
Мезофит |
Гликофит |
IV.26 |
1-2 |
|
|
73. |
Delphinium camptocarpum Fisch. et C.A. Mey. |
НР |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.2 |
4 |
|
|
74. |
Ranunculus falcatus L. |
ОЧ |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.7 |
1-4 |
|
|
17. Geraniaceae Juss. |
|||||||||
|
75. |
Erodium ciconium (L.) L'Her. |
Ч |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.1 |
1-2 |
|
|
76. |
E. cicutarium (L.) L'Her. |
ОЧ |
ОМ |
Терофит |
Мезофит |
Гликофит |
II.2 |
1-2 |
|
|
77. |
E. litwinowii Woronow |
НЧ |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.13 |
1-2 |
|
|
78. |
E. oxyrhynchum M. Bieb. |
Ч |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.2 |
1-2,4 |
|
|
79. |
Geranium rotundifolium L. |
НР |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.7 |
3 |
|
|
18. Biebersteiniaceae Agardh |
|||||||||
|
№ |
Семейства и виды |
о о рц |
Красная книга и эндемизм |
Жизненные формы по… |
Ареал |
н Д' |
|||
|
Серебрякову |
Раункиеру |
Вармингу |
отношению к засолению |
||||||
|
80. |
Biebersteinia multifida DC. |
НЧ |
КлбП |
Геофит |
Мезоксерофит |
Гипсофит |
IV.8 |
2 |
|
|
19. Nitrariaceae Lindl. |
|||||||||
|
81. |
Malacocarpus crithmifolius (Retz.) C.A. Mey. |
Р |
III (VU) |
КчН |
Фанерофит |
Мезофит |
Галогликофит |
IV.2 |
3 |
|
82. |
Nitraria schoberi L. |
Р |
КН |
Фанерофит |
Ксерофит |
Галофит |
IV.5 |
4 |
|
|
83. |
Peganum harmala L. |
ОЧ |
СП |
Гемикриптофит |
Мезоксерофит |
Галогликофит |
III.3 |
4 |
|
|
84. |
Tetradiclis tenella (Ehrenb.) Litv . |
Р |
ОС |
Терофит |
Ксерофит |
Галофит |
III.3 |
4 |
|
|
20. Rutaceae Juss. |
|||||||||
|
85. |
Haplophyllum bungei Trautv. |
Р |
СП |
Гемикриптофит |
Мезоксерофит |
Гликофит |
IV.2 |
4 |
|
|
86. |
H. obtusifolium (Ledeb.) Ledeb. |
Ч |
ПкчН |
Хамефит |
Мезоксерофит |
Галогликофит |
IV.14 |
2,4 |
|
|
21. Thymelaeaceae Juss. |
|||||||||
|
87. |
Diarthron vesiculosum (Fisch. et C.A. Mey. ex Kar. et Kir.) C.A. Mey. |
Ч |
ОМ |
Терофит |
Мезоксерофит |
Галогликофит |
III.2 |
1-4 |
|
|
88. |
D. lessertii (Wikstr.) Kit Tan |
Р |
КН |
Фанерофит |
Мезоксерофит |
Гипсофит |
IV.29 |
3 |
|
|
22. Brassicaceae Burnett. |
|||||||||
|
89. |
Aethionema carnea (Banks et Sol.) B. Fedtsch. |
Р |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.23 |
3 |
|
|
90. |
Alyssum dasycarpum Stephan ex Willd. |
НР |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.23 |
2,4 |
|
|
91. |
A. szovitsianum Fisch. et C.A. Mey. |
НР |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.2 |
1 |
|
|
92. |
A. turkestanicum Regel et Schmalh. |
Ч |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.2 |
4 |
|
|
93. |
Capsella bursa-pastoris (L.) Medik. |
НР |
ОМ |
Терофит |
Мезофит |
Гликофит |
I.1 |
1 |
|
|
94. |
Clypeola jonthlaspi L . |
НР |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.1 |
1-3 |
|
|
95. |
Crambe edentula Fisch. et C.A. Mey. ex Korsh. |
НР |
СП |
Гемикриптофит |
Мезофит |
Гипсофит |
IV.14 |
2 |
|
|
96. |
Cymatocarpus pilosissimus (Trautv.) O.E.Schulz |
Р |
ОМ |
Терофит |
Мезофит |
Гипсофит |
IV.15 |
2 |
|
|
97. |
Descurainia sophia (L.) Webb ex Prantl |
Р |
ОМ |
Терофит |
Мезофит |
Гликофит |
II.2 |
1 |
|
|
98. |
Diptychocarpus strictus (Fisch. ex M. Bieb.) Trautv. |
НЧ |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.2 |
1 |
|
|
99. |
Draba verna L. |
Ч |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.1 |
3 |
|
|
100. |
Eruca sativa Mill. |
НР |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.1 |
1-3 |
|
|
101. |
Euclidium syriacum (L.) W. T. Aiton |
НР |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.4 |
1 |
|
|
102. |
Goldbachia laevigata (M. Bieb.) DC. |
НР |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.2 |
2 |
|
|
103. |
G. torulosa DC. |
НЧ |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.2 |
2 |
|
|
104. |
Hornungia procumbens (L.) Hayek |
Р |
ОМ |
Терофит |
Мезоксерофит |
Галогликофит |
III.1 |
4 |
|
|
105. |
Isatis emarginata Kar. et Kir. |
Р |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.2 |
4 |
|
|
106. |
I. minima Bunge |
Р |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.2 |
4 |
|
|
107. |
I. trachycarpa Trautv. |
НЧ |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.2 |
3-4 |
|
|
108. |
I. violascens Bunge |
НР |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.14 |
4 |
|
|
№ |
Семейства и виды |
из н о о « |
Красная книга и эндемизм |
Жизненные формы по… |
Ареал |
н Д' |
|||
|
Серебрякову |
Раункиеру |
Вармингу |
отношению к засолению |
||||||
|
109. |
Lachnoloma lehmannii Bunge |
Р |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.14 |
4 |
|
|
110. |
L. perfoliatum L. |
Ч |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.1 |
1-4 |
|
|
111. |
Leptaleum filifolium (Willd.) DC. |
Ч |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.2 |
1-4 |
|
|
112. |
Litwinowia tenuissima (Pall.) Woronow ex Pavlov |
Р |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.5 |
1 |
|
|
113. |
Matthiola chenopodiifolia Fisch. et C.A. Mey. |
Р |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.18 |
4 |
|
|
114. |
M. farinosa Bunge |
НР |
СП |
Гемикриптофит |
Мезоксерофит |
Гипсофит |
IV.8 |
2 |
|
|
115. |
Meniocus linifolius (Stephan) DC. |
Ч |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.5 |
4 |
|
|
116. |
Neotorularia torulosa (Desf.) Hedge et J. Leonard |
НР |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.1 |
1-2 |
|
|
117. |
Olimarabidopsis pumila (Stephan ex Willd.) Al-Shehbaz, oKane et R.A. Price |
НР |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.2 |
1,3 |
|
|
118. |
Rapistrum rugosum (L.) All. |
НР |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.1 |
1-4 |
|
|
119. |
Sameraria armena Desv. |
НЧ |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.2 |
2 |
|
|
120. |
Spirorhynchus sabulosus Kar. et Kir. |
ОР |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.14 |
4 |
|
|
121. |
Streptoloma desertorum Bunge |
НЧ |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.14 |
2,4 |
|
|
122. |
Strigosella africana (L.) Botsch. |
НР |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.1 |
4 |
|
|
123. |
S. circinnata (Bunge) Botsch. |
НР |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.14 |
4 |
|
|
124. |
S. grandiflora (Bunge) Botsch. |
Р |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.2 |
1-4 |
|
|
125. |
S. intermedia (C.A. Mey.) Botsch. |
Р |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.14 |
1 |
|
|
126. |
Tetracme recurvata Bunge |
Р |
ОМ |
Терофит |
Мезоксерофит |
Гликофит |
IV.14 |
4 |
|
|
23. Capparaceae Juss. |
|||||||||
|
127. Capparis spinosa var. herbacea (Willd.) Fici |
Ч |
ПкчН |
Хамефит |
Мезоксерофит |
Гипсофит |
III.2 |
2-3 |
||
|
24 |
. Cleomaceae Bercht. et J. Presl |
||||||||
|
128. |
Cleome quinquenervia DC. |
ОР |
ОМ |
Терофит |
Мезоксерофит |
Гипсофит |
IV.14 |
2 |
|
|
129. |
C. turkmena Bobr. |
Р |
IV |
ОМ |
Терофит |
Мезоксерофит |
Гипсофит |
IV.29 |
2 |
|
25. Fabaceae Lindl. |
|||||||||
|
130. |
Alhagi pseudalhagi subsp. persarum (Boiss. et Buhse) Takht. |
Ч |
КотП |
Гемикриптофит |
Ксерофит |
Галогликофит |
IV.2 |
4 |
|
|
131. |
Ammodendron conollyi Bunge ex Boiss. |
НЧ |
ЛД |
Фанерофит |
Мезофит |
Гликофит |
IV.14 |
4 |
|
|
132. |
Astragalus ammodendron Bunge |
НЧ |
ПкН |
Хамефит |
Мезоксерофит |
Гликофит |
IV.14 |
4 |
|
|
133. |
A. ammophilus Kar. et Kir. |
НЧ |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.2 |
2,4 |
|
|
134. |
A. arpilobus Kar. et Kir. |
Ч |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.1 |
4 |
|
|
135. |
A. balchanensis Boriss. |
НР |
эндемик Б. |
СП |
Гемикриптофит |
Мезофит |
Гликофит |
IV.31 |
1,3 |
|
136. |
A. basilii Kamelin et Kovalevsk. |
ОР |
эндемик Б. |
СП |
Гемикриптофит |
Мезофит |
Гликофит |
IV.31 |
4 |
|
137. |
A. campylorhynchus Fisch. et C.A. Mey. |
НР |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.2 |
1-4 |
|
|
138. |
A. chiwensis Bunge |
НР |
СП |
Гемикриптофит |
Мезофит |
Гликофит |
IV.18 |
4 |
|
|
№ |
Семейства и виды |
из н о о « |
Красная книга и эндемизм |
Жизненные формы по… |
Ареал |
н Д' |
|||
|
Серебрякову |
Раункиеру |
Вармингу |
отношению к засолению |
||||||
|
139. |
A. cornubovis Lipsky |
НЧ |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.2 |
1,3 |
|
|
140. |
A. crenatus Schult. |
Р |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.23 |
1 |
|
|
141. |
A. curvipes Trautv. |
НР |
СП |
Гемикриптофит |
Мезофит |
Гипсофит |
IV.30 |
2-3 |
|
|
142. |
A. drobovii Popov et Vved. |
НЧ |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.18 |
4 |
|
|
143. |
A. flexus Fisch. |
НР |
СП |
Гемикриптофит |
Мезофит |
Гликофит |
IV.2 |
4 |
|
|
144. |
A. nephtonensis Freyn |
НР |
эндемик КХ |
СП |
Гемикриптофит |
Мезофит |
Гликофит |
IV.28 |
1-2 |
|
145. |
A. oxyglottis Steven ex M. Bieb. |
НЧ |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.5 |
1-4 |
|
|
146. |
A. rustamovii A.V. Pavlenko et Laktionov |
Р |
СП |
Гемикриптофит |
Мезофит |
Гликофит |
IV.18 |
4 |
|
|
147. |
A. turcomanicus Bunge |
НР |
ПкчН |
Хамефит |
Мезоксерофит |
Гликофит |
IV.19 |
2,4 |
|
|
148. |
A. velatus Trautv. |
Ч |
эндемик КХ |
СП |
Гемикриптофит |
Мезофит |
Гликофит |
IV.29 |
1 |
|
149. |
A. verus Olivier |
НЧ |
эндемик КХ |
ПКч |
Фанерофит |
Мезоксерофит |
Гликофит |
IV.28 |
1-2 |
|
150. |
A. winkleri Trautv. |
НР |
эндемик ЮТ |
СП |
Гемикриптофит |
Мезофит |
Гликофит |
IV.18 |
4 |
|
151. |
A. xiphidioides Freyn et Sint. |
НР |
СП |
Гемикриптофит |
Мезофит |
Гликофит |
IV.15 |
1-4 |
|
|
152. |
Caragana grandiflora (M. Bieb.) DC. |
Ч |
КчН |
Фанерофит |
Мезофит |
Гликофит |
IV.20 |
2 |
|
|
153. |
Glycyrrhiza glabra L. |
НР |
ДТП |
Геофит |
Мезофит |
Гликофит |
III.4 |
4-5 |
|
|
154. |
G. triphylla Fisch. et C.A. Mey. |
Р |
СП |
Гемикриптофит |
Мезоксерофит |
Гликофит |
IV.15 |
4 |
|
|
155. |
Hedysarum micropterum Bunge ex Boiss. |
НЧ |
СП |
Гемикриптофит |
Мезофит |
Гликофит |
IV.28 |
1-3 |
|
|
156. |
Medicago radiata L. |
Р |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.2 |
1 |
|
|
157. |
Prosopis farcta (Banks et Sol.) J.F.Macbr. |
НЧ |
КчН |
Фанерофит |
Мезоксерофит |
Гликофит |
III.2 |
2,4 |
|
|
26. Elaeagnaceae Juss. |
|||||||||
|
158. |
Elaeagnus orientalis L. |
ОР |
ЛД |
Фанерофит |
Мезофит |
Гликофит |
III.1 |
5 |
|
|
27. Rhamnaceae Juss. |
|||||||||
|
159. |
Rhamnus songorica Gontsch. |
НР |
КН |
Фанерофит |
Мезоксерофит |
Гликофит |
IV.19 |
3 |
|
|
28. Rosaceae Juss. |
|||||||||
|
160. |
Amygdalus turcomanica Lincz. |
Р |
КчН |
Фанерофит |
Мезофит |
Гликофит |
IV.23 |
3 |
|
|
161. |
Cerasus turcomanica Pojark. |
Р |
эндемик Б. |
КчН |
Фанерофит |
Мезоксерофит |
Гликофит |
IV.31 |
3 |
|
162. |
Prunus pseudoprostratus (Pojark.) Rech. f. |
НР |
КчН |
Фанерофит |
Мезоксерофит |
Гликофит |
IV.8 |
3 |
|
|
29. Urticaceae Juss. |
|||||||||
|
163. |
Parietaria judaica L . |
НЧ |
КТП |
Гемикриптофит |
Мезолитофит |
Гликофит |
III.2 |
3 |
|
|
30. Euphorbiaceae Juss. |
|||||||||
|
164. |
Chrozophora gracilis Fisch. et C.A. Mey. ex Ledeb. |
Р |
ОМ |
Терофит |
Мезоксерофит |
Гликофит |
IV.2 |
4 |
|
|
165. |
Euphorbia aserbajdzhanica Bordz. |
Р |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.2 |
1 |
|
|
166. |
E. bungei Boiss. |
Р |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.11 |
2 |
|
|
№ |
Семейства и виды |
из н о о « |
Красная книга и эндемизм |
Жизненные формы по… |
Ареал |
н Д' |
|||
|
Серебрякову |
Раункиеру |
Вармингу |
отношению к засолению |
||||||
|
167. |
E. chamaesyce L. |
НР |
ОМ |
Терофит |
Мезоксерофит |
Гликофит |
III.2 |
4 |
|
|
168. |
E. cheirolepis Fisch. et C.A. Mey. ex Ledeb. |
Р |
ОМ |
Терофит |
Мезоксерофит |
Гликофит |
IV.2 |
4 |
|
|
169. |
E. consanguinea Schrenk ex Fisch. et C.A. Mey. |
Р |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.14 |
1-2 |
|
|
170. |
E. densa Schrenk |
Р |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.2 |
3 |
|
|
171. |
E. granulata Forssk. |
НР |
ОМ |
Терофит |
Мезоксерофит |
Гликофит |
IV.2 |
1-2 |
|
|
172. |
E. humilis Ledeb. |
Р |
СП |
Гемикриптофит |
Мезофит |
Гликофит |
IV.17 |
1-2 |
|
|
173. |
E. inderiensis Less. ex Kar. et Kir. |
Р |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.3 |
2 |
|
|
174. |
E. monostyla Prokh. |
ОЧ |
СП |
Гемикриптофит |
Мезофит |
Гликофит |
IV.28 |
3 |
|
|
175. |
E. oidorhiza Pojark. |
НР |
IV |
КлбП |
Геофит |
Мезофит |
Гликофит |
IV.31 |
2 |
|
176. |
E. turczaninowii Kar. et Kir. |
НР |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.17 |
4 |
|
|
31. Hypericaceae Juss. |
|||||||||
|
177. Hypericum scabrum L. |
НЧ |
СП |
Гемикриптофит |
Мезофит |
Гликофит |
III.2 |
1,3 |
||
|
32. Linaceae S.F.Gray |
|||||||||
|
178. Linum corymbulosum Rchb. |
Р |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.1 |
1 |
||
|
33. Phyllanthaceae Martinov |
|||||||||
|
179. Andrachne telephioides L. |
НР |
ПкчН |
Хамефит |
Мезоксерофит |
Гипсофит |
III.2 |
3 |
||
|
34. Salicaceae Mirb. |
|||||||||
|
180. Populus euphratica Olivier |
Р |
ЛД |
Фанерофит |
Мезофит |
Галогликофит |
III.2 |
5 |
||
|
35. Zygophyllaceae R. Br. |
|||||||||
|
181. |
Tribulus terrestris L. |
Ч |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.1 |
3-5 |
|
|
182. |
Zygophyllum atriplicoides Fisch. et C.A. Mey. |
Ч |
КН |
Фанерофит |
Мезофит |
Гликофит |
IV.8 |
2-3 |
|
|
183. |
Z. eichwaldii C.A. Mey. |
НР |
ПкчН |
Хамефит |
Мезофит |
Гипсофит |
IV.18 |
4 |
|
|
184. |
Z. fabago L . |
Р |
СП |
Гемикриптофит |
Мезофит |
Гликофит |
III.5 |
5 |
|
|
185. |
Z. jaxarticum Popov |
ОР |
СП |
Гемикриптофит |
Мезофит |
Гипсофит |
IV.12 |
5 |
|
|
186. |
Z. macrophyllum Regel et Schmalh. |
Р |
СП |
Гемикриптофит |
Мезофит |
Гипсофит |
IV.14 |
2 |
|
|
187. |
Z. turcomanicum Fisch. ex Bunge |
Р |
СП |
Гемикриптофит |
Мезоксерофит |
Гипсофит |
IV.14 |
2 |
|
|
36. Amaranthaceae Juss. |
|||||||||
|
188. |
Agriophyllum latifolium Fisch. et C.A. Mey. |
НЧ |
ОМ |
Терофит |
Ксерофит |
Гликофит |
IV.2 |
4 |
|
|
189. |
Amaranthus albus L. |
НР |
ОМ |
Терофит |
Мезофит |
Гликофит |
V. |
1-4 |
|
|
190. |
Amaranthus blitoides S. Watson |
Р |
ОМ |
Терофит |
Мезофит |
Гликофит |
V. |
5 |
|
|
191. |
Anabasis eriopoda (Schrenk) Benth. ex Volkens |
НЧ |
ПкчС |
Хамефит |
Ксерофит |
Гипсофит |
IV.2 |
4 |
|
|
192. |
A. truncata (Schrenk) Bunge |
Р |
СП |
Гемикриптофит |
Ксерофит |
Гипсофит |
IV.17 |
4 |
|
|
193. |
Anthochlamys turcomanica Iljin |
НЧ |
ОМ |
Терофит |
Ксерофит |
Гипсофит |
IV.28 |
2 |
|
|
194. |
Atriplex dimorphostegia Kar. et Kir. |
Р |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.2 |
4 |
|
|
№ |
Семейства и виды |
н о о « рц |
Красная книга и эндемизм |
Жизненные формы по… |
Ареал |
н Д' |
|||
|
Серебрякову |
Раункиеру |
Вармингу |
отношению к засолению |
||||||
|
195. |
A. lehmanniana Bunge |
НЧ |
ПкчН |
Хамефит |
Мезоксерофит |
Галогликофит |
III.2 |
2 |
|
|
196. |
Bienertia cycloptera Bunge |
Р |
ОС |
Терофит |
Ксерофит |
Галофит |
IV.2 |
2 |
|
|
197. |
Caroxylon dendroides (Pall.) Tzvelev |
НР |
ПкчН |
Хамефит |
Ксерофит |
Галогликофит |
III.2 |
4 |
|
|
198. |
C. gemmascens (Pall.) Tzvelev |
НР |
ПкчН |
Хамефит |
Ксерофит |
Гипсофит |
IV.2 |
4 |
|
|
199. |
C. incanescens (C.A. Mey.) Akhani et Roalson |
НР |
ОМ |
Терофит |
Ксерофит |
Галогликофит |
IV.2 |
2,4 |
|
|
200. |
C. orientale (S.G. Gmel.) Tzvelev |
НР |
ПкчН |
Хамефит |
Ксерофит |
Галогликофит |
IV.12 |
4 |
|
|
201. |
C. scleranthum (C.A. Mey.) Akhani et Roalson |
НР |
ОМ |
Терофит |
Ксерофит |
Галогликофит |
IV.2 |
4 |
|
|
202. |
C. turkestanicum (Litv.) Akhani et Roalson |
Ч |
ОМ |
Терофит |
Ксерофит |
Галогликофит |
IV.2 |
1-4 |
|
|
203. |
Ceratocarpus arenarius L. |
Ч |
ОМ |
Терофит |
Ксерофит |
Галогликофит |
III.4 |
4 |
|
|
204. |
Chenopodium album L. |
Ч |
ОМ |
Терофит |
Мезофит |
Галогликофит |
II.1 |
1 |
|
|
205. |
Climacoptera lanata (Pall.) Botsch . |
ОЧ |
ОМ |
Терофит |
Ксерофит |
Галофит |
IV.2 |
1-4 |
|
|
206. |
C. ferganica (Drobow) Botsch. |
НР |
ОМ |
Терофит |
Ксерофит |
Галофит |
IV.18 |
4 |
|
|
207. |
C. turcomanica (Litv.) Botsch. |
НЧ |
ОМ |
Терофит |
Ксерофит |
Галофит |
IV.18 |
4 |
|
|
208. |
Corispermum lehmannianum Bunge |
Р |
ОМ |
Терофит |
Ксерофит |
Галогликофит |
IV.14 |
4 |
|
|
209. |
Girgensohnia oppositiflora (Pall.) Fenzl |
Р |
ОМ |
Терофит |
Ксерофит |
Галогликофит |
IV.2 |
3,4 |
|
|
210. |
Halimocnemis karelinii Moq. |
НЧ |
ОМ |
Терофит |
Ксерофит |
Галофит |
IV.14 |
4 |
|
|
211. |
H. longifolia Bunge |
НЧ |
ОМ |
Терофит |
Ксерофит |
Галогликофит |
IV.18 |
2 |
|
|
212. |
Halocharis hispida (C.A. Mey.) Bunge |
НР |
ОМ |
Терофит |
Ксерофит |
Галогликофит |
IV.2 |
2,4 |
|
|
213. |
Halocnemum strobilaceum (Pall.) M. Bieb. |
НР |
ПкчС |
Хамефит |
Ксерофит |
Галофит |
III.1 |
2,4 |
|
|
214. |
Halostachys caspica (M. Bieb.) C.A. Mey. |
НЧ |
КС |
Фанерофит |
Ксерофит |
Галофит |
III.4 |
2,4 |
|
|
215. |
Halothamnus glaucus (M. Bieb.) Botsch. |
Ч |
ПкчС |
Хамефит |
Ксерофит |
Галогликофит |
III.2 |
2 |
|
|
216. |
H. subaphyllus (C.A. Mey.) Botsch. |
НР |
ПкчС |
Хамефит |
Ксерофит |
Галогликофит |
IV.14 |
4 |
|
|
217. |
Haloxylon ammodendron (C.A. Mey.) Bunge ex Fenzl |
Ч |
КН |
Фанерофит |
Ксерофит |
Галогликофит |
IV.2 |
4 |
|
|
218. |
Horaninovia anomala (C.A. Mey.) Moq. |
НЧ |
ОМ |
Терофит |
Ксерофит |
Гликофит |
IV.3 |
4 |
|
|
219. |
Kalidium caspicum (L.) Ung.-Sternb. |
Р |
ПкчС |
Хамефит |
Ксерофит |
Галофит |
III.4 |
4 |
|
|
220. |
Kaviria aucheri (Moq.) Akhani |
НЧ |
ПкчС |
Хамефит |
Ксерофит |
Гипсофит |
IV.28 |
1-2 |
|
|
221. |
K. gossypina (Bunge ex Boiss.) Akhani |
НР |
ОМ |
Терофит |
Ксерофит |
Галогликофит |
IV.2 |
2,4 |
|
|
222. |
K. tomentosa (Moq.) Akhani |
Р |
ПкчН |
Хамефит |
Ксерофит |
Гипсофит |
III.2 |
2-3 |
|
|
223. |
Kochia odontoptera Schrenk |
Р |
ОМ |
Терофит |
Ксерофит |
Гликофит |
IV.14 |
4 |
|
|
224. |
Londesia eriantha Fisch. et C.A. Mey. |
НЧ |
ОМ |
Терофит |
Ксерофит |
Галогликофит |
IV.2 |
4 |
|
|
225. |
Noaea mucronata (Forssk.) Asch. et C. Schweinf. |
НЧ |
ПкчН |
Хамефит |
Ксерофит |
Галогликофит |
III.2 |
1-3 |
|
|
226. |
Petrosimonia glauca (Pall.) Bunge |
НР |
ОМ |
Терофит |
Ксерофит |
Галогликофит |
IV.2 |
2 |
|
|
227. |
Salsola arbusculiformis Drobow |
Р |
ПкчС |
Хамефит |
Ксерофит |
Галогликофит |
IV.2 |
2 |
|
|
228. |
Spinacia turkestanica Iljin |
НР |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.2 |
1-2 |
|
|
№ |
Семейства и виды |
из н о о « |
Красная книга и эндемизм |
Жизненные формы по… |
Ареал |
н Д' |
|||
|
Серебрякову |
Раункиеру |
Вармингу |
отношению к засолению |
||||||
|
229. |
Suaeda microphylla Pall. |
НР |
ПкчС |
Хамефит |
Ксерофит |
Галофит |
IV.2 |
4 |
|
|
230. |
Xylosalsola arbuscula (Pall.) Tzvelev |
НР |
ПкчС |
Хамефит |
Ксерофит |
Галогликофит |
IV.3 |
1-2,4 |
|
|
231. |
X. richteri (Moq.) Akhani et Roalson |
НР |
КН |
Фанерофит |
Ксерофит |
Галогликофит |
IV.2 |
4 |
|
|
37. Caryophyllaceae Juss. |
|||||||||
|
232. |
Acanthophyllum elatius Bunge |
Р |
ПкчН |
Хамефит |
Ксерофит |
Гликофит |
IV.18 |
4 |
|
|
233. |
A. pungens (Bunge) Boiss. |
НЧ |
ППкч |
Хамефит |
Ксерофит |
Гликофит |
IV.17 |
1-2,4 |
|
|
234. |
A. stenostegium Freyn |
ППкч |
Хамефит |
Ксерофит |
Гликофит |
IV.9 |
1-2,4 |
||
|
235. |
Gypsophila heteropoda Freyn et Sint. |
НЧ |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.8 |
4 |
|
|
236. |
G. pilosa Huds. |
НЧ |
ОМ |
Терофит |
Мезофит |
Гипсофит |
IV.8 |
2,4 |
|
|
237. |
Herniaria hirsuta L. |
НЧ |
ОМ |
Терофит |
Мезоксерофит |
Гликофит |
III.7 |
2,4 |
|
|
238. |
Holosteum umbellatum L. |
ОЧ |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.1 |
1-4 |
|
|
239. |
Minuartia meyeri (Boiss.) Bornm. |
Ч |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.2 |
3 |
|
|
240. |
Silene conoidea L. |
НЧ |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.2 |
1-3 |
|
|
241. |
S. nana Kar. et Kir. |
НЧ |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.12 |
4 |
|
|
242. |
Spergularia diandra ( Guss.) Heldr. et Sart. |
НР |
ОМ |
Терофит |
Мезоксерофит |
Галогликофит |
III.4 |
2,4 |
|
|
38. Frankeniaceae S. F. Gray |
|||||||||
|
243. |
Frankenia hirsuta L. |
НР |
СП |
Гемикриптофит |
Ксерофит |
Галофит |
III.2 |
4 |
|
|
39. Plumbaginaceae Lincz. |
|||||||||
|
244. |
Acantholimon balchanicum Korovin |
НЧ |
эндемик Б. |
ППкч |
Хамефит |
Ксерофит |
Гликофит |
IV.31 |
3 |
|
245. |
Limonium reniforme (Girard) Lincz. |
Р |
СП |
Гемикриптофит |
Мезоксерофит |
Галогликофит |
IV.4 |
4 |
|
|
246. |
L. suffruticosum (L.) Kuntze |
НР |
ПкчН |
Хамефит |
Ксерофит |
Галофит |
IV.2 |
2,4 |
|
|
40. Polygonaceae Juss. |
|||||||||
|
247. |
Atraphaxis spinosa L. |
НР |
КН |
Фанерофит |
Мезоксерофит |
Галогликофит |
III.4 |
1-4 |
|
|
248. |
Calligonum caput-medusae Schrenk |
Р |
КН |
Фанерофит |
Ксерофит |
Гликофит |
IV.14 |
4 |
|
|
249. |
C. junceum (Fisch. et C.A. Mey.) Litv. |
НЧ |
ПкчН |
Хамефит |
Ксерофит |
Гипсофит |
IV.14 |
3-4 |
|
|
250. |
C. leucocladum (Schrenk) Bunge |
Р |
КчН |
Фанерофит |
Ксерофит |
Гликофит |
IV.14 |
4 |
|
|
251. |
C. macrocarpum I. G. Borshch |
Р |
КН |
Фанерофит |
Ксерофит |
Гликофит |
IV.14 |
4 |
|
|
252. |
C. microcarpum I. G. Borshch. |
НР |
КчН |
Фанерофит |
Ксерофит |
Гликофит |
IV.14 |
4 |
|
|
253. |
C. triste Litv. |
ОР |
I (CR) |
КчН |
Фанерофит |
Ксерофит |
Гликофит |
IV.14 |
4 |
|
254. |
Polygonum aviculare L. |
НР |
ОМ |
Терофит |
Мезоксерофит |
Галогликофит |
II.1 |
1-4 |
|
|
255. |
P. pseudoarenarium Klokov |
P |
ОМ |
Терофит |
Мезогигрофит |
Гликофит |
III.7 |
1-4 |
|
|
256. |
Rheum turkestanicum Janisch. |
НЧ |
СП |
Гемикриптофит |
Мезофит |
Гликофит |
IV.22 |
1-4 |
|
|
257. |
Rumex chalepensis Mill. |
Р |
СП |
Гемикриптофит |
Мезофит |
Гликофит |
III.2 |
3 |
|
|
41. Tamaricaceae Link |
|||||||||
|
№ |
Семейства и виды |
из н о о « |
Красная книга и эндемизм |
Жизненные формы по… |
Ареал |
н Д' |
|||
|
Серебрякову |
Раункиеру |
Вармингу |
отношению к засолению |
||||||
|
258. |
Reaumuria fruticosa Bunge |
НР |
КН |
Фанерофит |
Ксерофит |
Галофит |
IV.6 |
4 |
|
|
259. |
R. oxiana (Ledeb.) Boiss . |
Р |
ПкчН |
Хамефит |
Ксерофит |
Галофит |
IV.2 |
4 |
|
|
260. |
R. reflexa Lipsky |
НЧ |
ПкчН |
Хамефит |
Мезоксерофит |
Галогликофит |
IV.24 |
3 |
|
|
261. |
R. turkestanica Gorschk. |
НР |
ПкчН |
Хамефит |
Мезоксерофит |
Гликофит |
IV.2 |
4 |
|
|
262. |
Tamarix florida Bunge |
НЧ |
КН |
Фанерофит |
Ксерофит |
Галогликофит |
III.1 |
2,4-5 |
|
|
263. |
T. ramosissima Ledeb. |
НР |
КН |
Фанерофит |
Ксерофит |
Галогликофит |
III.4 |
2,4-5 |
|
|
42. Primulaceae Vent. |
|||||||||
|
264. |
Androsace maxima L. |
Ч |
ОМ |
Терофит |
Мезофит |
Гликофит |
I.1 |
1-3 |
|
|
43. Apocynaceae Juss. |
|||||||||
|
265. |
Cynanchum acutum L. |
НЧ |
СП |
Гемикриптофит |
Мезофит |
Гликофит |
II.2 |
3 |
|
|
266. |
Trachomitum scabrum (Russanov) Pobed. |
НР |
ДТП |
Геофит |
Мезофит |
Галогликофит |
IV.2 |
3,5 |
|
|
267. |
Vincetoxicum pumilum Decaisne |
НЧ |
КТП |
Гемикриптофит |
Мезолитофит |
Гликофит |
III.2 |
3 |
|
|
44. Gentianaceae Juss. |
|||||||||
|
268. Gentiana olivieri Griseb. |
НЧ |
СП |
Гемикриптофит |
Мезофит |
Гликофит |
III.2 |
1 |
||
|
45. Rubiaceae Juss. |
|||||||||
|
269. |
Asperula balchanica Bobrov |
НЧ |
эндемик Б. |
СП |
Гемикриптофит |
Мезолитофит |
Гликофит |
IV.31 |
3 |
|
270. |
A. setosa Jaub. et Spach |
Р |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.2 |
1 |
|
|
271. |
A. trichodes J. Gay ex DC. |
НЧ |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.2 |
1 |
|
|
272. |
Callipeltis cucullaria (L.) Steven |
НЧ |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.1 |
3 |
|
|
273. |
Galium aparine L. |
НР |
ОМ |
Терофит |
Мезофит |
Гликофит |
II.2 |
3 |
|
|
274. |
G. setaceum Lam. |
Р |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.2 |
3 |
|
|
275. |
G. spurium L. |
НР |
ОМ |
Терофит |
Мезофит |
Гликофит |
II.1 |
1,3 |
|
|
276. |
Plocama bruguierei (A. Rich. ex DC.) M. Backlund et Thulin |
Р |
ПкчН |
Хамефит |
Ксерофит |
Гипсофит |
IV.23 |
2 |
|
|
277. |
Rubia florida Boiss. |
НР |
ПкчН |
Хамефит |
Мезоксерофит |
Гликофит |
IV.8 |
2-3 |
|
|
46. Boraginaceae Juss. |
|||||||||
|
278. |
Argusia sogdiana (Bunge) Czerep. |
НР |
ДТП |
Геофит |
Мезоксерофит |
Гликофит |
IV.14 |
4 |
|
|
279. |
Arnebia decumbens (Vent.) Coss. et Kralik |
НР |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.1 |
1-4 |
|
|
280. |
A. minima Wettst. ( A. linearifolia DC.) |
Р |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.2 |
4 |
|
|
281. |
A. transcaspica Popov |
НЧ |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.2 |
4 |
|
|
282. |
Asperugo procumbens L. |
НЧ |
ОМ |
Терофит |
Мезофит |
Гликофит |
II.2 |
1-5 |
|
|
283. |
Buglossoides tenuiflora (L. f.) I. M. Johnst. |
НР |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.1 |
1-4 |
|
|
284. |
Caccinia macranthera (Banks et Sol.) Brand |
НР |
СП |
Гемикриптофит |
Мезофит |
Гипсофит |
IV.4 |
2-3 |
|
|
285. |
Heliotropium arguzioides Kar. et Kir. |
НР |
ДТП |
Геофит |
Мезоксерофит |
Гликофит |
IV.14 |
4 |
|
|
№ |
Семейства и виды |
н о о « рц |
Красная книга и эндемизм |
Жизненные формы по… |
Ареал |
н Д' |
|||
|
Серебрякову |
Раункиеру |
Вармингу |
отношению к засолению |
||||||
|
286. |
H. dasycarpum Ledeb. |
НР |
СП |
Гемикриптофит |
Мезоксерофит |
Гликофит |
III.2 |
4 |
|
|
287. |
H. ellipticum Ledeb. |
Ч |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.1 |
1-4 |
|
|
288. |
H. litvinovii Popov |
Р |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.28 |
2 |
|
|
289. |
Lappula barbata (M. Bieb.) Gurke |
НЧ |
Дв |
Гемикриптофит |
Мезофит |
Гликофит |
III.2 |
1-3 |
|
|
290. |
L. semiglabra (Ledeb.) Gurke |
НР |
ОМ |
Терофит |
Мезоксерофит |
Гликофит |
IV.14 |
4 |
|
|
291. |
L. sessilifolia (Boiss.) Gurke |
НЧ |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.23 |
1-3 |
|
|
292. |
L. sinaica (DC.) Asch. ex Schweinf. |
НР |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.2 |
1-3 |
|
|
293. |
L. spinocarpos (Forssk.) Asch. ex Kuntze |
Ч |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.1 |
1-3 |
|
|
294. |
L. spinocarpos subsp. ceratophora (Popov) Y.J. Nasir |
НР |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.2 |
1-3 |
|
|
295. |
Microparacaryum intermedium (Fresen.) Hilger et Podlech |
НЧ |
ОМ |
Терофит |
Мезофит |
Гипсофит |
III.2 |
2 |
|
|
296. |
M. refracta Boiss. |
НР |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.1 |
1 |
|
|
297. |
Nonea caspica (Willd.) G. Don |
Ч |
ОМ |
Терофит |
Мезофит |
Галогликофит |
IV.2 |
4 |
|
|
298. |
Onosma leucocarpa Popov |
НЧ |
эндемик КХ |
СП |
Гемикриптофит |
Мезоксерофит |
Гипсофит |
IV.28 |
2 |
|
299. |
O. staminea Ledeb. |
Р |
СП |
Гемикриптофит |
Мезоксерофит |
Гипсофит |
IV.14 |
2 |
|
|
300. |
Paracaryum turcomanicum Bornm. et Sint. |
НР |
СП |
Гемикриптофит |
Мезоксерофит |
Гипсофит |
IV.30 |
2 |
|
|
301. |
Rochelia disperma (L.fil.) K. Koch |
НР |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.1 |
1 |
|
|
302. |
R. retorta (Pall.) Lipsky |
НЧ |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.2 |
1-3 |
|
|
303. |
Suchtelenia calycina (C.A. Mey.) A. DC. |
Р |
ОМ |
Терофит |
Мезофит |
Гипсофит |
III.2 |
2 |
|
|
47. Convolvulaceae Juss. |
|||||||||
|
304. |
Convolvulus erinaceus Ledeb. |
Р |
ПкчН |
Хамефит |
Ксерофит |
Гликофит |
IV.2 |
4 |
|
|
305. |
C. fruticosus Pall. |
Ч |
КчН |
Фанерофит |
Ксерофит |
Гипсофит |
IV.2 |
2,4 |
|
|
306. |
C. korolkowii Regel et Schmalh. |
НЧ |
СП |
Гемикриптофит |
Мезоксерофит |
Гликофит |
IV.18 |
4 |
|
|
307. |
Cuscuta brevistyla A. Braun ex A. Rich. |
НЧ |
Опар |
Терофит |
Мезофит |
Гликофит |
III.1 |
1-5 |
|
|
48. Solanaceae Juss. |
|||||||||
|
308. |
Lycium depressum Stocks |
НЧ |
КН |
Фанерофит |
Мезофит |
Галогликофит |
III.2 |
2,4 |
|
|
309. |
L. ruthenicum Murray |
НР |
КН |
Фанерофит |
Мезоксерофит |
Галогликофит |
III.4 |
4 |
|
|
49. Lamiaceae Lindl. |
|||||||||
|
310. |
Acinos rotundifolius Pers. |
Р |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.2 |
1 |
|
|
311. |
Chamaesphacos ilicifolius Schrenk |
Р |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.2 |
4 |
|
|
312. |
Lallemantia royleana ( Benth.) Benth. |
ОЧ |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.2 |
1-4 |
|
|
313. |
Marrubium anisodon K. Koch |
Р |
СП |
Гемикриптофит |
Мезофит |
Гликофит |
IV.8 |
3 |
|
|
314. |
M. vulgare L . |
НЧ |
СП |
Гемикриптофит |
Мезофит |
Гликофит |
III.7 |
3 |
|
|
315. |
Mentha longifolia (L.) Huds. |
Р |
КотП |
Гемикриптофит |
Мезофит |
Гликофит |
IV.8 |
5 |
|
|
316. |
Nepeta bituminosa (Fisch. et C.A. Mey.) Jamzad et |
Ч |
ПкчН |
Хамефит |
Мезофит |
Гликофит |
IV.11 |
1,3 |
|
|
№ |
Семейства и виды |
из н о о « ^ |
Красная книга и эндемизм |
Жизненные формы по… |
Ареал |
н Д' |
|||
|
Серебрякову |
Раункиеру |
Вармингу |
отношению к засолению |
||||||
|
Serpoosh. |
|||||||||
|
317. |
N. cataria L . |
НЧ |
СП |
Гемикриптофит |
Мезофит |
Гликофит |
III.4 |
3 |
|
|
318. |
Salvia abrotanoides (Kar.) Sytsma |
Ч |
ПкчН |
Хамефит |
Мезоксерофит |
Гликофит |
IV.10 |
3 |
|
|
319. |
Stachys turcomanica Trautv. |
НР |
ПкчН |
Хамефит |
Мезофит |
Гликофит |
IV.28 |
2-3 |
|
|
320. |
Teucrium polium L. |
НР |
СП |
Гемикриптофит |
Мезофит |
Гликофит |
III.1 |
2-3 |
|
|
321. |
Thuspeinantha persica (Boiss.) Briq. |
Р |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.2 |
4 |
|
|
322. |
Ziziphora tenuior L. |
Ч |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.4 |
1-4 |
|
|
50. Mazaceae Reveal |
|||||||||
|
323. |
Dodartia orientalis L. |
Р |
СП |
Гемикриптофит |
Мезофит |
Гликофит |
III.4 |
3 |
|
|
51. Orobanchaceae Vent. |
|||||||||
|
324. |
Cistanche fissa (C.A. Mey.) G.Beck |
Р |
ПарП |
Гемикриптофит |
Ксерофит |
Гликофит |
IV.7 |
4 |
|
|
325. |
C. mongolica G.Beck |
НР |
ПарП |
Гемикриптофит |
Ксерофит |
Гликофит |
III.1 |
2,4-5 |
|
|
326. |
C. salsa (C.A. Mey.) G. Beck |
Р |
ПарП |
Гемикриптофит |
Ксерофит |
Гликофит |
IV.2 |
4 |
|
|
327. |
Orobanche aegyptiaca Pers. |
НР |
ПарП |
Гемикриптофит |
Ксерофит |
Гликофит |
III.1 |
1-4 |
|
|
52. Plantaginaceae Juss. |
|||||||||
|
328. |
Linaria simplex (Willd.) DC. |
Р |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.5 |
1-2 |
|
|
329. |
Plantago lanceolata L. |
Р |
КисП |
Гемикриптофит |
Мезофит |
Гликофит |
II.2 |
1 |
|
|
330. |
Veronica campylopoda Boiss. |
Ч |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.2 |
1-4 |
|
|
331. |
Scrophularia scoparia Pennell |
НЧ |
СП |
Гемикриптофит |
Мезоксерофит |
Гипсофит |
IV.27 |
2 |
|
|
54. Asteraceae Dumort. |
|||||||||
|
332. |
Acantholepis orientalis Less. |
НР |
ОМ |
Терофит |
Ксерофит |
Гликофит |
III.2 |
2,4 |
|
|
333. |
Achillea arabica Kotschy |
ОЧ |
КТП |
Гемикриптофит |
Мезофит |
Гликофит |
III.4 |
1 |
|
|
334. |
A. wilhelmsii K. Koch |
Ч |
КТП |
Гемикриптофит |
Мезофит |
Гликофит |
IV.8 |
1-2 |
|
|
335. |
Amberboa amberboi (L.) Tzvelev |
НР |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.20 |
1 |
|
|
336. |
A. turanica Iljin |
НР |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.2 |
4 |
|
|
337. |
Artemisia badhysi Krasch. et Lincz. ex Poljakov |
ОЧ |
ПкчН |
Хамефит |
Ксерофит |
Гликофит |
IV.28 |
1-3 |
|
|
338. |
A. dumosa Poljak. |
НЧ |
ПкчН |
Хамефит |
Ксерофит |
Гликофит |
IV.18 |
2 |
|
|
339. |
A. santolina Schrenk |
НЧ |
СП |
Гемикриптофит |
Ксерофит |
Гликофит |
IV.2 |
4 |
|
|
340. |
A. scoparia Waldst. et Kit. |
НР |
Дв |
Гемикриптофит |
Ксерофит |
Гликофит |
III.7 |
1-4 |
|
|
341. |
Carduus arabicus subsp. cinereus (M. Bieb.) Kazmi |
НЧ |
ОМ |
Терофит |
Мезоксерофит |
Гликофит |
IV.8 |
3 |
|
|
342. |
Carthamus oxyacanthus M. Bieb . |
НЧ |
ОМ |
Терофит |
Ксерофит |
Гликофит |
III.2 |
3-4 |
|
|
343. |
Centaurea solstitialis L. |
НР |
ОМ |
Терофит |
Мезоксерофит |
Гликофит |
I.1 |
3 |
|
|
344. |
C. virgata subsp. squarrosa (Boiss.) Gugler |
Ч |
СП |
Гемикриптофит |
Мезоксерофит |
Гликофит |
III.3 |
1-3 |
|
|
№ |
Семейства и виды |
н о о « рц |
Красная книга и эндемизм |
Жизненные формы по… |
Ареал |
н Д' |
|||
|
Серебрякову |
Раункиеру |
Вармингу |
отношению к засолению |
||||||
|
345. |
Cichorium ihtybus L. |
НЧ |
СП |
Гемикриптофит |
Мезофит |
Гликофит |
III.7 |
5 |
|
|
346. |
Cnicus benedictus L. |
Р |
ОМ |
Терофит |
Мезоксерофит |
Гликофит |
III.7 |
1 |
|
|
347. |
Cousinia balchanica Tscherneva |
ОР |
эндемик Б. |
Дв |
Гемикриптофит |
Ксерофит |
Гликофит |
IV.31 |
3 |
|
348. |
C. erectispina Tscherneva |
Р |
ОМ |
Терофит |
Ксерофит |
Гликофит |
IV.18 |
4 |
|
|
349. |
C. glandulosa Kult. |
НР |
СП |
Гемикриптофит |
Ксерофит |
Гипсофит |
IV.29 |
2 |
|
|
350. |
C. onopordioides Ledeb. |
НЧ |
СП |
Гемикриптофит |
Ксерофит |
Гипсофит |
IV.2 |
2-3 |
|
|
351. |
C. oxiana Tscherneva |
Ч |
Дв |
Гемикриптофит |
Ксерофит |
Гликофит |
IV.4 |
4 |
|
|
352. |
C. schistoptera Juz. |
НЧ |
Дв |
Гемикриптофит |
Ксерофит |
Гликофит |
IV.18 |
4 |
|
|
353. |
C. turcomanica C.Winkl. |
НР |
ПкчН |
Хамефит |
Ксерофит |
Гипсофит |
IV.28 |
1-2 |
|
|
354. |
C. turkmenorum Bornm. |
НР |
Дв |
Гемикриптофит |
Ксерофит |
Гликофит |
IV.28 |
1 |
|
|
355. |
Crepis pulchra L. |
НР |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.2 |
1,3 |
|
|
356. |
Dipterocome pusilla Fisch. et C.A. Mey. |
НЧ |
ОМ |
Терофит |
Мезоксерофит |
Галогликофит |
IV.2 |
4 |
|
|
357. |
Echinops ritro L. |
Р |
СП |
Гемикриптофит |
Ксерофит |
Гликофит |
II.1 |
1-4 |
|
|
358. |
E. hemilasia (Bunge) Clarke |
ОЧ |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.6 |
1-4 |
|
|
359. |
Filago griffithii (A. Gray) Andres-Sanchez et Galbany |
Ч |
ОМ |
Терофит |
Мезоксерофит |
Гликофит |
III.2 |
1 |
|
|
360. |
Filago hurdwarica (DC.) Wagenitz |
НР |
ОМ |
Терофит |
Мезоксерофит |
Гликофит |
III.1 |
1-2 |
|
|
361. |
Garhadiolus papposus Boiss. et Buhse |
НЧ |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.2 |
1-4 |
|
|
362. |
Hyalea pulchella (Ledeb.) K. Koch |
Ч |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.2 |
4 |
|
|
363. |
Jurinea antonowii C. Winkl. |
НР |
ПкчН |
Хамефит |
Ксерофит |
Гипсофит |
IV.28 |
2 |
|
|
364. |
J. lydiae Iljin |
Р |
эндемик Б. |
СП |
Гемикриптофит |
Ксерофит |
Гипсофит |
IV.31 |
2 |
|
365. |
Karelinia caspia (Pall.) Less. |
НР |
КотП |
Гемикриптофит |
Гигрофит |
Галогликофит |
IV.2 |
5 |
|
|
366. |
Koelpinia linearis Pall. |
Ч |
ОМ |
Терофит |
Мезоксерофит |
Гликофит |
III.2 |
1-4 |
|
|
367. |
Lactuca orientalis (Boiss.) Boiss. |
НР |
ПкчН |
Хамефит |
Мезофит |
Гликофит |
III.1 |
5 |
|
|
368. |
L. tatarica (L.) C.A. Mey. |
Р |
КотП |
Гемикриптофит |
Мезофит |
Гликофит |
II.2 |
5 |
|
|
369. |
L. undulata Ledeb. |
НЧ |
ОМ |
Терофит |
Мезофит |
Гипсофит |
III.2 |
2 |
|
|
370. |
Lagoseris aralensis (Bunge) Boiss. |
НЧ |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.14 |
4 |
|
|
371. |
L. sancta (L.) K. Maly |
НР |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.5 |
4 |
|
|
372. |
Microcephala lamellata (Bunge) Pobed. |
Ч |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.14 |
1-4 |
|
|
373. |
Pseudopodospermum ovatum (Trautv.) Zaika, Sukhor. et N. Kilian |
НЧ |
КлбП |
Геофит |
Мезоксерофит |
Гипсофит |
IV.1 |
2 |
|
|
374. |
P. raddeanum (C. Winkl.) Zaika, Sukhor. et N. Kilian |
НЧ |
КлбП |
Геофит |
Мезоксерофит |
Гликофит |
IV.28 |
1-2 |
|
|
375. |
Rhabdotheca korovinii (Popov) Kirp. |
Р |
ОМ |
Терофит |
Ксерофит |
Гликофит |
IV.14 |
4 |
|
|
376. |
Rhaponticum repens (L.) Hidalgo |
НР |
КотП |
Гемикриптофит |
Мезофит |
Гликофит |
III.4 |
1 |
|
|
377. |
Senecio subdentatus Ledeb. |
НР |
ОМ |
Терофит |
Мезофит |
Гликофит |
IV.2 |
4 |
|
|
№ |
Семейства и виды |
н о о « рц |
Красная книга и эндемизм |
Жизненные формы по… |
Ареал |
н Д' |
|||
|
Серебрякову |
Раункиеру |
Вармингу |
отношению к засолению |
||||||
|
378. |
S. vernalis Waldst. et Kit. |
Ч |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.3 |
1-3 |
|
|
379. |
Takhtajaniantha pusilla (Pall.) Nazarova |
НР |
КлбП |
Геофит |
Мезоксерофит |
Гипсофит |
IV.2 |
2 |
|
|
380. |
Xanthium strumarium L. |
НЧ |
ОМ |
Терофит |
Мезофит |
Гликофит |
I.1 |
3 |
|
|
55. Caprifoliaceae Juss. |
|||||||||
|
381. |
Lomelosia olivieri (Coult.) Greuter et Burdet |
НЧ |
ОМ |
Терофит |
Мезоксерофит |
Гликофит |
III.7 |
1,4 |
|
|
382. |
L. rhodantha (Kar. et Kir.) Sojak |
НЧ |
ОМ |
Терофит |
Мезоксерофит |
Гликофит |
III.8 |
4 |
|
|
383. |
Valerianella dufresnia Bunge |
НР |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.1 |
4 |
|
|
384. |
V. leiocarpa (K. Koch) Kuntze |
Р |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.1 |
1 |
|
|
56. Apiaceae Lindl. |
|||||||||
|
385. |
Apium graveolens L. |
Р |
Дв |
Гемикриптофит |
Мезофит |
Гликофит |
III.7 |
5 |
|
|
386. |
Bupleurum exaltatum M. Bieb. |
НЧ |
СП |
Гемикриптофит |
Мезофит |
Гликофит |
III.2 |
3 |
|
|
387. |
Cuminum setifolium (Boiss.) Koso-Pol. |
Ч |
ОМ |
Терофит |
Мезоксерофит |
Гликофит |
IV.2 |
1-4 |
|
|
388. |
Elwendia longipes (Freyn) Pimenov et Kljuykov |
Ч |
КлбП |
Геофит |
Мезофит |
Гликофит |
IV.30 |
1-3 |
|
|
389. |
Eremodaucus lehmannii Bunge |
НР |
ОМ |
Терофит |
Мезоксерофит |
Гипсофит |
IV.8 |
4 |
|
|
390. |
Eryngium bungei Boiss. |
НР |
СП |
Гемикриптофит |
Ксерофит |
Гликофит |
IV.8 |
1-4 |
|
|
391. |
Ferula karakalensis Korovin |
НЧ |
ММ |
Гемикриптофит |
Мезофит |
Гликофит |
IV.29 |
2 |
|
|
392. |
F. karelinii Bunge |
НР |
ММ |
Гемикриптофит |
Мезоксерофит |
Гипсофит |
IV.2 |
2,4 |
|
|
393. |
F. litwinowiana Koso-Pol. |
Р |
ММ |
Гемикриптофит |
Мезоксерофит |
Гликофит |
IV.18 |
4 |
|
|
394. |
F. oopoda (Boiss. et Buhse) Boiss. |
НЧ |
СП |
Гемикриптофит |
Мезофит |
Гликофит |
IV.2 |
3 |
|
|
395. |
F. ovina (Boiss.) Boiss. |
Р |
СП |
Гемикриптофит |
Мезофит |
Гликофит |
IV.23 |
1-3 |
|
|
396. |
F. szowitsiana DC. |
НЧ |
ММ |
Гемикриптофит |
Мезофит |
Гипсофит |
IV.2 |
1-3 |
|
|
397. |
Prangos latiloba Korovin |
НЧ |
СП |
Гемикриптофит |
Мезоксерофит |
Гипсофит |
IV.28 |
2 |
|
|
398. |
Scandix stellata Banks et Sol. |
Ч |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.1 |
1-3 |
|
|
399. |
Seseli jomuticum Schischk. |
Р |
СП |
Гемикриптофит |
Мезоксерофит |
Гипсофит |
IV.2 |
2 |
|
|
400. |
Turgenia latifolia (L.) Hoffm. |
Р |
ОМ |
Терофит |
Мезофит |
Гликофит |
III.1 |
1-3 |
|
|
401. |
Zosima absinthifolia (Vent.) Link |
НР |
СП |
Гемикриптофит |
Мезофит |
Гликофит |
III.2 |
1-3 |
|
Пояснение к таблице . Сокращения в столбце «Редкость» по категориям встречаемости: ОЧ – очень часто; Ч – часто; НЧ – нечасто; НР – нередко; Р – редко; ОР – очень редко. В столбце «Красная книга и эндемизм» отмечены виды, занесённые в 4-ое издание Красной книги Туркменистана, отмечены их категории: I (CR) – вид на грани исчезновения; II (EN) – исчезающий вид; III (VU) – уязвимый вид; IV – редкий вид. За эндемичные виды мы принимаем только узколокальные балханские (Б) и копетдаг-хорасанские элементы (К-Х). Сокращения в столбце «Жизненные формы по Серебрякову»: дерево (ВД – вечнозеленое; ЛД – листопадное); кустарник (КН – несуккулентный; КС – суккулентный; БВК – безлистный вечнозеленый); кустарничек (КчН – несуккулентный; ПКч – подушковидный; ПарК – паразитический); полукустарнички (ПкчН – несуккулентный; ПкчС – суккулентный; ППкч -подушковидный); ПкН – полукустарник несуккулентный; травянистые поликарпики (СП – стержнекорневой; КисП – кистекорневой; КТП – короткокорневищный; ДТП – длиннокорневищный; ДрТП – дерновинообразующий; КлбП – клубнеобразующий и клубнекорневой; ЛП, КлП – луковичный и клубнелуковичный; КкП – клубнекорневищный; КлкП – клубнелуковично-корневищный; КотП – корнеотпрысковый; ПарП – паразитический); травянистые монокарпики (ОМ – однолетний; Дв – двулетний; ОС – однолетний суккулентный; Опар – однолетний паразитический; ММ – многолетний монокарпик). Сокращения в столбце «Ареалы» расписаны в таблице 4 статьи.
Основные угрозы
-
1. Неконтролируемый выпас скота проводится повсеместно как в предгорьях, так и на вершине хребта исключительно в зимне-весенний период, когда нет нужды регулярного водопоя. Единичные источники воды минерализованы и фактически для потребленя скотом не годятся. Большие территории Малого Балхана заняты бедлендами, которые не имеют пастбищной ценности. Хотя негативное влияние чрезмерной пастбищной нагрузки на более богатые травами участки ощущается отчетливо.
-
2. Природные пожары в прошлом уничтожили обширные арчовые редколесья, от которых осталось лишь два экземпляра. Большой урон арчовникам нанесла вырубка деревьев в начале прошлого века на нужды железнодорожников. Арчу, как и саксаул использовали в качестве топлива для паровозов. Возобновления арчовников в настоящее время не отмечено.
-
3. Добыча строительного камня проводится на северных склонах главного хребта в незначительных количествах, однако приводит к нарушению фитоценозов.
Заключение
Флора Малого Балхана начала формирование относительно позже флоры Большого Балхана, т.к. система последнего значительно выше Малого Балхана и появление суши произошло на Большом Балхане раньше. Своеобразные рельефы этих хребтов, сильно отличающиеся друг от друга, и несколько различные климатические условия, особенно верхних участков гор, в целом повлияли на их флористический состав. Анализируя сопредельные Малому Балхану флоры, можно сделать заключение, что флора данного региона более близка флоре Каракумов. Отмечается миграция туранских видов на горные территории, и, напротив, миграции реликтовых малобалханских видов в Каракумы мы не выявили. Наиболее древними реликтовыми растениями являются виды древнесредиземноморского генезиса, которые постепенно в процессе аридизации климата и ксерофитизации растительности вытесняются наиболее приспособленными ксерофитами.
Как уже было сказано выше, планируемая закладка Балханского государственного природного заповедника с Малобалханским заказником в своем составе непременно будет способствовать сохранению экосистемы. Планируются и посадки арчи, что необходимо для восстановления первичной целостной экосистемы с доминированием можжевеловых редколесий.
Авторы выражают глубокую признательность Фонду Михаэля Зуккова (Грайфсвальд, Германия) и Фонду Марион Дёнхофф за финансовую помощь, позволившую провести ряд экспедиций и обработку материалов статьи.
