Современные биологические характеристики сига Coregonus lavaretus, европейской ряпушки С. albula и европейской корюшки Osmerus eperlanus озера Имандра

Автор: Зубова Е.М., Кашулин Н.А., Терентьев П.М.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Зоология
Статья в выпуске: 3, 2020 года.
Бесплатный доступ
Исследованы современные биологические характеристики многочисленных видов рыб плёсов оз. Имандра - сига, европейской ряпушки и европейской корюшки. Показано, что в условиях продолжающегося негативного изменения гидрологического и гидрохимического режимов озера за последние 20 лет, произошли разнонаправленные изменения в структурах популяций этих видов. Если у короткоцикловых видов (корюшка, ряпушка) наблюдается положительная динамика ряда показателей, то у сига эти изменения носят негативный характер. Эврифагия корюшки оз. Имандра, включая переход на хищничество при достижении длины 150 мм, в сочетании с эффективной стратегией размножения и практическим отсутствием пресса хищников, обеспечивают ей высокую численность и успешную конкуренцию с другими видами.
Сиг, европейская ряпушка, европейская корюшка, биологические характеристики, оз. имандра
Короткий адрес: https://sciup.org/147229656
IDR: 147229656 | УДК: 597.553.2.574.3 | DOI: 10.17072/1994-9952-2020-3-210-226
Modern biological characteristics of whitefish Coregonus lavaretus, european vendace C. albula and European smelt Osmerus eperlanus from the Imandra lake
The modern biological characteristics of numerous fish species from the Imandra Lake reaches - white-fish, european vendace and european smelt was investigated. It was shown that under the conditions of the ongoing negative change in the hydrological and hydrochemical regimes of the lake over the past 20 years, there have been multidirectional changes in the population's structures of these species. If in short-cycle species (smelt, vendace) there is a positive dynamics of a number of indicators, then in whitefish these changes are negative. Euryphagia of smelt from the Imandra Lake, including the transition to preda-tion when reaching a length of 150 mm, combined with an effective breeding strategy and the virtual absence of a predator press, provide him high numbers and successful competition with other species.
Текст научной статьи Современные биологические характеристики сига Coregonus lavaretus, европейской ряпушки С. albula и европейской корюшки Osmerus eperlanus озера Имандра
Антропогенные сукцессии крупных арктических водоемов мало изучены в силу специфики условий, многолетней динамики глобальных и региональных факторов окружающей среды, относительной продолжительностью процесса и, как следствие, отсутствием полноценных рядов наблюдений, выполненных по единым методикам и позволяющие накапливать единообразные и сравнимые данные по всем компонентам экосистемы. Существуют генерализированные пред- ставления о направленности тех или иных изменений как ответ на различные виды воздействий, но они часто модифицируются совокупностью факторов различной природы и не могут служить надёжной основой для прогноза. Попытки решить возникающие методологические проблемы с помощью математического моделирования [Решетников, Терещенко, Лукин, 2011], к сожалению, не могут учесть все биотические и абиотические компоненты даже самых простых природных экосистем, а тем более, крупных водоемов. Все это требует проведения новых исследований, расширяю-
щих наши представления об особенностях функционирования экосистем в современных условиях.
Озеро Имандра – один из крупнейших водоемов арктической зоны РФ, испытывающий на протяжении многих десятков лет мощное многофакторное воздействие, включающее изменения гидрологического и гидрохимического режимов, структурно-функциональной организации биотической составляющей [Чижиков, 1980; Моисеенко, Яковлев, 1990; Антропогенные …, 2002; Кашулин и др., 2013; Даувальтер, Кашулин, 2015а, б; Dau-valter, Kashulin 2018]. Последние десятилетия среди модифицирующих факторов на первое место выходит эвтрофирование водоема [Денисов, Ка-шулин, 2016; Терентьева, Кашулин, Денисов, 2017]. Значительные изменения затронули все структурные компоненты биоты водоёма, начиная от первичных продуцентов, заканчивая рыбной частью сообщества, что привело к снижению его рыбопромыслового значения [Решетников, Терещенко, Лукин, 2011; Вандыш, 2012; Валькова и др., 2012; Кашулин и др., 2013; Даувальтер, Терентьев, 2018]. Изменилась и структура ихтиофауны: практически исчезли лососёвые виды, сократилась доля сигов, в то время как численность европейской корюшки резко увеличилась, и она расселяется по придаточным озерно-речным системам [Терентьев, Кашулин, Зубова, 2017]. Корюшка, благодаря короткому циклу развития, пищевой пластичности, отсутствию пресса хищников и выгодной стратегии размножения (весенний нерест в притоках), становится доминирующим видом. Рыбы, занимающие верхние трофические уровни арктических водоемов, интегрируют все изменения, происходящие в экосистеме, и являются надежными показателями долговременных трендов [Кашулин, 2004].
Цель работы – анализ современного состояния популяций наиболее многочисленных видов рыб оз. Имандра – сига, европейской ряпушки и европейской корюшки.
Материал и методы исследования
Изучение популяционной структуры сига, ряпушки и корюшки оз. Имандра проводилось в рамках комплексных исследований озера в 2011– 2016 гг. Районы исследований – три плёса озера (Большая, Йокостровская и Бабинская Имандры), расположенные по градиенту нагрузки от источников загрязнения [Антропогенные…, 2002; Зубова и др., 2018]. Северный плёс Большая Имандра (далее БоИ) – наиболее загрязняемая часть озера. Сюда поступают стоки комбинатов горнометаллургического комплекса «Олкон», «Северо-никель», «Апатит» и хозяйственно-бытовые стоки г. Мончегорска, Кировска и Апатит (рисунок). Южный плёс Бабинская Имандра (далее БаИ) – удаленный от источников загрязнения район озера, однако губа Молочная испытывает влияние подогретых сбросных вод Кольской АЭС. В плёсе Йо-костровская Имандра (далее ЙИ) смешиваются воды плёсов Большая и Бабинская Имандра, и происходит сток из озера через р. Ниву [Антропогенные…, 2002].
Отбор гидрохимического и гидробиологического материалов в исследуемых водоёмах производился в период гидробиологического лета (июль-август) и соответствовал местам сбора ихтиологических проб (рис. 1). Определение гидрохимических параметров проводилось в стационарных условиях в лаборатории Центра коллективного пользования Института проблем промышленной экологии Севера Кольского научного центраРАН (ИППЭС КНЦ РАН). Отбор, камеральная обработка и анализ проб фито-, зоопланктона и макрозообентоса проводились согласно общепринятым стандартным методам, описанным ранее [Руководство …, 1992; Денисов, 2011]. Во всех исследуемых р-нах оз. Имандра рыбу отлавливали стандартными наборами ставных жаберных сетей из нейлонового монофиламента. В литоральной зоне (на глубине 1.5–3 м) устанавливались сети длиной 25 м, высотой 1.5 м и размером ячеи 10–60 мм (что обеспечивало вылов рыбы длиной ≥ 5 см). Сети устанавливались порядками по 1–2 сети перпендикулярно берегу, в местах с песчаногравийными отмелями и крупными валунными отложениями. В профундальной зоне с глубинами более 18 м использовалось до 10 разноячеистых сетей в один порядок. В пелагической зоне водоема для отбора ихтиологического материала применялись плавные мультиразмерные сети высотой 3 м. За период с 2011 по 2016 гг. изучено 2 637 экземпляров трёх видов рыб. Подробная информация об объёме материала, местах и времени лова рыб представлена в табл. 1. Обработка материала проводилась по методике Г.П. Сидорова и Ю.С. Решетникова [2014]. Массу рыб определяли с точностью до 1 г, длину по Смиту (АС) измеряли с точностью до 1 мм. Для выделения внутривидовых форм у исследуемых сигов подсчитывали тычинки на первой жаберной дуге. Возраст исследуемых видов рыб определялся по чешуе первым автором по известным методикам [Van Oosten, 1929; McKenzie, 1957; Решетников, 1966; Заварзина, 2005]. Рыбы были отнесены нами к принимающим участие в нересте, если их гонады достигли стадии половой зрелости: у сига и ряпушки – III–IV [Решетников, Богданов, 2011], у корюшки – II-III (из осенних уловов) и III–IV.
теристики спектра питания использовали такие по-
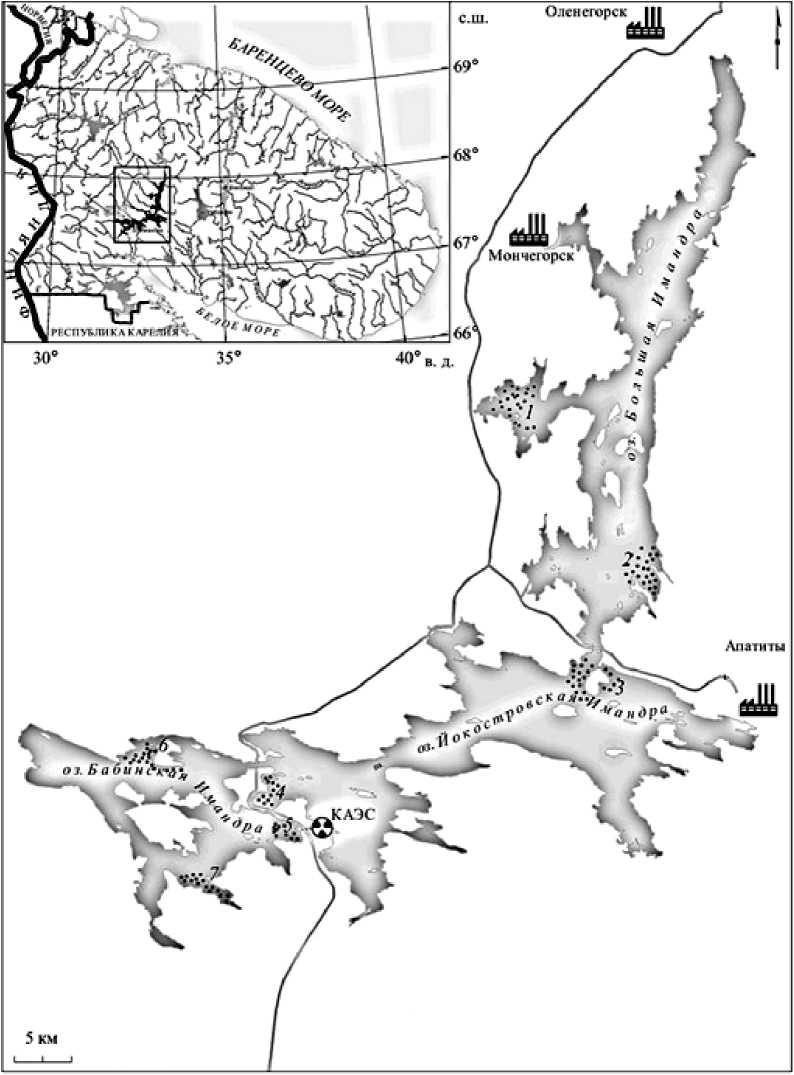
Карта-схема оз. Имандра и места сбора выборок (•) в 2011-2016 гг.:
1 – губа Вите, 2 – губа Белая, район о. Могильный, 3 – район о. Большой Йокостровский, 4 – прол. Узкая Салма, губа Глубокая, 5 – губа Молочная, 6 – район о. Хорт, 7 – губа Кунчаст
Таблица 1
Характеристика использованного материала
|
оз. Имандра, плëс |
Район исследования |
Период исследований |
n 1 |
n 2 |
n 3 |
|
Большая Имандра |
Губа Вите Губа Белая, остров Могильный |
Октябрь 2013 Г. Август-сентябрь 2012 Г., Октябрь 2013 Г. |
Сиг 45 Ряпушка 93 Корюшка 6 Сиг 46 Ряпушка 256 Корюшка 186 |
44 50 |
Сиг 14 Ряпушка 22 Корюшка 0 |
|
Йокостровская Имандра |
Остров Большой Йокостровский Пролив Узкая Сал-ма, губа Глубокая |
Август 2011 г. Ежемесячно с июля 2012 г. по май 2013 гг., Июль-сентябрь 2013 г., Октябрь 2014 г., Ноябрь – декабрь 2015 г., Август 2016 г. Август-сентябрь 2011 г., Июнь 2012 Г., Ноябрь 2015 г |
Сиг 496 Ряпушка 322 Корюшка 463 Сиг 72 Ряпушка 53 Корюшка 173 |
411 24 |
Сиг 76 Ряпушка 42 Корюшка 26 |
|
Бабинская Имандар |
Губа Молочная Остров Хорт Губа Кунчаст |
Октябрь 2011 г. Август-сентябрь 2011 г. Сентябрь 2011 г. |
Сиг 50 Ряпушка 102 Корюшка 14 Сиг 59 Ряпушка 1 Корюшка 28 Сиг 84 Ряпушка 5 Корюшка 83 |
50 59 84 |
Сиг 112 Ряпушка 2 Корюшка 10 |
Окончание табл. 2
|
Показатели |
Плëс |
||
|
БоИ |
ЙИ |
БаИ |
|
|
Биомасса зоопланктона, г/м3 |
4.3 |
2.1 |
0.6 |
|
Трофический статус* |
а - эвтрофный |
в - мезотрофный |
а - олиготрофный |
|
Численность зообентоса, экз/м2 |
4211 |
588 |
398 |
|
Биомасса зообентоса, г/м3 |
20.1 |
3.9 |
1.7 |
|
Трофический статус* |
в - эвтрофный |
а - мезотрофный |
в - олиготрофный |
Примечание.* - цит. по: Китаев, 2007.
|
в структуре рыбной части сообщества. В настоя- Видовой состав уловов. Уловы из трех плёсов щее время ее доля в составе уловов озера может оз. Имандра в 2011‒2016 гг. включали десять ви- варьировать от 30 до 36% (табл. 3). Относительно дов рыб (табл. 3). Наиболее многочисленными в высокая доля сига при этом (до 30%) сохраняется них были сиг, ряпушка, корюшка и окунь (от 12 до лишь в акватории БаИ. Критически снижена чис-48%). Одним из важных аспектов функционирова- ленность арктического гольца. ния современной экосистемы оз. Имандра является значительное усиление роли европейской корюшки Таблица 3 Долговременные изменения соотношения различных видов рыб в уловах оз. Имандра |
|||||||||||
|
Период исследований |
Виды рыб |
||||||||||
|
сиг |
голец |
кумжа |
хариус |
окунь |
щука |
налим |
форель |
ерш |
корюшка |
ряпушка |
|
|
Большая Имандра |
|||||||||||
|
1960 |
67.0 |
19.0 |
2.6 |
3.0 |
0.9 |
2.6 |
4.0 |
- |
- |
1.7 |
- |
|
1965 ‒ 1968 |
60.7 |
13.7 |
0.5 |
0.7 |
2.1 |
1.7 |
0.5 |
- |
0.9 |
18.2 |
|
|
1990-е |
80.0 |
11.5 |
0.4 |
0.4 |
4.0 |
- |
3.2 |
- |
0.5 |
||
|
2010-е |
12.0 |
0.3 |
- |
2.0 |
3.5 |
0.7 |
- |
- |
24.0 |
36.0 |
21.5 |
|
Йокостровская Имандра |
|||||||||||
|
1972 ‒ 1973 |
50.9 |
4.5 |
0.6 |
0.9 |
37.6 |
3.1 |
2.4 |
- |
- |
- |
- |
|
1990-е |
75.6 |
2.8 |
0.5 |
2.6 |
2.3 |
- |
- |
- |
1.6 |
13.9 |
0.7 |
|
2010-е |
19.0 |
- |
0.3 |
2.0 |
3.0 |
0.3 |
- |
- |
6.0 |
30.0 |
40.0 |
|
Бабинская Имандра |
|||||||||||
|
1972 ‒ 1973 |
42.8 |
42.0 |
3.0 |
7.4 |
1.1 |
0.9 |
2.8 |
- |
- |
- |
- |
|
1978 ‒ 1979 |
37.9 |
50.5 |
- |
- |
- |
6.5 |
- |
5.1 |
- |
- |
- |
|
1990-е |
86.0 |
6.6 |
0.7 |
2.9 |
0.7 |
- |
2.9 |
- |
- |
0.3 |
- |
|
2010-е |
31.0 |
3.0 |
- |
5.0 |
11.0 |
2.0 |
2.0 |
- |
12.0 |
32.0 |
2.0 |
|
Примечание. Доминантные виды составляют > 20% от улова, субдоминантные – 8‒20% – по: Решетников и др., 2011. Здесь и в табл. 4 ‒ 7 «-» – отсутствие в выборке. |
|||||||||||
Результаты
Гидрохимические и гидробиологические показатели трёх плесов оз. Имандра различаются, отражая уровни антропогенной нагрузки (табл. 2). Значительному содержанию биогенных элементов (общего фосфора и азота) в плёсах ЙИ и БоИ соот-
ветствуют высокие показатели первичной продукции, зоопланктона и макрозообентоса. Трофический статус этих плёсов определяется от мезо-трофного до эвтрофного; только воды плёса БаИ близки к природному состоянию и соответствуют олиготрофному трофическому статусу (табл. 2).
Таблица 2
Средние величины гидрохимических и гидробиологических показателей в трех плëсах оз. Имандра, 2011 ‒ 2013 гг.
|
Показатели |
Плëс |
||
|
БоИ |
ЙИ |
БаИ |
|
|
S , км2 |
311.6 |
352.2 |
148.7 |
|
Наибольшая глубина, м |
67.0 |
42.0 |
43.5 |
|
Средняя глубина, м |
14.7 |
10.9 |
16.3 |
|
О 2 , мг/л |
9.37 |
8.98 |
7.39 |
|
рН |
7.78 |
7.42 |
7.33 |
|
Общая минерализация, мг/л |
91.3 |
70.8 |
39.9 |
|
P общ, мкг/л |
55 |
19 |
6 |
|
N общ, мкг/л |
396 |
183 |
147 |
|
Биомасса фитопланктона, г/м3 |
2.98 |
1.57 |
0.46 |
|
Содержание хлорофилла « a », мг/м3 |
6.12 |
3.68 |
1.41 |
|
Трофический статус* |
β - мезотрофный |
α - мезотрофный |
α - олиготрофный |
|
Численность зоопланктона, экз/м3 |
1825.5 |
706.5 |
276.5 |
Внутривидовые формы сига. Сиг в уловах из оз. Имандра представлен двумя формами: малотычинковой и среднетычинковой. У малотычинкового сига число тычинок варьирует от 15 до 30 (в среднем для озера 23.2 ± 0.1); у среднетычинкового – от 32 до 43 (38.3 ± 0.7). Малотычинковый сиг распространен по всему озеру, среднетычинковый – малочислен, и его распределение по озеру крайне неравномерное. Большая часть среднетычинкового сига поймана в плёсе БоИ (15% от уловов сига), в ЙИ его доля составила 1%, в БаИ – 0%. Особи среднетычинкового сига из ЙИ и БоИ при рассмотрении были объединены и описывались как среднетычинковый сиг оз. Имандра.
Возрастной состав. В целом малотычинковый сиг и корюшка в оз. Имандра были представлены десятью возрастными группами: от 0+ до 9+ и от 1+ до 10+ лет соответственно, ряпушка – семью возрастными группами: от 0+ до 6+, среднетычинковый сиг – шестью: от 2+ до 7+ (табл. 4).
Однако возрастная структура уловов рыб в трех плёсах различалась. Особи всех возрастов у исследуемых видов встречались только в уловах из ЙИ. В БоИ и ЙИ преобладали малотычинковые сиги в возрасте 3+, 4+ и 5+ (в среднем 76–82%), в БаИ – в возрасте 4+–6+ (82%) (табл. 4). В ЙИ малотычинковые сиги старше 7+ встречались единично и в основном были представлены самками. В уловах среднетычинкового сига наиболее часто встречались особи двух возрастов – 3+–4+ (75%). У ряпушки в БоИ и ЙИ преобладали рыбы в возрасте 2+–3+ (от 63 до 71%), в то время как в БаИ – меньшего возраста: 1+–2+ (82 %) (табл. 4). Ряпуш- ки максимального возраста (6+) встречались в уловах из плёсов БаИ и ЙИ в единичных экземплярах и были представлены самками. У корюшки в ЙИ и БоИ преобладали рыбы в возрасте от 4+ до
7+ (от 73 до 85% соответственно), в БаИ - в возрасте 4+ и 5+ (80%) (табл. 4). Корюшки старше 8+ лет в ЙИ и БаИ были представлены главным образом самками и встречались единично (1-3%).
Таблица 4
Возрастной и половой состав у малотычинкового (мт.с.) и среднетычинкового (ст.с.) сига
Coregonus lavaretus , европейской ряпушки C. albula и европейской корюшки Osmerus eperlanus в плёсах оз. Имандра, 2011 - 2016 гг.
|
Место лова, плёс |
Воз |
раст, лет |
В целом для |
||||||||||
|
Виды |
0+ |
1+ |
2+ |
3+ |
4+ |
5+ |
6+ |
7+ |
8+ |
9+ |
10+ |
выборки |
|
|
рыб |
juv /самцы/самки, |
n |
juv /самцы/ |
||||||||||
|
(доля возрастной группы от общей численности выборки, %) |
самки, n |
||||||||||||
|
БоИ |
мт.с. |
0/0/1 |
0/1/4 |
0/8/1 |
0/26/13 |
0/10/5 |
0/3/3 |
0/1/1 |
0/49/28 |
||||
|
- |
(1.3) |
(6.5) |
(11.7) |
(50.6) |
(19.5) |
(7.8) |
(2.6) |
- |
- |
- |
|||
|
ряпушка |
2/0/0 |
3/9/12 |
1/58/79 |
1/44/54 |
0/7/46 |
0/2/16 |
7/120/207 |
||||||
|
(0.6) |
(7.2) |
(41.3) |
(29.6) |
(15.9) |
(5.4) |
||||||||
|
корюшка |
1/2/2 |
0/4/13 |
0/6/19 |
0/7/19 |
0/5/33 |
0/1/22 |
0/1/14 |
0/0/4 |
1/26/126 |
||||
|
ЙИ |
(3.3) |
(11.1) |
(16.3) |
(17.0) |
(24.9) |
(15.0) |
(9.8) |
(2.6) |
|||||
|
мт.с. |
0/0/2 |
4/2/12 |
6/18/25 |
0/66/60 |
0/76/108 |
0/37/72 |
0/17/29 |
0/2/12 |
0/0/2 |
0/1/1 |
10/219/323 |
||
|
(0.5) |
(2.9) |
(9.0) |
(22.8) |
(33.3) |
(19.7) |
(8.3) |
(2.5) |
(0.5) |
(0.5) |
- |
|||
|
ряпушка |
21/1/5 |
8/14/23 |
4/48/75 |
0/16/53 |
0/5/27 |
0/0/11 |
0/0/1 |
33/84/195 |
|||||
|
(8.7) |
(14.4) |
(40.7) |
(22.2) |
(10.2) |
(3.5) |
(0.3) |
- |
- |
- |
- |
|||
|
корюшка |
0/0/1 |
3/2/4 |
4/5/21 |
3/14/43 |
0/24/72 |
0/20/107 |
0/2/44 |
0/0/12 |
0/0/3 |
0/0/1 |
10/67/308 |
||
|
- |
(0.3) |
(2.3) |
(7.8) |
(15.6) |
(24.9) |
(33.0) |
(11.9) |
(3.1) |
(0.8) |
(0.3) |
|||
|
БаИ |
мт.с. |
0/0/1 |
1/5/13 |
1/14/14 |
0/20/26 |
0/22/28 |
0/8/23 |
0/4/10 |
2/73/115 |
||||
|
- |
(0.5) |
(10.0) |
(15.3) |
(24.2) |
(26.3) |
(16.3) |
(7.4) |
- |
- |
- |
|||
|
ряпушка |
0/0/1 |
0/17/6 |
0/40/15 |
0/12/1 |
0/1/1 |
0/0/1 |
0/70/25 |
||||||
|
(1.1) |
(24.3) |
(57.9) |
(13.7) |
(2.0) |
- |
(1.0) |
- |
- |
- |
- |
|||
|
корюшка |
1/0/0 |
0/0/4 |
0/4/24 |
0/4/35 |
0/0/8 |
0/0/2 |
0/0/2 |
1/8/75 |
|||||
|
- |
- |
(1.2) |
(4.8) |
(33.3) |
(46.4) |
(9.5) |
(2.4) |
(2.4) |
- |
- |
|||
|
Оз. |
ст.с. |
0/1/0 |
0/2/2 |
0/5/6 |
0/0/3 |
0/1/0 |
0/9/11 |
||||||
|
Имандра |
- |
- |
(5.0) |
(20.0) |
(55.0) |
(15.0) |
(5.0) |
- |
- |
- |
- |
||
Половой состав. В половой структуре малотычинкового сига из БаИ и ЙИ доминировали самки (табл. 4). Соотношение самцов и самок в обоих плесах составило в среднем 1:1.5. В уловах из БоИ самцов малотычинкового сига было больше - 2:1. У среднетычинкового сига количество самцов и самок в выборках было почти одинаковым (1:1). В уловах ряпушки из плëса БаИ преобладали самцы, соотношение самцов и самок составило 3:1, в то время как в ЙИ и БоИ - самок: 1:2. В половой структуре корюшки Имандры доминировали самки. Соотношение полов в БаИ составило 1:9, в ЙИ и БоИ - 1:5.
Особенности линейно-весовых показателей. У малотычинкового и среднетычинкового сига оз. Имандра средние наблюденные длина и масса самцов и самок в течение жизни значимо не различались. У ряпушки и корюшки различия в этих показателях были незначительные. Далее приведены обобщенные размерно-весовые характеристики исследуемых видов рыб плёсов БоИ, ЙИ и БаИ.
Выборка малотычинкового сига из БоИ была представлена особями длиной 212-422 (334 ± 4.8) мм и массой 109-1285 (535 ± 24.7) г., из ЙИ - соответственно 113–500 (272 ± 2.1) мм и 15–1660 (255 ± 7.0) г; из БаИ - 150-436 (277 ± 4.1) мм и 29-1350 (282 ± 13.8) г. Средние величины длины и массы малотычинкового сига разных возрастных групп ЙИ и БаИ различались незначительно (в возрасте 3+, p < 0.001). Линейно-весовые показатели малотычинкового сига в БоИ были выше ( p < 0.001), чем в ЙИ и БаИ до возраста 5+ (табл. 5). Начиная с возраста 6+, размерновесовые показатели малотычинкового сига в разных плесах оз. Имандра значимо не различались.
Таблица 5
Средние наблюденные длина ( АС ), мм и масса ( W ), г у малотычинкового (мт.с.) и среднетычинкового (ст.с.) сига Coregonus lavaretus , европейской ряпушки C. albula и европейской корюшки Osmerus eperlanus в плёсах оз. Имандра, 2011 - 2016 гг.
|
Виды рыб |
Возраст, лет |
||||||||||
|
0+ |
1+ |
2+ |
3+ |
4+ |
5+ |
6+ |
7+ |
8+ |
9+ |
10+ |
|
|
АС |
|||||||||||
|
плёс Большая Имандра |
|||||||||||
|
мт.с. |
- |
220 |
248 ± 14.1 |
313 ± 8.8 |
344 ± 5.0 |
358 ± 7.4 |
338 ± 18.3 |
319 ± 13.5 |
- |
- |
- |
|
ряпушка |
99 ± 1.5 |
100 ± 1.5 |
112 ± 0.7 |
123 ± 1.5 |
149 ± 1.5 |
162 ± 2.7 |
- |
- |
- |
- |
- |
|
корюшка |
- |
- |
117 ± 3.5 |
131 ± 2.6 |
145 ± 2.0 |
155 ± 3.0 |
165 ± 2.8 |
175 ± 4.0 |
180 ± 6.3 |
196 ± 1.0 |
- |
|
плёс Йокостровская Имандра |
|||||||||||
|
мт.с. |
116 ± 2.5 |
166 ± 4.3 |
198 ± 4.2 |
257 ± 2.5 |
276 ± 1.9 |
302 ± 3.5 |
318 ± 6.5 |
329 ± 11.2 |
334 ± 10.5 |
380 ± 18.5 |
- |
|
ряпушка |
81 ± 1.9 |
103 ± 1.3 |
111 ± 2.8 |
119 ± 1.8 |
149 ± 4.1 |
180 ± 6.7 |
208 |
- |
- |
- |
- |
|
корюшка |
- |
94 |
104 ± 3.1 |
123 ± 2.7 |
143 ± 2.9 |
157 ± 2.3 |
178 ± 2.2 |
187 ± 3.3 |
200 ± 7.2 |
231 ± 1.5 |
216 |
Окончание табл. 5
|
Виды рыб |
Возраст, лет |
||||||||||
|
0+ 1 |
1+ 1 |
2+ 1 |
3+ 1 |
4+ 1 |
5+ 1 |
6+ 1 |
7+ 1 |
8+ 1 |
9+ 1 |
10+ |
|
|
плëс Бабинская Имандра |
|||||||||||
|
мт.с. |
- |
165 |
185 ± 5.7 |
224 ± 5.6 |
269 ± 4.6 |
301 ± 4.1 |
322 ± 5.8 |
345 ± 11.6 |
- |
- |
- |
|
ряпушка |
99 |
107 ± 1.6 |
110 ± 2.5 |
102 ± 2.3 |
197 ± 1.8 |
- |
160 |
- |
- |
- |
- |
|
корюшка |
- |
- |
91 |
123 ± 10.1 |
173 ± 5.1 |
180 ± 2.7 |
207 ± 8.8 |
193 ± 12.5 |
218 ± 7.5 |
- |
- |
|
Оз. Имандра |
|||||||||||
|
ст.с |
- 1 |
- 1 |
308 |
290 ± 17.1 |
289 ± 9.9 |
336 ± 23.01 |
- 1 |
337 |
- 1 |
- 1 |
- |
|
W |
|||||||||||
|
плëс Большая Имандра |
|||||||||||
|
мт.с. |
- |
131 |
184 ± 33.6 |
437 ± 36.9 |
556 ± 27.8 |
662 ± 51.0 |
611±139.4 |
477 ± 68.0 |
- |
- |
- |
|
ряпушка |
7 ± 0.3 |
9 ± 0.4 |
12 ± 0.3 |
17 ± 0.8 |
32 ± 1.1 |
42 ± 2.1 |
- |
- |
- |
- |
- |
|
корюшка |
- |
- |
11 ± 1.7 |
17 ± 1.2 |
22 ± 1.2 |
27 ± 2.0 |
31 ± 2.0 |
38 ± 2.7 |
45 ± 5.3 |
53 ± 12.2 |
- |
|
1.плëс Йокостровская Имандра |
|||||||||||
|
мт.с. |
16 ± 0.6 |
47 ± 4.4 |
86 ± 6.6 |
202 ± 7.7 |
247 ± 6.5 |
342 ± 17.4 |
409 ± 34.5 |
502 ± 66.3 |
433 ± 83.0 |
715 ± 85.0 |
- |
|
ряпушка |
4 ± 0.5 |
9 ± 0.4 |
11 ± 0.4 |
15 ± 0.9 |
31 ± 2.6 |
58 ± 5.8 |
81 |
- |
- |
- |
- |
|
корюшка |
- |
6 |
8 ± 0.9 |
12 ± 0.9 |
19 ± 1.6 |
26 ± 1.4 |
43 ± 2.2 |
51 ± 2.9 |
56 ± 5.1 |
112 ± 22.5 |
91 |
|
плëс Бабинская Имандра |
|||||||||||
|
мт.с. |
- |
50 |
63 ± 6.6 |
119 ± 9.0 |
223 ± 14.6 |
335 ± 16.2 |
428 ± 29.8 |
560 ± 76.9 |
- |
- |
- |
|
ряпушка |
6 |
8 ± 0.3 |
11 ± 1.4 |
9 ± 0.5 |
72 ± 12.5 |
- |
35 |
- |
- |
- |
- |
|
корюшка |
- |
- |
4 |
11 ± 3.6 |
39 ± 3.3 |
43 ± 2.1 |
65 ± 8.3 |
54 ± 11.0 |
74 ± 11.0 |
- |
- |
|
Оз. Имандра |
|||||||||||
|
ст.с |
- 1 |
- 1 |
315 1 |
339 ± 52.8 |
327 ± 36.71 |
579±117.5 |
- 1 |
529 |
- 1 |
- 1 |
- |
Среднетычинковые сиги в уловах из оз. Имандра встречались длиной 250–382 (299 ± 9.1) мм и массой 186–784 (377 ± 34,3) г. Линейновесовые показатели среднетычинкового сига в разных возрастах наиболее близки к показателям малотычинкового из плёса БоИ (табл. 5).
Ряпушка из БоИ в целом по выборке была представлена особями длиной: 86–180 (123 ± 1.1) мм и массой 5–51 (18 ± 0.6) г, из ЙИ и БаИ – соответственно длиной 58–208 (114 ± 1.2) мм и 89–214 (110 ± 2.0) мм и массой 2–86 (14 ± 0.7) г и 6–84 (11 ± 1.1) г. Различия средних значений длины и массы ряпушки в разных возрастах из трех плесов были незначительны (табл. 5): в возрасте 3+ рыбы из БаИ имели меньшие характеристики, нежели из ЙИ и БоИ ( p < 0.01 и p < 0.001 соответственно).
Выборка корюшки из плёса БаИ в целом была представлена особями значимо большей ( p < 0.01) длины: 91–240 (180 ± 2.4) мм и массы: 4–97 (43 ± 1.6) г по сравнению с корюшкой из плёсов ЙИ и БоИ, где рыбы имели соответственно длину 92– 255 (163 ± 1.3) мм и 110–225 (159 ± 1.7) мм, и массу 5–201 (34 ± 0.9) г и 6–84 (30 ± 1.1) г. Это связано с тем, что, в возрастах 4+–6+ корюшки из БаИ росли быстрее ( p < 0.01) по сравнению с корюшкой из других плесов озера (табл. 5). Начиная с возраста 6+ лет, у корюшки из плёса ЙИ наблюдаются бóльшие ( p < 0.05) линейно-весовые показатели по сравнению с корюшками из БоИ.
Созревание. Доля особей малотычинкового сига, готовящихся к нересту, в БоИ составила 43%, в ЙИ и БаИ – соответственно 17 и 16%. В БоИ половозрелые самцы встречались в возрасте от 3+ до 7+, самки – от 4+ до 6+. Модальный возраст созревания у первых соответствовал 4+, у вторых – 4+–5+. В ЙИ самцы и самки малотычинкового сига вступали в нерест соответственно в возрасте 3+–
9+ и 3+–8+, модальный возраст у обоих полов соответствовал 4+–5+. В БаИ половозрелые самцы малотычинкового сига встречались в возрасте от 4+ до 7+, самки – от 5+ до 7+, модальный возраст созревания у первых 5+, у вторых – 6+. В среднем линейно-весовые характеристики у половозрелых особей малотычинкового сига из БоИ были самыми большими по сравнению с теми же характеристиками в остальных плёсах и составили у самок 392 ± 12.1 (345–422) мм и 931 ± 109.8 (536–1285) г, у самцов – 356 ± 4.0 (302–395) мм и 600 ± 27.6 (248–900). В ЙИ половозрелые самцы малотычинкового сига среднем имели длину 321 ± 8.2 (260– 464) мм и массу 440 ± 46.1 (192–1295) г, самки – 302 ± 5.0 (228–460) мм и 358 ± 30.1 (114–1660) г. В БаИ половозрелые самцы и самки имели длину соответственно 342 ± 11.6 (275–436) и 298 ± 9.9 (245–395) мм и массу 568 ± 8.1 (241–1350) и 332 ± 42.4 (151–830) г.
В полученных выборках среднетычинкового сига оз. Имандры все особи имели II стадию зрелости гонад.
Доля половозрелых ряпушек в выборках из плёса БоИ составила 71%, в ЙИ и БаИ – соответственно 38 и 74%. Во всех плесах оз. Имандра половозрелые самцы ряпушки встречались в возрасте от 1+ до 4+, половозрелые самки – в ЙИ и БаИ – в возрасте 1+-6+, в БоИ – в возрасте 1+–5+. Модальный возраст созревания у самцов соответствовал 2+ –3+, у самок – 2+–4+. В среднем длина и масса половозрелых самцов составляла: в БоИ – 111 ± 0.9 (93–139) мм и 12 ± 0.3 (7–22) г., в ЙИ – 115 ± 2.4 (100–153) мм и 13 ± 1.3 (7–40) г, в БаИ – 106 ± 1.6 (89–179) мм и 9.5 ± 0.9 (6–59) г. Средняя длина и масса половозрелых самок составляла: в БоИ – 132 ± 1.7 (88–180) мм и 23 ± 1.0 (5–51) г, в ЙИ – 125 ± 2.5 (100–208) мм и 19 ± 1.5 (6–86) г, в
БаИ – 111 ± 5.2 (90–214) мм и 13 ± 3.3 (6–84) г.
Доля половозрелой корюшки в разных плёсах составила: в БоИ – 68%, в ЙИ и БаИ – 48%. В плёсе БоИ самцы корюшки созревали в возрасте от 2+ до 8+, в ЙИ – от 2+ до +7+, в БаИ – от 4+ до 5+. Модальный возраст созревания у самцов во всех плёсах 4+–6+. Самки корюшки созревали возрасте: в БоИ – 3+–9+, в ЙИ – 2+–10+, БаИ – 4+– 8+. Модальный возраст созревания самок, как и у самцов, наблюдался в 4–7+. В БоИ и ЙИ самцы корюшки созревали при близкой длине и массе, а именно 145–147 (105–192) мм и 23–24 г (5–-58) г. В БаИ эти показатели были выше: 171 ± 5.4 (135– 192) мм и 38 ± 3.7 (14–52) г. В БоИ самки корюшки при созревании имели среднюю длину 165 ± 2.4 (113–225) мм и 35 ± 1.6 (10–84) г, в ЙИ – 182 ± 1.6 (101–255) мм и 48 ± 1.6 (6–201) г, в БаИ – 190 ± 3.3 (117–240) мм и 38 ± 3.7 (14–52) г.
Таблица 6
Питание малотычинкового сига Coregonus lavaretus в летне-осенний период из различных плёсов оз. Имандра, 2011 - 2016 гг.
|
Компоненты |
Плëс Большая Имандра |
Плëс Йокостровская Имандра |
Плëс Бабинская Имандра |
||||||
|
Размерные группы, длина по Смиту ( АС ), мм |
Размерные группы, длина по Смиту ( АС ), мм |
Размерные группы, длина по Смиту ( АС ), мм |
|||||||
|
200-299 |
300-399 |
100-199 |
200-299 |
300-399 |
400-499 |
100-199 |
200-299 |
300-399 |
|
|
- |
P, % / F, % |
P, % / F, % |
P, % / F, % |
P, % / F, % |
P, % / F, % |
P, % / F, % |
P, % / F, % |
P, % / F, % |
P, % / F, % |
|
Зоопланктон |
- |
3.5 / 12.5 |
53.8 / 55.6 |
13.3 / 41.5 |
1.2 / 8.7 |
- |
20.1 / 25.0 |
0.2 / 8.9 |
0.05 / 1.7 |
|
Бентос: |
100 / 100 |
96.5 / 87.5 |
40.0 / 66.7 |
32.6 / 95.1 |
70.2 / 87.0 |
100 / 100 |
62.0 / 87.5 |
95.0 / 96.0 |
98.1 / 100 |
|
поденки (лич.) |
- |
- |
- |
- |
- |
- |
- |
- |
0.05 / 1.7 |
|
веснянки (лич.) |
- |
- |
- |
- |
- |
- |
- |
- |
- |
|
ручейники (лич.) |
- |
0.3 / 12.5 |
- |
14.5 / 7.3 |
8.5 / 21.7 |
66.8 / 50.0 |
- |
29.7 / 31.1 |
67.0 / 55.9 |
|
хирономиды (лич.) |
- |
0.1 / 25.0 |
18.1 / 44.4 |
1.8 / 31.7 |
17.9 / 34.8 |
- |
38.4 / 75.0 |
1.1 / 26.7 |
0.3 / 6.8 |
|
перепончатокрылые (имаго) жуки (имаго) |
- |
0.2 / 9.8 |
- |
- |
9.0 / 12.5 |
0.5 / 2.0 |
0.2 / 1.7 |
||
|
- |
- |
- |
- |
- |
0.1 / 2.0 |
0.1 / 3.4 |
|||
|
водные клещи (имаго) |
- |
0.1 / 7.3 |
- |
- |
- |
0.1 / 2.0 |
- |
||
|
бокоплавы (имаго) |
- |
- |
- |
- |
- |
1.5 / 1.7 |
|||
|
моллюски |
26.9 / 100 |
93.8 / 87.5 |
21.9 / 44.4 |
16.2 / 63.4 |
43.7 / 73.9 |
33.2 / 100 |
14.6 / 25.0 |
63.5 / 84.4 |
29.5 / 78.0 |
|
олигохеты |
- |
2.3 / 12.5 |
- |
- |
- |
- |
- |
- |
|
|
Рыба |
- |
- |
- |
2.1 / 2.4 |
- |
0.6 / 2.2 |
0.8 / 1.7 |
||
|
Икра рыб |
- |
- |
- |
50.0 / 26.8 |
28.6 / 17.4 |
- |
- |
- |
- |
|
Остатки растений |
73.1 / 50.0 |
- |
- |
- |
- |
- |
- |
- |
- |
|
Аморфная масса |
- |
- |
6.2 / 11.1 |
- |
- |
- |
17.9 / 12.5 |
4.2 / 13.3 |
- |
|
Непищевые компоненты (гальки, домики ручейников) |
- |
- |
- |
- |
- |
- |
- |
- |
0.5 / 1.7 |
|
Средний размер кормовых организ- |
- |
7.3 |
5.0 |
6.6 |
7.4 |
12.1 |
5.2 |
5.8 |
6.9 |
|
мов, мм |
|||||||||
|
Средняя масса пи- |
80.0 |
2062.4 |
176.3 |
1772.1 |
2314.6 |
2570.0 |
203.6 |
999.1 |
2979.0 |
|
щевого комка. мг |
|||||||||
|
Средний Iн, 0/ 000 |
3.3 |
48.3 |
24.4 |
79.0 |
63.9 |
69.3 |
25.4 |
47.5 |
67.4 |
|
Количество экз. |
2 |
8 |
9 |
41 |
23 |
2 |
8 |
45 |
59 |
Примечания: Здесь и в табл. 7: F , % – частота встречаемости компонента; P , % – доля компонента пищи по массе; зоопланктон – Acanthocyclops sp., Bythotrephes sp., Cyclops sp., Eurycercus sp.; бентос – поденки: Centroptilum sp; ручейники: Agraylea sp., Athripsodes sp., Molana sp., Oxyethira sp., Phryganea sp.; хирономиды: Ablabesmyia sp., Chironomus sp., Cricotopus sp., Endochironomus sp., Hyptotendipes sp., Macropelapia sp., Mallochohelea sp., Micropsectra sp.; Polipedium sp., Procladius sp., Prodiamesa sp.; жуки: Platambus sp.; двустворчатые моллюски: Euglesa sp., Sphaerium sp.; брюхоногие моллюски: Limnea sp., Valvata sp., рыба: Pungitius pungitius .
Таблица 7
Питание европейской ряпушки Coregonus albula в летне-осенний период из различных плёсов оз. Имандра, 2011 - 2016 гг.
|
Компоненты |
Плëс Большая Имандра |
Плëс Йокостровская Имандра |
Плëс Бабинская Имандра |
|||||
|
Размерные группы, длина по Смиту ( АС ), мм |
Размерные группы, длина по Смиту ( АС ), мм |
Размерные группы, длина по Смиту ( АС ), мм |
||||||
|
101-150 |
151-200 |
0-100 |
101-150 |
151-200 |
201-250 |
0-100 |
101-150 |
|
|
- |
P, % / F, % |
P, % / F, % |
P, % / F, % |
P, % / F, % |
P, % / F, % |
P, % / F, % |
P, % / F, % |
P, % / F, % |
|
Зоопланктон |
18.3 / 25.0 |
11.2 / 20.0 |
34.2 / 63.6 |
59.8 / 47.8 |
5.3 / 25.0 |
90.1 / 50.0 |
100 / 100 |
100 / 100 |
|
Ветвистоус. рачки |
6.5 / 16.7 |
11.2 / 20.0 |
34.2 / 63.6 |
52.6 / 52.6 |
5.3 / 25.0 |
90.1 / 50.0 |
100 /100 |
100 / 100 |
|
веслоногие рачки |
11.8 / 8.3 |
- |
- |
7.2 / 13.0 |
- |
- |
- |
|
|
Бентос: |
53.0 / 66.7 |
87.1 / 100 |
64.4 / 36.4 |
37.9 / 43.5 |
94.7 / 75.0 |
9.9 / 50.0 |
- |
- |
|
поденки |
- |
- |
- |
- |
1.2 / 12.5 |
- |
- |
- |
|
веснянки (имаго) |
- |
0.6 / 20.0 |
- |
- |
- |
- |
- |
- |
|
ручейники (кук.) |
0.8 / 8.3 |
17.1 / 20.0 |
- |
5.4 / 8.7 |
44.2 / 37.5 |
- |
- |
- |
|
двукрылые (лич., кук.) |
13.4 / 41.7 |
42.9 / 70.0 |
64.4 / 36.4 |
32.5 / 39.1 |
49.3 / 37.5 |
- |
- |
- |
|
моллюски |
0.3 / 8.3 |
0.9 / 10.0 |
- |
- |
- |
9.9 / 50.0 |
- |
- |
|
жесткокрылые (лич.) |
22.4 / 16.7 |
11.8 / 20.0 |
- |
- |
- |
- |
- |
- |
|
клопы (имаго) |
- |
1.3 / 10.0 |
- |
- |
- |
- |
- |
- |
|
пиявки |
4.7 / 16.7 |
12.5 / 10.0 |
- |
- |
- |
- |
- |
- |
|
круглые черви |
1.6 / 8.3 |
- |
- |
- |
- |
- |
- |
- |
|
бокоплавы |
9.8 / 8.3 |
- |
- |
- |
- |
- |
- |
- |
|
Аморфная масса Средний размер |
28.7 / 25.0 |
1.7 / 20.0 |
1.4 / 9.1 |
2.3 / 4.3 |
- |
- |
- |
- |
|
кормовых организмов, мм |
7.2 |
8.2 |
3.1 |
4.7 |
10.1 |
3.6 |
0.5 |
0.5 |
|
Средняя масса пищевого комка, мг |
110.8 |
135.5 |
17.5 |
43.0 |
313.4 |
199.0 |
25.0 |
9.0 |
|
Средний Iн, 0/ 000 |
47.5 |
34.6 |
41.2 |
32.5 |
58.1 |
23.1 |
41.7 |
10.0 |
|
Количество экз. |
12 |
10 |
11 |
23 |
8 |
2 |
1 |
1 |
Примечание. Зоопланктон – ветвистоусые рачки: Bosmina sp., Bythotrephes sp., Daphnia sp., веслоногие рачки – Acantocyclops sp., Сyclops sp.; бентос – хирономиды: Ablabesmyia sp., Centroptilum sp., Cricotopus sp., Diamesa sp., Endochironomus sp., Luttipelopia sp., Prodiamesa sp., Psectrocladius sp., Strictochironomus sp.; ручейники: Rhyacophila sp., Limnephillidae , Leptoceridae , Phryganeidae ; моллюски: двустворчатые – Euglesa sp., брюхоногие – Valvata sp., жесткокрылые: Dytiscus sp., клопы: Corixidae , бокоплавы: Gammarus sp.
Обсуждение
Основу рыбной части сообщества оз. Имандра в середине прошлого столетия составляли ряпушка и сиг (см. табл. 3). В уловах повсеместно встречался арктический голец и кумжа [Галкин, Колюшев, Покровский, 1966; Беляева, Чижиков, Долнин, 1969]. В БоИ в 1960-х гг. в уловах численность сига составляла от 40 до 73%, арктического гольца – 19–31%. На долю корюшки (снеток) в этот период приходилось около 0.5% [Беляева, Чижиков, Дол-нин, 1969]. В начале 1970-х гг. в БаИ сиг в уловах по численности составлял 37.8–56.5%, голец – 33.7–44.6%. Для ЙИ в указанный период встречаемость сига в уловах соответствовала 44.7% [Чижиков и др., 1976]. Интенсификация процессов промышленного загрязнения на протяжении последующих десятилетий в совокупности с неблагоприятным гидрологическим режимом (зимняя сработка воды до 3–4 м) и климатом, прекращение искусственного воспроизводства сига в 90-е гг. прошлого века, браконьерский вылов сига и гольца на нерестилищах, неразумные меры регулирования промыслом (временный запрет на вылов корюшки) привели к серьезным нарушениям типичной структуры рыбной части сообщества оз. Имандры. В настоящее время произошло снижение численности сига, критически сократилась доля лососевых видов [Антропогенные…, 2002; Моисеенко и Яковлев, 1990; Решетников, Терещенко, Лукин, 2011]). Численность хищных видов рыб также крайне низка (см. табл. 3). При этом успешная стратегия воспроизводства в реках, исключающая негативное влияние флуктуаций гидрологического и гидрохимического режимов оз. Имандры, достаточная обеспеченность кормовыми ресурсами, обеспечивают европейской корюшке доминирование в структуре сообщества. Следует отметить, что численность вида стабильно растет, несмотря на ее практически неограниченный вылов в период нереста. Кроме того, для корюшки в последние годы отмечено активное саморасселение в крупных речных системах бассейна оз. Имандры [Терентьев, Кашулин, Зубова, 2017].
В целом, современное «рыбное ядро» оз. Имандры формируют три вида: сиг, ряпушка и корюшка. При наблюдаемых сукцессионных перестройках обнаруживается ряд изменений в их биологических ха- рактеристиках. Сравнение полученных данных с литературными указывает на увеличение числа возрастных групп у корюшки и ряпушки оз. Имандры [Галкин, Колюшев, Покровский, 1966; Смирнов, 1977; Моисеенко, Яковлев, 1990; Антропогенные…, 2002], в то время как тенденция к омоложению стада малотычинкового сига сохраняется во всех плесах озера и в настоящем [Крогиус, 1926а; Решетников, 1966, 1980; Галкин и др., 1966; Комплексное…, 1969; Моисеенко, Яковлев, 1990; Антропогенные..., 2002; Зубова, 2015].
Такими же причинами можно объяснить и незначительное увеличение возраста у ряпушки. Омоложение популяции такого олиготоксобного вида, как сиг, связано, в первую очередь, с длительным негативным воздействием на озеро горнопромышленного комплекса [Моисеенко, Яковлев, 1990; Антропогенные…, 2002; Зубова, 2015]. Тем более, современные концентрации меди, никеля и стронция в воде и в поверхностном слое донных отложений всех плëсов оз. Имандра превышают фоновые значения, что говорит о сохраняющемся высоком уровне техногенной нагрузки на водоем [Зубова, Кашулин, Терентьев, 2015; Даувальтер, Кашулин, 2016].
Современное соотношение самцов и самок (далее полов) у малотычинкового сига в ЙИ и БаИ (1:1.5), а также у ряпушки в БоИ и ЙИ (1:2) характерно для преднерестовых группировок этих видов в оз. Имандра [Комплексное…, 1969]. Преобладание самцов над самками у малотычинкового сига в БоИ и у ряпушки в БаИ, выловленных в сентябре-октябре, может указывать на то, что вылов рыб производился в нерестовом стаде. Половая структура корюшки оз. Имандра также сильно изменилась за последние 60 лет, как и возрастная. Так, если в период с 1982 по 1997 гг. соотношение полов в ЙИ составляло в среднем 1:2 [Антропогенные…, 2002], то современное их соотношение в плёсах БоИ и ЙИ составляет в среднем 1:5, а в БаИ - 1:9. Таким образом, мы наблюдаем увеличение числа самок в популяции корюшки Имандры, что, возможно, говорит о большей выживаемости данного пола в современных условиях озера и большом потенциале популяции к увеличению численности. Также описываемую разницу возможно объяснить тем, что применялись разные способы сбора материала. Соотношение у среднетычинкового сига в оз. Имандра (1:1), возможно, соответствует его природному состоянию.
Общие линейно-весовые показатели выборки малотычинкового сига озера на сегодняшний день имеют наименьшие значения, начиная с 1930 г.: так, средняя длина рыб в возрасте от 3+до 8+ уменьшилась в среднем от 379 до 280 мм, масса -от 720 до 290 г, что связано с омоложением популяции малотычинкового сига и доминированием особей более раннего возраста [Крогиус, 1926а; Галкин, Колюшев, Покровский, 1966; Антропогенные..., 2002]. Как было показано ранее, большие показатели линейных характеристик у малотычинкового сига плëса БоИ до возраста 5+ объясняются более высокими их темпами роста в первый год жизни в этом плесе и связаны здесь с более высокой биомассой как зоопланктонных, так и зообентосных организмов, входящих в состав пищи рыб [Зубова, 2015; Зубова, Кашулин, Терентьев, 2015]. Средние линейно-весовые показатели среднетычинкового сига наиболее близки к показателям малотычинкового сига плеса БоИ, где он в основном и обитает.
Средняя длина и масса выборок корюшки и ряпушки в разных плесах оз. Имандра за последние годы сильно не изменились, при этом увеличились максимальные значения этих характеристик [Галкин, Колюшев, Покровский, 1966; Антропогенные…, 2002]. У ряпушки максимальные значения длины и массы выросли от 165 до 214 мм и от 40
до 86 г соответственно, у корюшки от 245 до 255 мм и от 110 до 201 г соответственно. Рост линейно-весовых характеристик ряпушки и корюшки можно объяснить тем, что эти виды рыб начали доживать до более старших возрастных групп, нежели ранее. Наблюдаемая разница в линейновесовых характеристиках корюшки из разных плесов оз. Имандры в некоторых возрастах требует более детального изучения их темпов роста в дальнейшем.
Доля половозрелых рыб у малотычинкового сига увеличивается от чистых вод БаИ к загрязненным водам БоИ. В плесе БаИ самцы и самки малотычинкового сига начинают созревать в возрасте 4+ и 5+ соответственно. В БоИ и ЙИ, как самцы, так самки, созревают раньше – в возрасте 3+–4+. Учитывая неравномерность вступления в нерестовое стадо, нерегулярность нереста и общую продолжительность жизни малотычинкового сига оз. Имандра, большая их часть успевает лишь один раз участвовать в нересте. Большинство особей впервые нерестящегося малотычинкового сига оз. Имандра в наших уловах достигали линейных и весовых размеров, которые характерны для созревающего сига из чистых водоемов: самцы – 27–30 см и 200–350 г, самки – 28–30 см и 230–400 г [Решетников, 1966, 1980; Решетников, Богданов, 2011], несмотря на снижение возраста начала созревания самцов и самок в БоИ и ЙИ по сравнению с теми же характеристиками в БаИ. Связано это с более высокими темпами роста малотычинкового сига в первых двух плесах в первый год жизни [Зубова, 2015; Зубова, Кашулин, Терентьев, 2015]. Таким образом, снижения размерных характеристик впервые нерестящихся малотычинковых сигов в более загрязненных водах ЙИ и БоИ, по сравнению с чистыми водами БаИ, не происходит. Здесь не встречались особи (в частности, самки), нерестящиеся в текущем году при длине 20 см, что, например, можно было наблюдать у малотычинкового сига в период максимального загрязнения оз. Имандра с 1982 по 1992 гг. [Моисеенко, 1997, 2002]. Следует учитывать ухудшающиеся условия воспроизводства сига. Значительные зимние сработки воды в сочетании с высокими уровнями загрязнения делают малопригодными озерные нерестовые участки. Воспроизводство сига до 90-х гг. прошлого века шло в основном за счет деятельности рыбоводного завода «Имандра», размножения озерно-речных форм и мигрантов из придаточных водоемов. Закрытие рыбоводного завода, агрессивная «оккупация» пригодных для нереста рек корюшкой и расширяющаяся ее экспансия на придаточные системы, ухудшение структуры популяции сига делают перспективы его выживания весьма пессимистическими. Если не будут предприняты меры по исправлению ситуа- ции, сига оз. Имандры в ближайшие годы ждет судьба арктического гольца.
Возраст начала и массового созревания особей ряпушки в оз. Имандра согласуется с природными: соответственно 1+–2+ и 2+–4+. Нами не обнаружены рыбы, готовые к нересту на первом году жизни (0+), как это наблюдалось в период интенсивной техногенной нагрузки на озеро (1982–1984 гг.) [Антропогенные..., 2002]. Линейно-весовые характеристики половозрелых ряпушек оз. Имандры ранее не описывались и, возможно, приведенные нами современные их показатели также соответствуют природным: средняя длина 11–13 см и масса 10–23 г.
По литературным данным [Антропогенные…, 2002], в период с начала и до середины 1990-х гг. популяция корюшки оз. Имандры приобретает структуру, свойственную снетковому типу (мелкой форме европейской корюшки): в уловах появляются половозрелые особи в возрасте 1+ длиной 108– 115 мм. В настоящее время корюшка в оз. Имандра начинает созревать в возрасте 2+ лет, массово – в возрасте 4+-5+ лет. Половозрелые самцы в возрасте от 2+ до 5+ лет имели длину от 110 до 196 (в среднем 151) мм и массу от 8 до 53 г (29 г), половозрелые самки того же возрастного интервала имели длину от 117 до 208 (165) мм и массу – от 21 до 70 г (34) г. Созревание корюшки различных плесов различалось и в более эвтрофной БоИ начиналось в возрасте 2+ (50% особей), в то время как в БаИ первые половозрелые ооби встречались только в возрасте 4+. При этом крупные особи, как правило, все были половозрелыми, что, вероятно, связано с переходом на хищничество. Таким образом, минимальные размеры впервые созревающих корюшек, начиная с 1990-х гг. по 2015 г., не изменились и, возможно, соответствуют снетковому типу, при этом средние размеры созревающих рыб соответствуют размерам крупной формы европейской корюшки (или просто корюшки) [Иванова, 1968; Антропогенные…, 200]). В подразделении форм на крупную (корюшку) и мелкую (снеток) основным критерием различия являются их размеры и особенности созревания [Иванова, 1982]. Считается, что длина тела снетка не превышает 10 см, тогда как корюшка становится половозрелой при длине 15 см. В то же время В.В. Петров (1940) указывал, что «резкой границы между корюшками и снетками не существует» [цит. по: Антропогенные…, 2002, стр. 297], а их биологические особенности в основном зависят от места обитания рыб. Мы считаем, что при разделении корюшки на мелкую и крупную формы необходимо учитывать комплекс таких биологических характеристик рыб, как возрастной состав и размеры, а также особенности их полового созревания. На основе выше описанных биологических характеристик корюш- ки, европейскую корюшку оз. Имандры в настоящее время нельзя относить к снетку с коротким циклом развития. Присутствие в популяции возрастных групп 8+ – 10+, неоднократно участвующих в нересте, свидетельствует о длинноцикловой стратегии развития.
Среди первоочередных мер по стабилизации ситуации необходимо восстановить деятельность рыбоводного завода, возобновив искусственное воспроизводство не только сига, но и гольца и кумжи. Учитывая, что нерест корюшки происходит в относительно короткий период в небольшом количестве рек, возможна организация эффективного регулирования ее численности. Но главной проблемой озера продолжают оставаться его загрязнение, эвтрофирование и неразумный водный режим. Решение всех этих проблем лежит целиком в области администрирования и управления природными ресурсами.
Работа выполнена в рамках темы НИР № 02262019-0045 и частично при поддержке гранта РФФИ 18-05-60125 Арктика и гранта РНФ № 1977-10007. Интерпретация результатов по гидрохимическим и гидробиологическим показателям выполнена при поддержке гранта РНФ № 19-77
10007; интерпретация результатов по ихтиологическим показателям – гранта РФФИ 18-05-60125 Арктика.
Список литературы Современные биологические характеристики сига Coregonus lavaretus, европейской ряпушки С. albula и европейской корюшки Osmerus eperlanus озера Имандра
- Антропогенные модификации экосистемы оз. Имандра / под ред. Т.И. Моисеенко. М.: Наука, 2002. 403 с.
- Атлас пресноводных рыб России / под ред. Ю.С. Решетникова: в 2 т. М.: Наука, 2003. Т. 1. 379 с.
- Беляева Г.В., Чижиков В.В., Долнин Т.В. Комплексное изучение и охрана оз. Большая Имандра. Апатиты: Изд-во Кольского научного центра, 1969. 133 с.
- Валькова, С.А. и др. Структура и динамика сообществ зообентоса оз. Имандра в зоне влияния медно-никелевого комбината // Труды Кольского научного центра РАН. 2012. № 1(2). С. 23-40.
- Вандыш О.И. Особенности зоопланктонного сообщества субарктического оз. Имандра в зонах техногенного воздействия (губы Монче, Белая, Молочная) // Экология. 2012. №. 5. С. 366-366.
- Галкин Г.Г., Колюшев А.А., Покровский В.В. Ихтиофауна водохранилищ и озер Мурманской области // Рыбы Мурманской области. Условия обитания, жизнь и промысел. Мурманск, 1966. С. 177-193.
- Даувальтер В.А., Кашулин Н.А. Изменение концентраций никеля и меди в поверхностных слоях донных отложений оз. Имандра за последние полвека // Вестник Мурманского государственного технического университета. 2015а. Т. 18, №. 2. С. 307-321.
- Даувальтер В.А., Кашулин Н.А. Влияние деятельности горно-металлургических предприятий на химический состав донных отложений оз. Имандра, Мурманская область // Биосфера. 2015б. Т. 7, №. 3. С. 295-314.
- Даувальтер В.А., Кашулин Н.А. Реконструкция накопления элементов в оз. Имандра как отражение динамики качества воды // Труды Ферсмановской научной сессии ГИ КНЦ РАН. 2016. № 13. С. 218-221.
- Даувальтер В.А., Терентьев П.М. Аккумуляция тяжелых металлов в донных отложениях и органах, и тканях сига (Coregonus 1ауагеШ') оз. Имандра // Труды Ферсмановской научной сессии ГИ КНЦ РАН. 2018. № 15. С. 445-448.
- Денисов Д.Б. Проблемы современной биоиндикации состояния субарктических водных экосистем на основе водорослевых сообществ // Биоиндикация в мониторинге пресноводных экосистем: материалы междунар. конф. СПб., 2011. С. 68-73.
- Денисов Д.Б., Кашулин Н.А. Цианопрокариоты в составе планктона оз. Имандра (Кольский полуостров) // Труды Кольского научного центра РАН. 2016. № 7 (41). С. 40-57.
- Заварзина Н.К. О методике определения возраста и размерно-возрастных особенностях малоротых корюшек рода НуротеБиБ острова Сахалин // Чтения памяти Владимира Яковлевича Леванидова. 2005. № 3. С. 585-593.
- Зубова Е.М. Линейный рост европейского сига Coregonus 1ауагеШ' (Ь.) в антропогенно-модифицированных водоемах европейской субарктики (на примере Мурманской области): автореф. дис. ... канд. биол. наук. Пермь, 2015. 28 с.
- Зубова Е.М., Кашулин Н.А., Терентьев П.М. Линейный рост малотычинкового сига Сoregonus 1ауагеШ' (Сoregonidae) оз. Имандра (Мурманская область) // Вопросы ихтиологии. 2015. Т. 56, № 4. С. 463-473.
- Зубова Е.М. и др. Новые данные по морфологическим особенностям жаберного аппарата мало- и сред-нетычинковой форм сига Coregonus 1ауагеШ' (Ь.) из крупнейшего субарктического озера // Биология внутренних вод. 2018. № 4. С. 63-74.
- Иванова М.Н. Популяционная изменчивость пресноводных корюшек // Труды Института биологии внутренних вод. 1968. Вып. 15 (53). 145 с.
- Методическое пособие по изучению питания и пищевых отношений рыб в естественных условиях / Е.В. Боруцкий [и др.]. М.: Наука, 1974. 254 с.
- Моисеенко Т.И. Теоретические основы нормирования антропогенных нагрузок на водоемы Субарктики. Апатиты, 1997. 261 с.
- Моисеенко Т.И. Изменение стратегии жизненного цикла рыб под воздействием хронического загрязнения вод // Экология. 2002. № 1. С. 50-60.
- Моисеенко Т.И., Яковлев В.А. Антропогенные преобразования водных экосистем Кольского Севера. Л.: Наука, 1990. 219 с.
- Кашулин Н.А. Рыбы малых озер Северной Фен-носкандии в условиях аэротехногенного загрязнения. Апатиты: Изд-во КНЦ РАН, 2004. 130 с.
- Кашулин Н.А. и др. Некоторые аспекты современного состояния пресноводных ресурсов Мурманской области // Вестник Мурманского государственного технического университета. 2013. Т. 16, № 1. С. 98-107.
- Комплексное изучение и охрана оз. Большая Имандра: отчет о НИР. Апатиты, Изд-во КНЦ РАН, 1969. 145 с.
- Крогиус Ф.В. Ихтиологические работы на оз Имандра // Работы Мурманской биологической станции. 1926а. Т. 2. С. 150-152.
- Крогиус Ф.В. Материалы по возрасту и темпу роста сига оз. Имандра // Работы Мурманской биологической станции. 1926б. Т. 2. С. 77-87.
- Крючков В.В., Моисеенко Т.И., Яковлев В.А. Экология водоемов-охладителей в условиях Заполярья. Апатиты: Изд-во КНЦ РАН, 1985. 131 с.
- Определитель зоопланктона и зообентоса пресных вод Европейской России / под ред. В.Р. Алексеева. М.; СПб.: Т-во науч. изд. КМК, 2016. Т. 2. Зообентос. 457 с.
- Решетников Ю.С. Особенности роста и созревания сигов в водоемах Севера // Закономерности динамики численности рыб Белого моря и его бассейна. М.: Наука, 1966. С. 93-155.
- Решетников Ю.С. Экология и систематика сиговых рыб. М.: Наука, 1980. 301 с.
- Решетников Ю.С., Богданов В.Д. Особенности воспроизводства сиговых рыб // Вопросы ихтиологии. 2011. Т. 51, № 4. С. 502-525.
- Решетников Ю.С., Терещенко В.Г., Лукин А.А. Динамика рыбной части сообщества в изменяющихся условиях обитания (на примере оз. Имандра) // Рыбное хозяйство. 2011. № 6. С. 48-51.
- Руководство по изучению питания рыб в естественных условиях / Карпевич А.Ф. [и др.]. М.: Изд-во АН СССР, 1961. 262 с.
- Руководство по гидробиологическому мониторингу пресноводных экосиситем. СПб.: Гидромет-издат, 1992. 318 с.
- Сидоров Г.П., Решетников Ю.С. Лососеобразные рыбы водоемов европейского северо-востока. М.: Т-во науч. изд. КМК, 2014. 346 с.
- Смирнов А.Ф. Рыбы озера Имандра // Рыбы озер Кольского полуострова: сб. науч. тр. Петрозаводск: Наука, 1977. С. 56-76.
- Терентьев П.М., Кашулин Н.А., Зубова Е.М. Роль европейской корюшки Osmerus eperlanus (Linnaeus) в структуре ихтиофауны бассейна оз. Имандра (Мурманская область) // Труды Зоологического института РАН. 2017. Т. 321, № 2. С. 228-243.
- Терентьева И.А., Кашулин Н.А., Денисов Д.Б. 2017. Оценка трофического статуса субарктического оз. Имандра // Вестник Мурманского государственного технического университета. 2017. Т. 20, № 1/2. С. 197-204.
- Чижиков В.В. и др. Окончательный отчет по теме «Комплексное изучение Йокостровской и Ба-бинской Имандры и разработка рекомендаций по рациональному использованию водоема» № 11 - 72 - 16 в соответствии с планом научно-исследовательских работ на 1971 - 1976 гг.: в 2 т. Апатиты: Изд-во КНЦ РАН, 1976. Т. 1. 276 с.
- Чижиков В.В. Гидрохимия и донные отложения оз. Имандра // Экосистема оз. Имандра под влиянием техногенного загрязнения: сб. науч. тр. Апатиты: Изд-во КНЦ РАН, 1980. С. 24-67.
- Berezina N.A., Strelnikova A.P., Maximov A.A. The benthos as the basis of vendace, Coregonus albu-la, and perch, Perca fluviatilis, diets in an oligo-trophic sub-Arctic lake // Polar Biology. 2018. № 41. pp. 1789-1799.
- Czarkowski T.K. et al. Feeding ecology of vendace, Coregonus albula (L.), in Lake Wigry (northeastern Poland) // Arch. Pol. Fish. 2007. № 15. Р. 117-128.
- Dauvalter V.A., Kashulin N.A. Mercury Pollution of Lake Imandra Sediments, the Murmansk Region, Russia // International Journal of Environmental Research. 2018. V. 12, №. 6. Р. 939-953.
- McKenzie R.A. Age and growth of smelt, Osmerus mordax (Mitchill), of the Miramichi River, New Brunswick // Journal of the Fisheries Research Board of Canada. 1957. № 15. Р. 1313-1327.
- Northcote T. G., Hammar J. Feeding ecology of Core-gonus albula and Osmerus eperlanus in the limnetic waters of Lake Malaren, Sweden // Boreal Env. Res. 2006. № 11. Р. 229-246.
- Van Oosten J. Life history of the lake herring (Leucichthys artedi Le Sueur) of Lake Huron as revealed by its scales, with a critique of the scale method // USA Bureau of Fisheries Bulletin. 1929. № 44. Р. 265-428.
- Vander Zanden M.J. , Vadeboncoeur Y. Fishes as integrators of benthic and pelagic food webs in lakes // Ecology. 2002. № 83. P. 2152-2161.
