Современные исследования ископаемого костного детрита: палеонтология, минералогия, геохимия

Автор: Силаев В.И., Пономарев Д.В., Симакова Ю.С., Шанина С.Н., Смолева И.В., Тропников Е.М., Хазов А.Ф.
Журнал: Вестник геонаук @vestnik-geo
Рубрика: Научные статьи
Статья в выпуске: 5 (257), 2016 года.
Бесплатный доступ
Охарактеризованы принципы и методы минералого-геохимических исследований ископаемого костного детрита в целях получения разнообразной информации о первичных биологических свойствах соответствующих организмов и об условиях и результатах фоссилизации их костных останков. Приведены примеры изучения микростроения костей (оптическая, сканирующая электронная, атомно-силовая зондовая микроскопия), нанопористости (по кинетике абсорбции-десорбции азота), химического состава (рентгенофлюоресцентный анализ), микроэлементов (масс-спектрометрия индуктивно-связанной плазмы), фазового состава биоминеральной компоненты (рентгеновская дифрактометрия, ИК-спектроскопия), элементного (газовая хроматография), молекулярно-группового (ИК-спектроскопия), аминокислотного (газовая хроматография) состава костного коллагена, изотопного состава С, О и N (изотопная масс-спектрометрия в соединении с газовой хроматографией). Сделан вывод о том, что лишь осознанное освоение и плодотворное комплексное использование множества прецизионных минералогических, физических, физико-химических и химических методов может обеспечить палеонтологии успешное будущее.
Ископаемые кости, минералого-геохимические исследования, генетическая информация
Короткий адрес: https://sciup.org/149129203
IDR: 149129203 | УДК: 552.321.6 | DOI: 10.19110/2221-1381-2016-5-19-31
Current research of fossil bone detritus: paleontology, mineralogy, geochemistry
We characterize the principles and methods of mineralogical and geochemical studies of fossil bone detritus in order to obtain a variety of information about the primary biological properties of the organisms and the conditions and results of the fossilization of their skeletal remains. Examples of studying the microstructure of the bone (optical, scanning electron, atomic force probe microscopy), and nano porous (kinetic nitrogen adsorption-desorption), chemical composition (X-ray fluorescence analysis), trace elements (mass spectrometry, inductively coupled plasma), phase composition biomineral components (X-ray diffraction, infrared spectroscopy), elemental (gas chromatography), a molecular group (IR spectroscopy), an amino acid (gas chromatography) of the composition of bone collagen, the isotopic composition of C, O and N (isotope ratio mass spectrometry in conjunction with gas chromatography) are discussed. It is concluded that the development of a conscious and fruitful use of multiple precision complex mineralogical, physical, physica-chemical and chemical methods can ensure a successful future of paleontology.
Текст научной статьи Современные исследования ископаемого костного детрита: палеонтология, минералогия, геохимия
«Вы никогда не решите проблему, если сохраните то же мышление и тот же подход, которые привели вас к этой проблеме» А. Эйнштейн, отрывок из письма
В настоящее время все большую актуальность приобретают междисциплинарные научные исследования, проблематика которых лежит на маргинальных границах между отдельными научными дисциплинами (рис. 1). Главной причиной возникновения междисциплинарности является стремление к паллиативу между объектами и методами исследований, формально относящимися к разным наукам, но при этом вполне сочетающимися в процессах исследований конкретных природных феноменов. Как известно, академик Н. П. Юшкин был весьма чуток к паллиативам такого рода, неслучайно став основоположником нескольких пограничных научных дисциплин, например, генетико-информационной минералогии, топоминералогии, витаминералогии, архео логической минералогии. Большой вклад, при этом, он сделал в исследования именно органоминеральных композиций как биогенного, так и абиогенного происхождения.
В области наук о Земле одним из наиболее очевидных объектов междисциплинарных исследований является ископаемый костный детрит животных, включая человека, который служит источником палеонтологической, палеоантропологической, археологической и палеоэкологической информации. Понятно, что получение такой информации на современном уровне может быть обеспечено только использованием высокотехнологичных аналитических методов [2, 4, 8, 12—18], уже доказавших свою эффективность в минералогии и геохимии
Объекты и методы исследований
Объектами наших исследований служат несколько коллекций и отдельных образцов костного детрита животных, включая человека, из местонахождений в Печорском Приуралье, Западной и Центральной Сибири, датирую- 19
щихся диапазоном от среднего неоплейстоцена (палеолита) до среднего-позднего средневековья (рис. 2). Авторами этих коллекций и образцов являются Д. В. Пономарев, А. А. Бондарев, С. М. Слепченко, П. А. Косинский, Г. В. Шнейдер, А. М. Клементьев, Д. В. Кобылкин.
Как известно, кости животных представляют собой очень сложный органоминеральный композит с на-
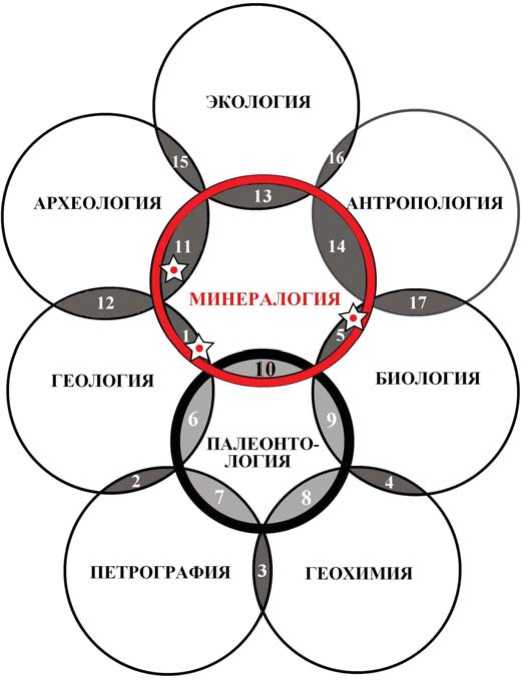
Рис. 1*. Схема образования пограничных научных дисциплин (показаны заливкой) в результате развития междисциплинарных научных исследований: 1 — топомине-ралогия, 2 — геопетрология, 3 — петрогеохимия, 4 — биогеохимия, 5 — биоминералогия, 6 — биостратиграфия, 7 — палеобиогеография, 8 — палеобиогеохимия, 9 — палеобиология, 10 — минералогия ископаемых организмов, 11 — археологическая минералогия, 12 — геоархеология, 13 — экоминералогия, 14 — антропоминералогия, 15 — экоархеология, 16 — экоантропология, 17 — биоантропология. Звездочками отмечены дисциплины, одним из основоположников которых был академик Н. П. Юшкин
Fig. 1. Scheme of boundary disciplines (filled) as a result of the development of interdisciplinary scientific researches: 1 — topomineralogy 2 — geopetrology, 3 — petrogeochemistry, 4 — biogeochemistry, 5 — biomineralogy, 6 — biostratigraphy 7 — paleobiogeography 8 — paleobiogeochemistry, 9 — paleobiology, 10 — fossil mineralogy, 11 — archaeological mineralogy, 12-geoarcheology, 13 — ecomineralogy, 14 — anthropomineralogy, 15 — ecoarcheology, 16 — ecoanthropology, 17 — bioanthropology. Asterisks show disciplines, which one of the founders was Academician N. P. Yushkin нометровой размерностью индивидов и пор, полноценное исследование которого возможно лишь при использовании широкого комплекса прецизионных методов. В число используемых нами методов входят: 1) оптическая (Полам Р-312 в сочетании с компьютеризированным комплексом OLYMPUS BX51), растровая электронная (JSM-6400 Jeol, Tescan Vega3 LMH) и зондовая атомно-силовая (NT-MDT JNTEGRA PRIMA) микроскопия — для изучения микростроения костей и выявления визуальных признаков их эпигенетического изменения (фоссилизации) ; 2) определение нанопористости в диапазоне от < 1 до 50 нм статистическим объемометрическим методом по кинетике адсорбции/десорбции азота (Nova 1200e) — для оценки степени сохранности органического матрикса в костях и состояния срастаний индивидов биоапатита и коллагена; 3) рентгенофлюоресцентный анализ (XRF-1800 Shimadzu) — определение валового химического состава костей для оценки характера и степени их загрязнения в результате иллювиирования вещества из вмещающих терригенных осадков; 4) определение содержания органического углерода методом кулонометрического титрования по величине pH (Ан-7529М) — оценка степени «выгорания» органического матрикса в костях при их фоссилиза-ции; 5) определение содержания микроэлементов методом ИСП-МС (Perkin Elmer ELAN 9000) — оценка степени эпигенетического изменения костей в зависимости от геохимических условий захоронения; 6) рентгеновская дифрактометрия (Shimadzu XRD-6000) — анализ фазового состава био- и ксеноминеральной компонент, определение рентгеноструктурных параметров костного биоапатита для оценки характера и степени его структурного и кристаллохимического изменения при фоссилизации; 7) инфракрасная спектроскопия биоминеральной компоненты (ИнфраЛюм ФТ-2) и коллагена (AVATAR 360 FT-IR с приставкой МНПВО «NICOLET Smart MIRacle») — определение структурной позиции карбонатной примеси в биоапатите и оценка характера и степени молекулярной деградации костного белка; 8) рентгеноспектральный ми-крозондовый анализ (JSM-6400 Jeol, Tescan Vega3 LMH с энергодисперсионной приставкой X-max50) — диагностика и определение химического состава и кристаллохимических особенностей биоапатита и ксеноминеральных примесей; 9) термический анализ (Shimadzu DTG-60A /60AH) — оценка степени сохранности органического матрикса в костях как критерия относительного их возраста; 10) рамановская спектроскопия (Renishaw InVia, лазеры 514 и 785 нм) — оценка степени карбонизации костного белка; 11) определение элементного состава коллагена (газовый хроматограф EA1110 (CHNO—O) — оценка степени химической деградации коллагена; 12) аминокислотный анализ (хроматограф GC-17A Shimadzu, капиллярная колонка Chirasil-L-Val) — определение аминокислотного состава костного коллагена и оценка степени его изменения в результате фоссилизации костей; 13) изотопная масс-спектрометрия раздельно по аминокислотам (DeltaVAdvantage (ThermoFinnigan) в соединении с газовым хроматографом TraceGC 2000 ThermoFinnigan (капиллярная колонка DB-5) — определение вариации изотоп-
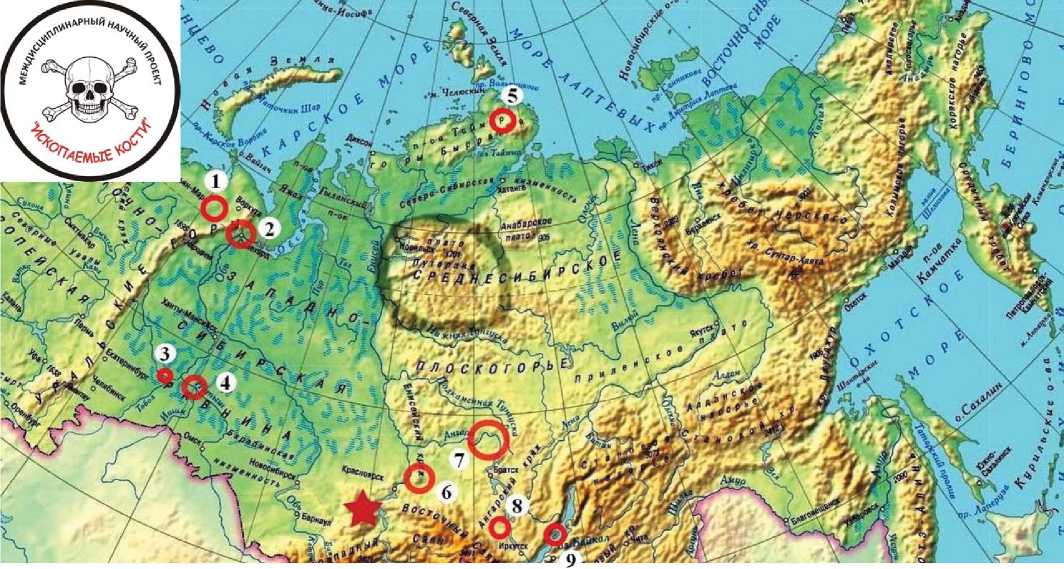
Рис. 2. Районы местонахождений исследуемого костного детрита крупных млекопитающих плейстоцена—голоцена: 1 — Печорское Приуралье (мамонтовая фауна, неоплейстоцен); 2 — святилище Усть-Полуй (человеческие захоронения, от позднего бронзового до раннего железного веков); 3 — Байгара (таранная кость человека, неолит); 4 — Среднее Прииртышье (мамонтовая фауна, неоплейстоцен; усть-шимская кость, палеолит; костный детрит тоболо-иртышского тюрка из Красноярского археологического комплекса, раннее-позднее средневековье); 5 — Северный Таймыр (мамонт, неоплейстоцен); 6 — Верховье Енисея (могильники, от неолита до средневековья); 7 — Северное Приангарье (могильники, от бронзового века до средневековья); 8 — Окинское плато (кость пещерной гиены, неоплейстоцен); 9 — оз. Ольхон (костные фрагменты миоценового носорога «Михалыча»). Звездой показан район расположения пещер Чагырской, А. П. Окладникова, Денисовской и Кара-Бом с костными останками гоминид палеолитического возраста (неандертальцы и денисовские люди)
Fig. 2. Localities of the studied bone detritus of large Pleistocene-Holocene mammals: 1 — Pechora Suburals (mammoths, Neopleistocene); 2 — Ust-Polui sanctuary (human burials from the Late Bronze to the Early Iron ages); 3 — Baygara (anklebone of "Tobolsk" man, Neolithic); 4 — Middle Irtysh (mammoths, Neopleistocene; Ust-Shim bone, Paleolithic; bone detritus of Tobol-Irtysh Turk from the Krasnoyarsk archeological complex, Early-Late Middle Ages); 5 — North Taimyr (mammoth, Neopleistocene); 6 — Upper Yenisei (burials, from Neolithic to Middle Ages); 7 — North Angara (burials, from Bronze Age to Middle Ages); 8 — Oka plateau (bone of cave hyena, Neopleistocene); 9 — Lake Olkhon (bone fragments of Miocene rhinoceros "Mikhalych"). The star shows the location of the caves: Chagyrskaya, Okladnikov, Denisov and Kara-Bom with bone remains of Paleolithic hominids (Neanderthalers and Denisov humans)
ного состава углерода в аминокислотах костного коллагена с оценкой тенденций и степени его вторичных изменений; 14) изотопная масс-спектрометрия биоапатита и коллагена (DeltaVAdvantage (ThermoFinnigan) соединенного с газовым хроматографом TraceGC 2000 (ThermoFinnigan, капиллярная колонка DB-5) — определение изотопного состава углерода, кислорода и азота в биоапатите и коллагене для оценки палеоклиматических и палеоэкологических условий существования животных.
Микростроение ископаемых костей
Костный детрит в зависимости от возраста варьируется по окраске от буровато-желтого до темно-бурого. Заметно колеблется и его механическая прочность, как правило, многократно снижающаяся с возрастом кости. Под оптическим микроскопом хорошо сохранившийся костный материал обнаруживает первичное концентри-чески-слоистое, или спутано-тонковолокнистое строение с локальными пустотами размером 200—800 мкм (рис. 3, а). Сильные вторичные изменения приводят к нарушениям сплошности первичной костной ткани вследствие механических деформаций и растворения, а также приводят к коллофанизации — замещению более или менее кристалличного биоапатита аморфной не прозрачной массой без существенного изменения химического состава. В последнем случае сохраняются только реликты первичного строения костной ткани (рис. 3, б). В режиме зондовой атомно-силовой микроскопии даже для хорошо сохранившихся костей наблюдается явная тенденция к последовательному изменению их структуры с возрастом, выражающаяся в постепенной утрате сплошности, нарастанию признаков дезинтеграции уже на субмикронно-мезонанометровом уровне, постепенном уменьшении размера элементов надмолекулярной структуры костей (рис. 3, в—д).
В сканирующем электронном микроскопе хорошо проявляются не только первичные свойства ископаемых костей, но и результаты их вторичных изменений. В частности отчетливо наблюдается ультрадисперсное оксиги-дроксидное ожелезнение костного биоапатита, обусловленное фоссилизацией (рис. 4, а—г). Кроме того, в крупных порах образуются агрегаты бактериогенного фрамбоидального пирита и сгустки более позднего гётита (рис. 4, д, е). Образование этих минералов явно происходило в два этапа — ранний восстановительный (суль-фатредукция и образование фрамбоидального пирита) и поздний окислительный (образование гётита, зачастую по пириту).
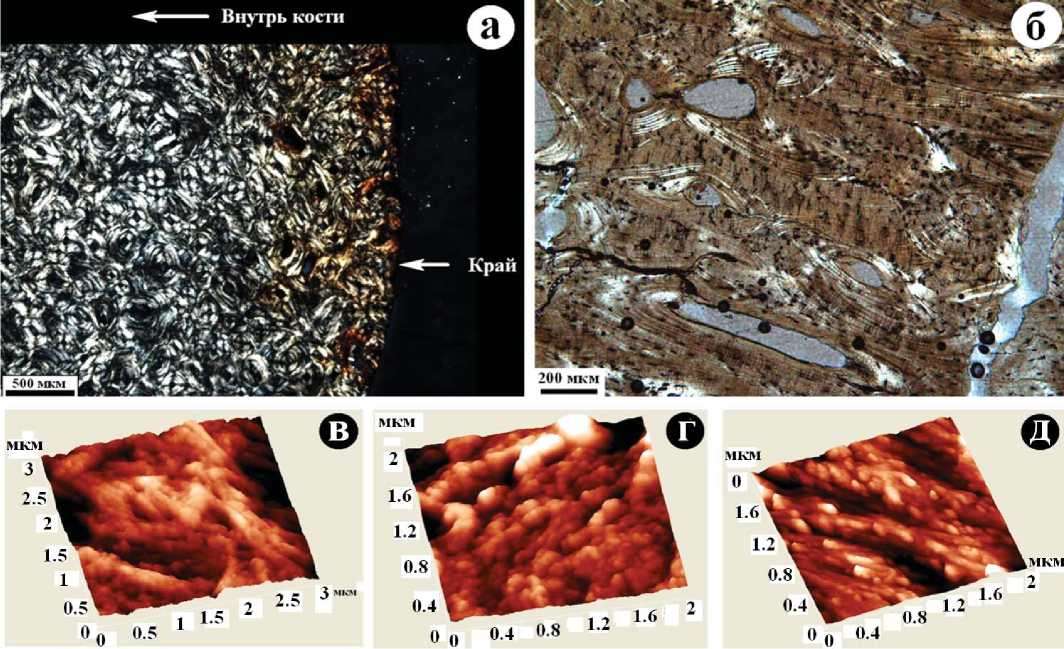
Рис. 3. Вариации микронаностроения костного детрита, выявляемого методами оптической (а, б) и зондовой атомно-силовой (в—д) микроскопии: а, д — усть-ишимская кость, б — костный детрит из святилища Усть-Полуй, в — кость современного человека, г — костные останки тоболо-иртышского тюрка
Fig. 3. Variations of the microstructure of the bone detritus detectable by optical (a, б) and atomic force probe (в-д) microscopy: a, д — Ust-Ishim bone, б — bone detritus from the sanctuary Ust-Polui, в — bone of modern man, г — bones of Tobol-Irtysh Turk

Рис. 4. Микростроение усть-ишимской кости, выявляющееся в СЭМ в режиме упруго-отраженных электронов: а — общий вид; б—г — костный биоапатит (1) с участками ультрадисперсного оксигидроксидного ожелезнения (1а); д, е — эпигенетическая минерализация пирита (2) и гётита (3) в крупных порах кости
Fig. 4. Microstructure of Ust-Ishim bone in the scanning electron microscope in elastically backscattered electrons mode: a — general view; б-г — bone bioapatite (1) with areas of ultradisperse oxyhydroxide ferrugination (1a); д, e — epigenetic mineralization of pyrite (2) and goethite (3) in large pores of the bone
Нанопористость
Важнейшим первичным генетическим свойством органоминерального костного композита является пористость в нанометровом диапазоне размеров (нанопористость), тесно связанная со структурой костного органоминерального композита. В настоящее время известно, что поры нанометрового размера в костях располагаются в стыках кристаллитов биоапатита и на контактах кристаллитов с волокнами коллагена. Размер этих пор колеблется от первых единиц до первых десятков нанометров. В ходе анализа определяются значения удельной общей поверхности пор, совокупного объема пор, среднего их размера, общего объема и среднего размера мезонанометровых (550 нм) и микронанометровых (< 5 нм) пор по отдельности.
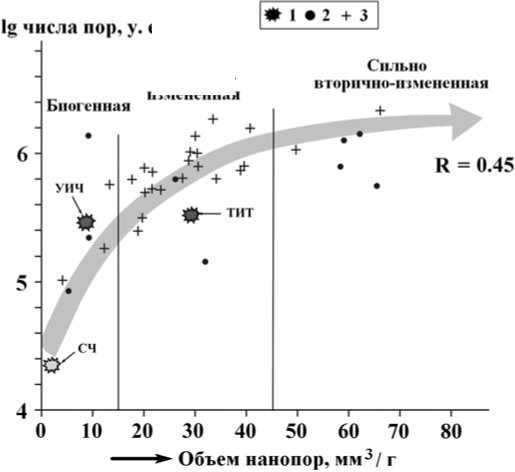
На рис. 5 показаны результаты, полученные в ходе исследований костного детрита неоплейстоценовых животных и костных останков людей. Из этих результатов следует, что общий объем нанопор и их число прямо и довольно сильно коррелируются между собой, согласованно возрастая по мере увеличения степени фоссилизации костей. Последняя при этом, очевидно, обусловлена не только возрастом костей, но и условиями захоронения. Обращает на себя внимание расположение на диаграмме точек человеческих костей относительно генерального тренда изменчивости нанопористости. Наименьшей пористостью, вполне ожидаемо, характеризуется кость современного человека. Кость средневекового тоболо-иртышского тюрка отличается многократно большей
Bi торпчио- е,
и{Меченная
Рис. 5. Изменение нанопористости ископаемых костей в зависимости от степени их фоссилизации: 1 — костный детрит людей (СЧ — современный человек, ТИТ — тоболо-иртышский тюрок, УИЧ — усть-ишимский человек; 2, 3 — костный детрит неоплейстоценовой мамонтовой фауны из Печорского Приуралья (2) и Среднего Прииртышья (3). Заливкой показан генеральный тренд согласованного увеличения общего объема и числа нанопор по мере увеличения возраста костного детрита
Fig. 5. Change of nanoporosity of fossil bones, depending on their degree of fossilization: 1 — bone detritus of human (СЧ — modern man, ТИТ — Tobol-Irtysh Turk, УИЧ — Ust-Ishim man;
-
2, 3 — bone detritus of Pleistocene mammoth fauna from Pechora Suburals (2) and Middle Irtysh (3). Fill shows general trend of the agreed increase in the total volume and number of nanopores with increasing age of the bone detritus
пористостью, сопоставимой с пористостью костей неоплейстоценовых животных. А кость палеолитического усть-ишимского человека, напротив, обнаруживает аномально низкую пористость, что, возможно, отражает уникальность условий его захоронения.
Химический состав
Валовый химический состав ископаемых костей колеблется в широких пределах, включая не только компоненты состава биоапатита, но и множество других компонентов, появление которых, очевидно, обусловлено фос-силизацией. Примером этого могут служить данные о составе костного детрита плейстоценовых млекопитающих Печорского Приуралья (мас. %): SiO2 — 0.33—18.42; Al2O3 — 0.23-5.85; Fe2O3 — 0.26-16.41; MnO — 0-4.24; MgO — 0.77-2.08; CaO — 33.48-60.42; SrO — 0-0.20; Na2O — 0-1.08; K2O — 0-1.04; P2O5 — 24.15-36.66; SO3 — 0-0.38. В более экстремальных случаях содержание эпигенетических примесей в костном детрите может оказаться гораздо большим. Примером этого может служить таранная кость человека возрастом около 10 тыс. лет [9] с валовым составом (мас. %): SiO2 — 36.48; TiO2 — 0.35; Al2O3 — 6.80; Fe2O3 — 5.12; MnO — 0.61; MgO — 1.20; CaO — 28.29; Na2O — 0.52; K2O — 1.03; P2O5 — 18.69; SO3 — 0.91.
Корреляционный анализ показывает, что химические компоненты костей подразделяются на две конкурирующие группы: 1) CaO+Na2O+P2O5+SO3 — компоненты состава биоапатита; 2) SiO2+Al2O3+Fe2O3+MnO+ MgO+SrO+K2O — компоненты состава терригенных загрязнений, обусловленных средой захоронения (результат иллювиирования). Примесь такого рода в костях может варьироваться от нескольких до 50 мас. %. Расчеты показали, что основными нормативными минералами терригенных загрязнений являются кварц, полевые шпаты и глинистые минералы (хлориты, гидрослюды). Кроме того, может вычисляться нормативная примесь пирита и гётита, которая отражает факт эпигенетической минерализации костей. На соответствующей диаграмме (рис. 6), построенной по данным исследования костного детрита мамонтовой фауны Печорского Приуралья, видно, что лишь часть проанализированных образцов располагается в полях состава кварцевых речных осадков. Большинство же точек широко рассосредоточилось, что может говорить не только о разном составе грунтов, из которых отбирали кости, но и о неоднократном переотложении костей в различные по составу грунты.
Микроэлементы
Содержание микроэлементов в костном детрите является важнейшим источником не только первичной генетической информации, но, еще в большей степени, информации об условиях, геохимической направленности и интенсивности процесса фоссилизации. В составе исследуемого нами костного детрита было выявлено около 50 микроэлементов, включая 14 лантаноидов. Общее содержание элементов варьируется в пределах от 500 в зубной эмали мамонтов до 2500 г/т в костных останках. Нормирование на средние содержания микроэлементов в наземных организмах (по В. В. Ковальскому) обнаруживает сильное обогащение ископаемых костей 36 микроэлементами из 49, причем степень такого обогащения изменяется в диапазоне пяти порядков (от 10 до 105 раз). К первой группе относятся микроэлементы, характеризующиеся увеличением относительных концентра- 23
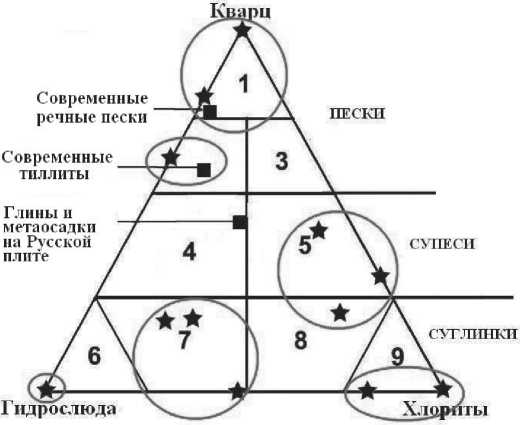
Рис. 6. Треугольник нормативно-минерального состава иллюви-ированной примеси в ископаемых костях в сравнении с составом терригенных осадков. Поля состава осадков: 1-3 — пески соответственно кварцевые, хлорит-гидрослюдисто-кварцевые и гидрослюдисто-хлорит-кварцевые; 4, 5 — супеси соответственно хлорит-кварц-гидрослюдистые и гидрослюдисто-кварц-хло-ритовые; 6-9 — суглинки и глины соответственно кварц-гидрослюдистые, кварц-хлорит-гидрослюдистые, кварц-гидро-слюдисто-хлоритовые, кварц-хлоритовые. Черные квадраты — средние составы некоторых терригенных формаций. Звездочками показаны исследованные образцы костного детрита неоплейстоценовых млекопитающих Печорского Приуралья.
Fig. 6. Triangle of normative mineral composition of illuvial impurity in fossil bones in comparison with the composition of clastic sediments. Fields of composition of sediments: 1—3 — sand, respectively quartz, chlorite- hydromica-quartz and hydromicachlorite-quartz; 4, 5 — sandy loams, respectively chlorite-quartz-hydromica and hydromica-quartz-chlorite; 6—9 — loams and clays, respectively quartz-hydromica, quartz-chlorite-hydromica, quartzhydromica- chlorite, quartz-chlorite. Black squares — average compositions of some clastic formations. Asterisks show the studied samples of bone detritus of Pleistocene mammals from Pechora Suburals ций в костных остатках в десятки раз (* — эссенциальные элементы, ** — элементы ксенобиотные): V*, Mo*, Ag**, Cs**, Sr**, Hf**, As**. Вторую группу образуют микроэлементы, концентрация которых возрастает в сотни-тысячи раз: Zn*, Ni*, Co*, Cr*, V*, S*, Li**, Ba**, Ga**, Ge**, Zr**, U**, Ln**, Sb**. В третью группу входят Y**, Ln**, Sc** Nb**, относительная концентрация которых может подскочить в десятки и даже сотни тысяч раз. Из приведенных групп следует, что возрастание степени обогащения ископаемых костей происходит в направлении от щелочных и щелочноземельных элементов-литофилов к переходным элементам-халькосидерофилам, а затем к многозарядным в ионной форме элементам-гидролизатам, включая иттрий и лантаноиды. Такой тренд от легко подвижных в экзогенных обстановках элементов к инертным, легко осаждаемым на большинстве геохимических барьеров, только подчеркивает эпигенетическую природу обогащения микроэлементами костного детрита.
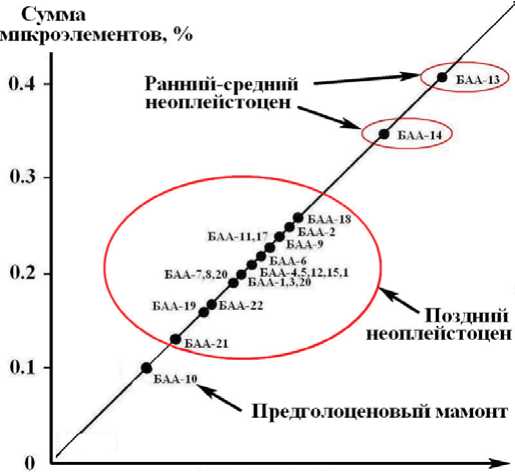
Если исходить из выше приведенного соображения, то можно построить график корреляции содержания микроэлементов, особенно ксенобиотных, со степенью фоссилизации ископаемых костей (рис. 7). Как оказалось, расположение точек на этом графике неплохо согласовалось с относительным возрастом костных 24
Продолжительность фоссчшпзашш
Рис. 7. Тренд валовой концентрации микроэлементов в ископаемых костях плейстоценовых млекопитающих Печорского Приуралья как отражение зависимости микроэлементного состава от степени их фоссилизации
Fig. 7. Trend of gross concentration of microelements in the fossil bones of Pleistocene mammals from Pechora Suburals as a reflection of microelement composition depending on their degree of fossilization образцов, что подтверждает возможность использования концентрации микроэлементов в ископаемых костях как критерия степени фоссилизации.
Фазовый состав биоминеральной компоненты
Согласно рентгеноструктурным данным, биоапа-титовая фаза в костном детрите широко варьируется по степени кристалличности. На дифрактограммах костей в области 30-35° углов 2 0 , где реализуются два основных отражения от (211) и (300) в структуре апатита, наблюдается одиночный, но сильно уширенный и почти нерас-щепленный пик (рис. 8, а). В случае дентина или бивней основной пик становится более интенсивным, а число и четкость дополнительных отражений увеличиваются (рис. 8, б). На рентгенограммах зубной эмали разрешается практически вся система основных и дополнительных отражений, характерных для хорошо окристаллизованно-го апатита (рис. 8, в). Соответственно, костный биоапатит характеризуется широким варьированием по величинам параметров элементарной ячейки (нм): ао = 0.932-0.948; со = 0.682-0.691. Полученные значения могут быть вполне определенно интерпретированы как параметры элементарной ячейки карбонатапатита В-типа. Есть также основания предполагать, что по мере фоссилизации эти параметры изменяются не совсем хаотично, указывая на некоторое возрастание карбонатности биоапатита.
В спектрах ИК-поглощения наблюдается весь набор линий валентных и деформационных колебаний PO4-тетраэдров и замещающих эти тетраэдры СО3-групп (рис. 8, г). Кроме того, в спектрах проявилась довольно интенсивная линия валентных колебаний Н2О. В целом полученные ИК-спектры вполне сопоставимы со спектрами, приведенными в литературе для ископаемого
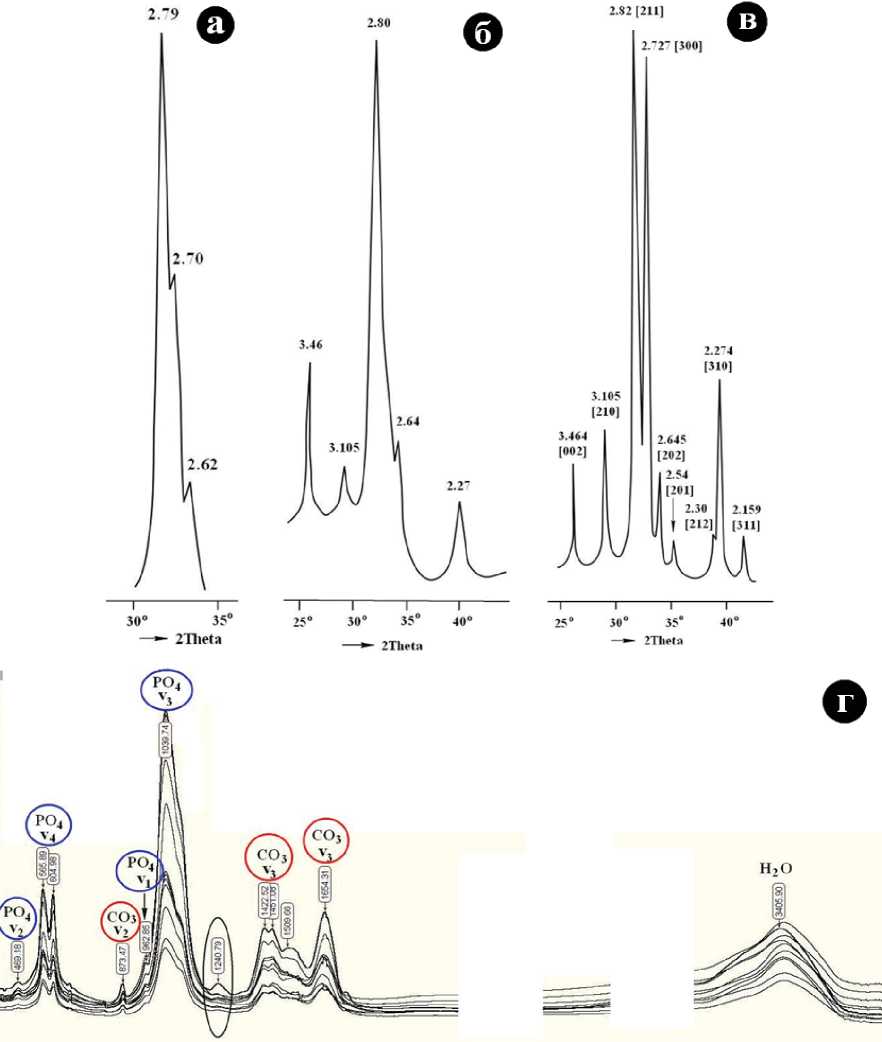
Рис. 8. Рентгеновские дифрактограммы с наиболее сильными отражениями, полученные от костного вещества мамонтов (а-в), приведенные в последовательности повышения степени кристалличности карбонатапатита (а — череп, б — бивень, в — эмаль коренного зуба) и типичные спектры ИК-поглощения в образцах костного детрита плейстоценовых млекопитающих (г)
Fig. 8. X-ray diffraction patterns with the strongest reflections received from mammoth bone substance (a-в) listed in the sequence of increasing degree of crystallinity of carbonate-apatite (a — skull, б — tusk, в — molar enamel) and typical IR absorption spectra of bone detritus samples of Pleistocene mammals (г)
костного детрита кайнозойских животных [2, 25]. Анализ показал, что интенсивности линий РО4- и СО3-групп обратно коррелируются (r = —0.45), что как раз и отражает изоморфные замещения по схеме СО3 ^ РО4.
Сопоставление результатов рентгеновских и ИК-спектроскопических исследований указывает на обратную корреляцию степени окристаллизованности биоапатита с содержанием в нем структурной примеси СО3. При этом выявляется устойчивый тренд возрастания кристалличности и снижения карбонатности биоапатита в направлении от костей к зубному дентину и зубной эмали. Показателем степени карбонатности биоапатита является так называемый апатитовый модуль — атомная пропорция между кальцием и фосфором, значение которой в стехиометричном апатите составляет 1.67. В нашем случае значение апатитового модуля колеблется в пределах от 1.62 до 2.3, перекрывая весь диапазон варьирования состава карбонатапатита как природного, так и искусственного происхождения (рис. 9).
В результате рентгенофазового анализа в составе костного детрита в качестве ксеноминеральных примесей установлены кварц, полевые шпаты, гидрослюды, хлорит, барит, кальцит, доломит, гётит, олим-пит Na3[PO4], холтедалит Mg2[PO4](OH)6, пирофосфат меди Cu2P2O7 x 5H2O, монофосфат калия KH2[PO4]. Происхождение этих примесей различно. Присутствие
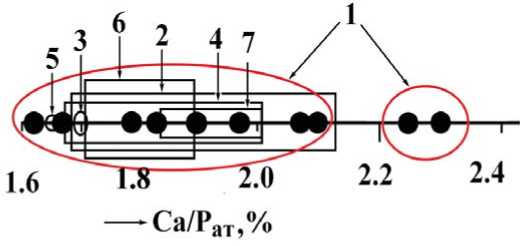
Рис. 9. Вариации апатитового модуля в карбонатапатите В-типа различного происхождения: 1 — исследуемый костный детрит плейстоценовых млекопитающих; 2 — костные остатки голоценовых млекопитающих [17]; 3 — конодонты [11]; 4 — мезозойские копролиты [18]; 5 — кость и зубная эмаль человека [11]; 6 — гипергенные ф
Fig. 9. Variations of apatite module in carbonate-apatite of B-type of different origin: 1 — analyzed bone detritus of Pleistocene mammals;
2 — Holocene fossil remains of mammals [17]; 3 — Conodonts [11]; 4 — Mesozoic coprolites [18]; 5 — bone and tooth enamel of human [11]; 6 — hypergene phosphorites [12]; 7 — synthetic analogs [1]
кварца и алюмисиликатов можно объяснить иллювииро-ванием в кости терригенного материала из вмещающих грунтов. Барит, карбонаты и фосфаты — результат топохимических изменений костного вещества, обусловленных воздействием на детрит грунтовых вод. В случаях захоронения костных останков животных в известняковых пещерах происходит замещение биопапатита на 40—60 % карбонатами.
Костный коллаген
Важнейшим источником генетической информации в ископаемых костях выступает костный белок — коллаген. В настоящее время у палеонтологов практически единственным методом оценки содержания и степени деградации органического матрикса в костном детрите является термический анализ, в ходе которого костный белок в интервале 250—500 °C выгорает с образованием на кривой нагревания одного или двух экзотермических экстремумов [17]. В нашем случае это осуществлялось не только термическим методом, но и непосредственным определением в костях валового содержания Сорг. Проведенный анализ показал, что в ископаемом костном детрите палеоплейстоцен-голоценового возраста содержание Сорг снижается от 15 до 3—4 мас. %, обратно коррелируясь с возрастом костей. Учитывая, что в «живых» костях содержание Сорг составляет 25—30 мас. %, получаем замечательный критерий относительного возраста и степени фоссилизации ископаемого костного детрита, основанный на факте непрерывности деградации органического матрикса в захороненных костях.
Высококвалифицированным химиком О. В. Кокшаровой было осуществлено выделение костного коллагена из исследованных нами образцов методом осторожного химического растворения биоминеральной компоненты. В результате обнаружилось, что коллаген имеет спутанно-волокнистое строение и широко варьируется по окраске от желтовато-оранжевого в костях современных людей до буровато-черного и черного в костях нео-плейстоценовых и более древних животных (рис. 10). Это обусловлено, как известно [3], постепенным окислением, молекулярным разложением и вторичной полимери- 26
зацией костных белков при фоссилизации. По данным элементного анализа коллаген в ископаемых костях по сравнению с таковым в костях современных позвоночных содержит меньше углерода и водорода, но больше азота и кислорода. Таким образом, органическое вещество в костном детрите действительно обнаруживает признаки сильного химического преобразования.
В органическом матриксе ископаемых костей обнаруживаются 13—15 аминокислот, общее содержание которых на 15—20 % уступает их содержанию в кости современного человека. Для некоторых аминокислот установлено присутствие рацематов — смесей левых (L) и правых (D) энантиомеров. Появление последних прямо свидетельствует о деградации коллагена, степень которого отражают именно D/L-отношения. Так, для усть-ишимской кости значение этого отношения в алла-нине составило 0.06, а в аспариновой кислоте — 0.03. Аналогичные оценки для костного детрита неоплейсто-ценовой мамонтовой фауны Прииртышского района в 5 раз выше.
В ИК-спектрах поглощения в исследуемом коллагене в диапазоне 1230—3400 см-1 проявилось множество линий, отвечающих всем основным химическим связям и функциональным группам в белковых молекулах. Большая часть зарегистрированных линий обусловлена колебаниями пептидных связей -СО-NH-, —NH, NH2- и CN в первичных и во вторичных амидах. Линии в области 960—930 см-1 соответствуют валентным колебаниям связей N-О в группировке C=N-OH, а линии в области 900-600 см-1 можно объяснить внеплоскостны-ми веерными колебаниями групп NH. ИК-поглощение, обусловленное алифатическими группами, проявляется в областях 2990-2970 и 2900-2870 см-1, а также при 1440 см-1. Оно вызвано валентными и деформационными колебаниями групп соответственно СН2 и СН3. Линия при 725-760 см-1, наблюдающаяся только в спектрах наиболее измененных костей, отвечает колебаниям групп СН2 в открытых цепях (СН2)4. Карбонильные группы диагностируются по линии поглощения при 1700 см-1, отвечающей колебаниям связей С=О, а также по линиям асимметричных валентных колебаний связей С-О в простых эфирах в области 1140-1160 см-1.
Из полученных данных следует, что ископаемые кости даже близкого возраста, но разного географического положения могут существенно различаться по степени деградации в них органического матрикса, образуя, тем не менее, единый тренд сокращения роли алифатических и карбонильных групп при возрастании роли пептидных связей в амидных группах (рис. 11). Такое изменение ИК-поглощения считается отражением генеральной тенденции деградации первичного органического вещества по мере фоссилизации костей [3].
Нами, вероятно впервые, в коллагене ископаемых костей обнаружено множество минералов в основном эпигенетического происхождения, но некоторые из них могут оказаться и первичными, т. е. биогенными. Общий список этих минералов, диагностированных рентгеноструктурным и рентгеноспектральными микрозондовым методами, включает кварц, альбит, ортоклаз, роговую обманку, клиноцоизит, диоктаэдрическую гидрослюду, циркон, ильменит, барит, карбонаты (кальцит, доломит), монацит (куларит), пирит, галогенсульфиды меди, магнетит, оксигидроксиды марганца, поликомпонентные самородно-металлические фазы.

Рис. 10. Изменение цвета и текстуры костного коллагена в зависимости от возраста костей: а — современный человек; б, в — из средневековых погребений, Верхне-Енисейский район; г — из погребений бронзового века, Северное Приангарье; д — из неолитовых погребений, Верхне-Енисейский район; е — усть-ишимский человек, палеолит
Fig. 10. Change of the color and texture of the bone collagen depending on the age of the bones: a — a modern man; б, в — medieval burials, Upper Yenisei region; г — burials of the Bronze Age, North Angara; д — Neolithic burials, Upper Yenisei region; e — Ust-Ishim man, Palaeolithic
Наибольший интерес из этих минералов представляют собой микровключения магнетита и клиноцоизита, обнаруженные в коллагене усть-ишимской кости (рис. 12), а также металлические фазы, найденные в колллагене плейстоценовых млекопитающих Среднего Прииртышья. Фазы характеризуются не совсем обычной для абиогенных самородно-металлических минералов композицией, подразделяясь на несколько групп: 1) самородное желе зо Fe 0.90-0.99 Ni 0-0.08 Cu 0-0.05 Mn 0-0.0J 2) ЖеЛе30 ™K№ стое Fe 0.57-0.88 Ni 0.10-0.46 Cu 0-0.02 Zn 0-0.06 и медистое Fe 0.67-0.86 Cu 0.04-0.30 Ni 0-0.03 Zn 0-0.06 i 3) ™ке Л ь и Н ике Л ь жеёе- ЗИСТЫЙ Ni 0.50-0.97 Fe 0.03-0.48 Cu 0-0.02 Zn 0-0.01 Cr 0-0.01 V 0-0.01 ; 4) ме д ь Ц и НК истая ( лагун ь) Cu 0.50-0.66 Zn 0.32-0.47 Fe 0-0.04 Ni 0-0.04 Pb 0-0.01 , Н ике Л истая (ме Л ьхио р) Cu 0.85 N i 0.14 Fe 0.01 и же Л езистая Cu 0.48-0.57 Fe 0.40-0.47 Ni 0.03-0.04 Cr 0-0.01 ; 5) само р о дн ый Ц и Н к Zn 0.93_0.96 Fe 0-0.05 Cu 0-0.04 Ni 0-0.02 , Ц™ к Же-ёезо-ме д истый Zn 0.70-0.72 Cu 0.16-0.18 Fe 0.04-0.12 Co 0-0.08 , Ц™к никелисто-железистый Zn0 54Fe0 22Ni y 16Cu0 05Co0 03 и цинк х р омистый Zn 0. 55 Cr 0.30 Fe 0.05 V 0.01 "
Изотопия
Важнейшим источником палеонтологической и палеоэкологической информации в последние 15-20 лет становятся изотопно-геохимические данные, полученные как по биоминеральному, так и органическому веществу костей [5, 6, 19-26]. Идейным основанием для таких исследований служит, во-первых, обусловленность изотопного состава углерода и кислорода в костном биоапатите бикарбонат-ионами, растворенными в воде, потребляемой животными, а во-вторых, зависимостью изотопного состава углерода и азота в органическом матриксе костей от пищи, поглощаемой животными в течение своей жизни. Следует отметить, что несмотря на открытость вопросов о степени изотопной одно родности упомянутых выше элементов в организмах и степени устойчивости изотопных отношений в процессах фоссилизации, именно реконструкция палеодиет по коллагену оказалась в настоящее время самым востребованным направлением изотопных исследований в палеонтологии.
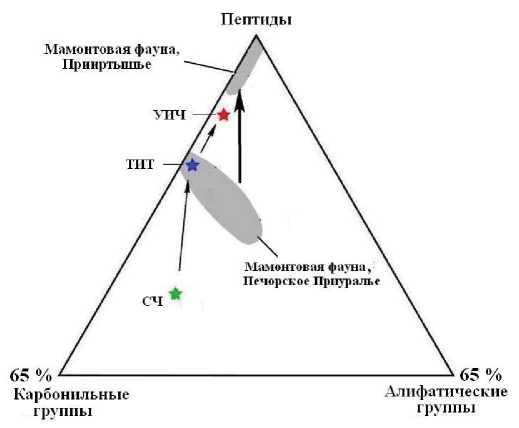
Рис. 11. Генеральная тенденция изменения химикомолекулярной структуры органического вещества в костном детрите неоплейстоценовых млекопитающих. Звездочки — состав коллагена, выделенного из костных останков людей: СЧ — современный человек, ТИТ — средневековый тоболо-иртышский тюрок, УИЧ — усть-ишимский человек
Fig. 11. General trend of change in the chemical and molecular structure of organic matter in the bone detritus of Neopleistocene mammals. Stars - composition of collagen from the skeletal remains of human: СЧ - modern man, ТИТ - medieval Tobol-Irtysh Turk, УИЧ - Ust-Ishim man
Проведенные нами исследования показали, что изотопный состав углерода и кислорода в биоапатите ископаемых костей плейстоценовых животных оказался довольно однородным, неплохо согласуясь с таковым в пластовых водах, в карбонатных раковинах раннеголоценовых пресноводных улиток, а также в карбонатапати-те из гипергенных фосфоритов кайнозойского возраста (рис. 13, а). Кроме того, полученные нами данные оказались близкими к аналогичным данным для мамонтов Западной Европы. На этом фоне обращает на себя внимание явное отклонение от полей мамонтовой фауны точек изотопного состава карбонатных С и О в биоапатите человеческих костей. Последние отличаются изотопно более легким углеродом и, напротив, изотопно более тяжелым кислородом. Объяснение этого мы видим в том, что, как центрально-европейская, так и евразийская неоплей-стоценовая мамонтовая фауна в отличие от усть-ишимско-го человека, тоболо-иртышского тюрка и, тем более, современных людей потребляла опресненную постледниковую воду, обедненную бактериальной органикой.
Полученные изотопные данные по коллагену свидетельствуют о широком разбросе значений изотопных коэффициентов (%с): 5 13С = —25...—17, 5 15N = —0.5^14. Тем не менее, полученная картина изотопной неоднородности (рис. 13, б) представляется закономерной. Так, поле исследованных нами в Печорском Приуралье неоплей-стоценовых северных оленей приходится на промежуток между стандартными полями для северных оленей и лосей. Это можно расценивать как указание на стабильную для этих животных смешанную лишайниково-кустарниковую диету. Поле печоро-приуральского шерстистого носорога сдвинуто относительно области, обычно характерной для нежвачных травоядных, в область более изотопно-легких углерода и азота. Это вероятно можно объяснить, во-первых, обитанием исследованных нами животных в зоне перехода от редколесья к лесам, а, во-вторых, развитием соответствующей кормовой базы на кислых почвах.
Весьма необычным является положение изотопного поля печоро-приуральского мамонта, который, как известно, считаются «изотопно-уникальным» животным из-за изотопно-легкого (липидогенного) углерода и аномально изотопно-тяжелого азота в коллагене костей [19]. Причину этого видят в особенностях пищеварения [21] и копрофагии [22, 26]. Однако в нашем случае поле изо топного состава сильно сдвинулось в область изотопнолёгкого азота, наложившись на аналогичные поля для шерстистых носорогов, лошадей и быков. Из этого следует, что в Печорском Приуралье мамонты в отличие от западно-европейских животных были лучше обеспечены качественной растительной пищей.
Но поистине сенсационно выглядят данные, полученные нами для пещерных медведей. Согласно этим данным, костный детрит пещерных медведей характеризуется умеренно изотопно-легким углеродом ( 5 13С = —20.8 ± 1.02 %) и изотопно-легким азотом ( 5 15N = 3.94 ± 0.62 %), обычно характерным для растениеядных животных. Таким образом, вопреки традиционному отнесению пещерных медведей к хищникам, исследованные нами экземпляры были вовсе не хищниками, а вполне себе травоядными. Недавно к такому же выводу пришли и финские палеонтологи (Жаклин Мустакас, университет Хельсинки), правда на основе косвенного критерия — строения зубов. Наши финские коллеги предположили, что именно неспособность пещерных медведей к смене растительной диеты на растительно-мясную или мясную и послужила главной причиной их вымирания в период последнего неоплейсто-ценового оледенения. Очень может быть, что именно так все и было. Например, костный коллаген переживших это оледенение неоплейстоценовых бурых медведей (западно-сибирская популяция) характеризуется примерно таким же углеродом ( 5 13С = —20.32 ± 0.42 %), но гораздо более изотопно-тяжелым азотом ( 5 15N = 8.98 ± 0.99 %). То есть поздне-неоплейстоценовые бурые медведи были уже хищниками. Проведенные нами изотопные исследования волос показал и что современные таежные бурые медведи — преимущественно растениеядные животные, но на Камчатке они практикуют смешанную диету, чередуя поедание изотопно-легких по азоту растений с поеданием изотопно-тяжелой по азоту заходящей в реки на нерест морской рыбы. Соответственно этому их белки характеризуются гораздо более изотопно-тяжелым азотом.
На фоне полей изотопного состава костного коллагена травоядных животных аналогичные поля людей на рис. 13 существенно сдвинуты в область более изотопнотяжелых значений для углерода и азота, что можно объяснить существенно мясной диетой, как неандертальцев, так и Homo Sapiens. При этом обнаруживается, что на-
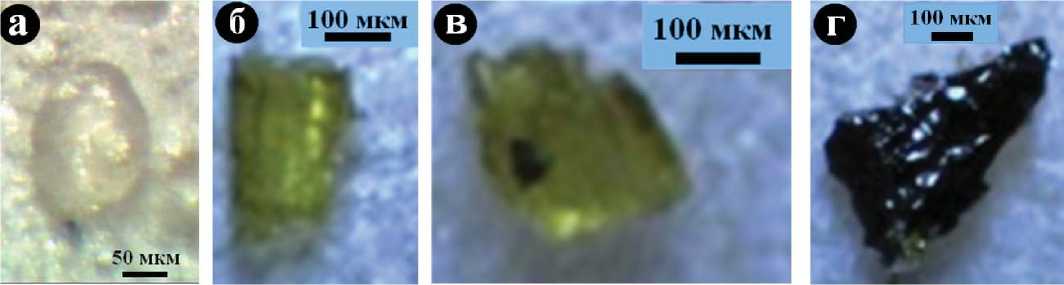
Рис. 12 . Минеральные примеси, обнаруженные в коллагене усть-ишимской кости: а — кварц; б, в — клиноцоизит состава (Ca 1 89-2.05Mg0-0.18)1.95-2.05(Al2-2.14Fe0.87-1)2.99-3.13[Si3 O 12](OH); г маГНеТИТ COCTaBa (Fe o^-i Mn O-O^X Fe l^^ Al O-O^X O ( миНаЛЫ, мол. %: маг нетит 97-98, герцинит 0-3, якобсит 0-2)
Fig. 12. Mineral impurities in collagen of Ust-Ishim bone: a — quartz, б; в — clinozoisite (Ca1 89-2 05Mg0-0 18)1 95-2 05(Al2-214Fe0 87-1) 2 99-3 13 [Si3O12](OH); г - magnetite (Fe0 98-1Мп у -0 02)(Fe195-2Al0-0 05)2O4 (minals, mol %:. magnetite 97-98, hercynite 0-3, jacobsite 0-2)
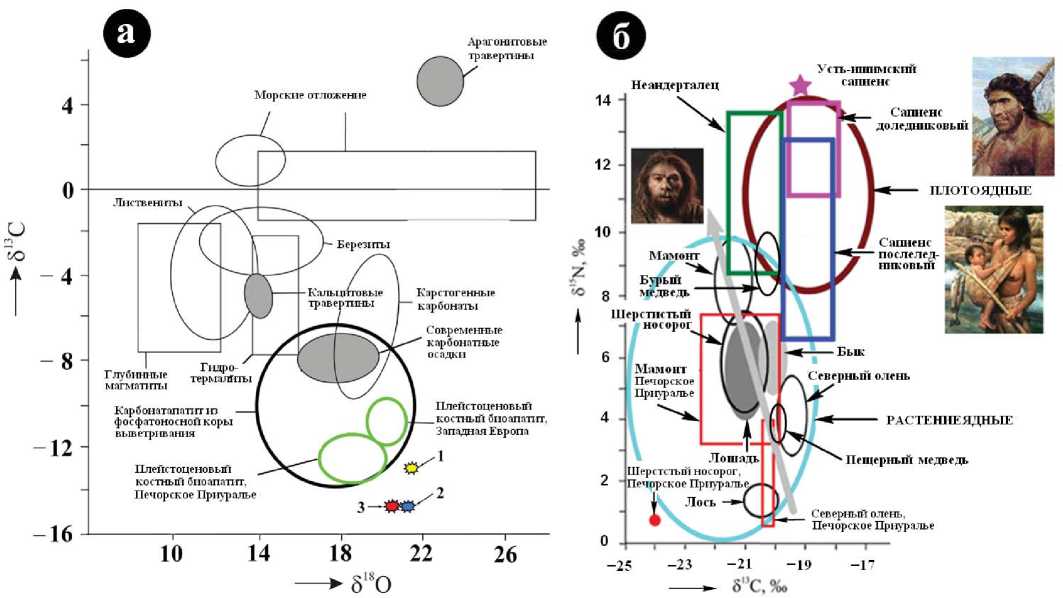
Рис. 13. Вариации изотопного состава углерода, кислорода, азота в биоапатите (а) и коллагене (б, частично использованы данные из [19]) ископаемого костного детрита
Fig. 13. Variations of isotopic composition of carbon, oxygen, nitrogen in bioapatite (a) and collagen (б data was partially used from [19]) of fossil bone detritus
ибольшими мясоедами были доледниковые неоплейсто-ценовые люди (усть-ишимский человек), а вот в диете послеледниковых предголоценовых и раннеголоценовых Homo Sapiens доля мяса была даже ниже, чем у неандертальцев. Очевидно, что это обусловлено предголоцено-вой биотической катастрофой — массовым вымиранием мамонтовой фауны, на которую привыкли охотиться люди палеолита.
Заключение
Не смотря на некоторые прогрессивные декларации [10], палеонтология в практическом смысле остается наукой, в которой основные выводы делаются на основе так называемых органолептических критериев. Это отсылает ее даже не в прошлый, а в девятнадцатый век, когда аналитические методы только зарождались и палеонтологи были вынуждены в основном ограничиваться тем, чем их самих биологически наградила природа — зрением, обонянием, осязанием и т. п. Однако в XXI веке так продолжаться не может. Только осознанное освоение и плодотворное комплексное использование множества прецизионных минералогических, физических, физикохимических и химических методов может обеспечить палеонтологии успешное будущее.
За помощь и сотрудничество в исследованиях авторы благодарны С. М. Слепченко, А. А. Бондареву, П. А. Косинцеву, Д. В. Киселёвой, Е. А. Васильеву, О. В. Мартиросян, В. Н. Филиппову, О. В. Кокшаровой, С. Т. Неверову.
Исследования выполнены при поддержке грантом Президента РФ «Для государственной поддержки ведущих научных школ Российской Федерации» НШ-9723.2016.5
Список литературы Современные исследования ископаемого костного детрита: палеонтология, минералогия, геохимия
- Брик А. Б., Франк-Каменецкая О. В., Дубок В. А. и др. Особенности изоморфных замещений в синтетических карбонатфторапатитах//Минералог. журн. 2013. № 3. С. 3-10.
- Вотяков С. Л., Киселёва Д. В., Щапова Ю. В., Смирнов Н. Г., Садыкова Н. О. Физико-химические характеристики ископаемых костных остатков млекопитающих и проблема оценки их относительного возраста. Ч. 2. ИК-и радиоспектроскопия, микроскопия. Екатеринбург: Изд-во «Гощицкий, 2009а. 82 с.
- Дроздова Т.В. Геохимия аминокислот. М.: Наука, 1977. 199 с.
- Майдль Т. В., Безносов П. А., Смолева И. В., Силаев В. И. Результаты минералого-геохимических исследований ихтиофоссилий и необычная микроминерализация из девонских карбонатных отложений на Южном Тимане//Современные проблемы теоретической, экспериментальной и прикладной минералогии (Юшкинские чтения-2016). Сыктывкар: Геопринт, 2016. С.152-153.
- Николаев В. И., Барбиери М., Даванзо С., Кузнецова Т. В., Лонжинелли А., Сулержицкий Л. Д., Якумин П. Комплексные изотопные исследования мамонтов Якутии//Квартер-2005: Материалы IV Всероссийского совещания по изучению четвертичного периода. Сыктывкар: Геопринт, 2005. С. 297-295.
