Создание исходного материала для маркер-опосредованной селекции родительских линий картофеля (Solanum tuberosum L.) на диплоидном уровне (обзор)
 на диплоидном уровне (обзор)")
Автор: Ермишин А.П., Воронкова Е.В.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Картофелеводство: наука и технологии
Статья в выпуске: 1 т.52, 2017 года.
Бесплатный доступ
Для повышения эффективности селекции картофеля важно, чтобы селекционные линии были мультиплексными (имеющими по два и более доминантных аллеля) по многим хозяйственно ценным генам (прежде всего, генам устойчивости к болезням и вредителям) (J.E. Bradshaw с cоавт., 1994). Мультиплексные родительские линии по нескольким генам устойчивости среди существующих сортов встречаются достаточно редко (А.П. Ермишин с соавт., 2016) и вследствие биологических особенностей культуры картофеля могут быть получены только при отборе на диплоидном уровне. В представляемом обзоре описаны генетические принципы и опыт их практического применения в исследованиях авторов по созданию исходного материала, позволяющего проводить эффективную маркер-опосредованную селекцию родительских линий на диплоидном уровне. Материал включает коллекцию первичных дигаплоидов сортов картофеля, отобранных по жизнеспособности, клубневой продуктивности, культурным признакам и наличию ДНК-маркеров генов устойчивости к болезням и вредителям; диплоидный селекционный материал с широким набором генов устойчивости к фитофторозу и вирусам, интрогрессированных от диких видов с помощью оригинальных методов преодоления межвидовых репродуктивных барьеров; диплоидные линии - доноры генов самосовместимости и высокой мужской фертильности, эффективных как в гетерозиготном, так и гомозиготном состоянии; диплоидные линии - доноры генов образования мужски фертильных FDR (first division restitution) 2 n -гамет. Лучшие диплоидные линии с комплексом генов устойчивости к болезням и вредителям, выделенные по результатам диплоидной селекции (на основе первичных дигаплоидов, межвидовых гибридов и доноров фертильности), используют для получения мультиплексных тетраплоидных родительских линий с помощью митотического удвоения хромосом. С использованием доноров генов образования фертильных FDR 2 n -гамет выводят диплоидные родительские линии, способные к гибридизации с сортами картофеля. Кроме того, созданный диплоидный исходный материал представляет интерес для перспективных альтернативных направлений селекции картофеля с применением отбора на диплоидном уровне: выведения диплоидных сортов картофеля, а также родительских линий для получения размножаемых ботаническими семенами гибридов и гибридных популяций.
Картофель, отбор на диплоидном уровне, маркер-опосредованная селекция (мос, мультиплексные родительские линии, мужская фертильность, нередуцированные гаметы
Короткий адрес: https://sciup.org/142213769
IDR: 142213769 | УДК: 635.21:631.527.7:571.21 | DOI: 10.15389/agrobiology.2017.1.50rus
Development of initial material for marker assisted potato (Solanum tuberosum L.) parental line breeding at the diploid level (review)
Текст обзорной статьи Создание исходного материала для маркер-опосредованной селекции родительских линий картофеля (Solanum tuberosum L.) на диплоидном уровне (обзор)
Для повышения эффективности селекции картофеля наиболее перспективной считается стратегия, основанная на получении гибридных популяций, имеющих высокую селекционную ценность, то есть тех, в которых генотипы, отвечающие требованиям, предъявляемым к сортам картофеля, представлены с более высокой частотой, чем при случайных скрещиваниях. Такие гибридные популяции получают, используя в качестве одного или обоих родителей предварительно изученные селекционные линии, обладающие высокой комбинационной способностью по показателям продуктивности и несущие комплекс генов устойчивости к болезням и вредителям. Важно, чтобы эти селекционные линии были мультиплексными (имеющими по два и более доминантных аллеля) по многим хозяйственно ценным генам (прежде всего, генам устойчивости к болезням и вредителям) (1).
Мультиплексные родительские линии по нескольким генам устойчивости среди существующих сортов встречаются достаточно редко (2). Вследствие биологических особенностей культуры картофеля (вегетативно размножаемый автотетраплоид) эффективная селекция родительских линий, мультиплексных по нескольким генам устойчивости, возможна только при условии применения отбора на диплоидном уровне.
На диплоидном уровне (благодаря более простому наследованию хозяйственно ценных признаков) значительно проще отбирать гибриды с комплексом желаемых признаков, чем на тетраплоидном уровне. Применение ДНК-маркеров позволяет существенно повысить эффективность диплоидной селекции. Генотипы, отобранные по комплексу маркеров генов устойчивости, а также показателям продуктивности, культурным признакам растений и клубней, могут быть переведены на тетраплоидный уровень с помощью митотического удвоения хромосом. В результате получают тетраплоидные родительские линии с комплексом генов, имеющимся у исходных дигаплоидов. При этом они представляют собой дуплексы по соответствующим генам, если дигаплоиды были гетерозиготами, и квадруплексы — в случае удвоения хромосом у гомозигот.
Перспективным считается создание диплоидных родительских линий, способных образовывать фертильную нередуцированную пыльцу, с помощью комбинативной селекции на диплоидном уровне. Их использование в скрещиваниях с тетраплоидными сортами дает возможность получать выравненные, продуктивные, экологически стабильные тетраплоид-ные гибридные популяции (3), практически не расщепляющиеся по комплексу признаков, сформированному на диплоидном уровне.
Для того чтобы вывести названные родительские линии, необходим исходный селекционный материал, позволяющий проводить эффективный отбор на диплоидном уровне. Он должен включать коллекцию дигаплоидов с широкой генетической основой, обладающих приемлемой фертильностью и несущих мутации, связанные с образованием нередуцированных гамет, что важно для выведения диплоидных родительских линий. Большинство селекционных компаний и генетические банки по картофелю таким материалом не располагают. В настоящей статье описаны генетические принципы, лежащие в основе создания исходного материала для диплоидной селекции картофеля, а также опыт их практического применения в исследованиях авторов.
Получение дигаплоидов картофеля. В генетике картофеля гаплоиды, полученные из тетраплоидов (2 n = 4½ = 48), обозначают как дигаплоиды (2 n = 2½ = 24), а гаплоиды от диплоидов (2 n = 2½ = 24) называют моноплоидами, моногаплоидами (2 n = ½ = 12). То есть термин «дигаплоид» не соответствует значению «удвоенный гаплоид», который используется в генетике диплоидных видов растений.
Основной метод получения дигаплоидов картофеля — опыление тетраплоидных сортов или селекционных клонов пыльцой гаплопродюсе-ров Solanum phureja Juz. & Bukasov. Цитологический механизм, с помощью которого возникают дигаплоидные зародыши, — псевдогамия и партеногенез (4). При опылении пыльцой S . phureja оба ядра ( n = ½ = 12) пыльцевого зерна сливаются с центральным ядром зародышевого мешка S . tuberosum (2 n = 4½ = 48), приводя к формированию гексаплоидного эндосперма. Ядро яйцеклетки ( n = 2½ = 24) остается неоплодотворенным, но стимулируется к дифференцировке. Имеются данные (5), что образование дигаплоидов может иметь и другой механизм и происходить за счет избирательной элиминации хромосом S . phureja в процессе формирования гибридных зародышей.
Наиболее широкое применение в селекции картофеля нашли голландские формы S. phureja IvP 35, IvP 48 и IvP 101, которые сочетают высокую гаплопродуцирующую способность с гомозиготным состоянием маркерного гена «эмбрионального пятна» (embryo-spot), контролирующего антоциановую окраску семядолей (6). Эмбриональное пятно легко детектируется при просмотре под бинокулярной лупой, поэтому с введением перечисленных гаплопродюсеров в селекционную практику появилась возможность выделять дигаплоиды картофеля (по отсутствию у них маркерного признака) с минимальными трудозатратами. Дигаплоиды, отобранные в потомстве от опыления гаплопродюсерами тетраплоидных сортов или селекционных клонов, называют первичными дигаплоидами, а потомство от их скрещиваний — вторичными дигаплоидами.
В лаборатории генетики картофеля Института генетики и цитологии НАН Беларуси исследования по отбору на диплоидном уровне в селекции картофеля ведутся с начала 1990-х годов на основе материала, предоставленного С.И. Лиореком (Украинский НИИ картофельного хозяйства, Киевская обл.). В этот период была создана большая коллекция диплоидного селекционного материала, которая включала первичные дигаплоиды картофеля, полученные в лаборатории, вторичные дигаплоиды на их основе, а также вторичные дигаплоиды, поступившие из других организаций. В связи с разработкой методов детекции ДНК-маркеров генов устойчивости к болезням и вредителям картофеля была поставлена задача улучшить качественный состав коллекции за счет получения новых первичных дигаплоидов, имеющих как можно большее разнообразие и высокую частоту маркеров генов устойчивости.
Было проведено изучение характеристик сортов картофеля различного происхождения, представленных в каталогах, в результате чего отобраны сорта, которые по описаниям могли иметь в геноме несколько генов устойчивости к болезням и вредителям. Наличие конкретных генов устойчивости подтверждали, используя соответствующие ДНК-маркеры. С помощью гаплопродюсера S . phureja IvP 35 получили около 100 первичных дигаплоидов ряда сортов и ценных селекционных клонов. При переходе на диплоидный уровень часть ДНК-маркеров, имеющихся у исходных сортов, утрачивалась в результате расщепления. Тем не менее, частота большинства анализируемых маркеров в полученной популяции первичных дигаплоидов оказалась достаточно высокой (7). Ежегодно коллекция пополняется новыми дигаплоидами сортов картофеля, предварительно оцененных по наличию ДНК-маркеров генов устойчивости к болезням и вредителям. Лучшие из дигаплоидов (при отборе, помимо наличия ДНК-маркеров, учитывается жизнеспособность растений, клубневая продуктивность, компактность гнезда, форма клубней, глубина глазков, способность к цветению) включают в программу скрещиваний с диплоидными линиями — донорами генов фертильности и формирования 2 n -пыльцы, а также донорами новых генов устойчивости к фитофторозу и вирусам, интрогрес-сированных нами в диплоидный селекционный материал от диких видов.
Диплоидный селекционный материал с генами устойчивости к фитофторозу и вирусам, инрогрессированными от диких видов. Одно из важных преимуществ отбора на диплоидном уровне по сравнению с тетраплоидным — более высокая эффективность вовлечения в селекцию ценного генофонда диких и примитивных культурных видов картофеля. Около 70 % описанных диких и культурных видов картофеля — ди-плоиды, многие из которых относительно хорошо скрещиваются с дигаплоидами S . tuberosum . Благодаря более простому наследованию на диплоидном уровне в беккроссных поколениях межвидовых гибридов быстрее и с меньшими затратами можно проводить отбор по отдельным генам, концентрировать желательные и избавляться от нежелательных генов диких видов (8).
Особенностью исследований нашей лаборатории в области отдаленной гибридизации картофеля было изучение механизмов межвидовых репродуктивных барьеров и разработка эффективных методов их преодоления с целью интрогрессии в селекционный материал ценных генов диких 52
видов, которые практически не скрещиваются с культурным картофелем. При этом среди достижений следует отметить разработку и реализацию подходов к преодолению презиготной несовместимости, основанных на использовании явления расщепления по S-генам родительских видов. С их помощью вовлечены в селекцию тетраплоидные соматические гибриды между дигаплоидами S . tuberosum и мексиканскими диплоидными дикими видами S . bulbocastanum и S . pinnatisectum (9, 10). Кроме того, было обосновано использование дикого диплоидного вида картофеля S . verrucosum и созданных в лаборатории на его основе SvSv -линий, у которых отсутствуют пестичные S-РНКазы, для устранения пре- и постзиготной несовместимости при межвидовых скрещиваниях. С помощью S . verrucosum и SvSv -линий удалось вовлечь в селекцию S . bulbocastanum , S . pinnatisectum , S . polyadenium , S . circaeifolium , S . commersonii , а также преодолеть одностороннюю несовместимость, характерную для аллотетраплоидных диких видов (11-14). Также нами обнаружено новое явление — образование диплоидных гибридов в скрещиваниях между аллотетраплоидными дикими видами картофеля S . acaule , S . stoloniferum , S . fendlery , S . polytrichon и дигаплоидами S . tuberosum . Получение таких гибридов дает возможность значительно упростить использование аллотетраплоидных видов в селекции ценного генофонда (15, 16). В результате применения перечисленных подходов получен уникальный диплоидный селекционный материал с комплексом генов высокой долговременной устойчивости к фитофторозу и вирусам (17, 18).
Создание доноров генов фертильности. Считается, что переход с тетраплоидного уровня на диплоидный при получении дигаплоидов картофеля соответствует приблизительно трем поколениям самоопыления (19). В результате большинство дигаплоидов имеют пониженную жизнеспособность, отстают в развитии по сравнению с исходными сортами. Одним из основных проявлений инбредной депрессии первичных дигаплоидов является их стерильность или пониженная фертильность.
Около половины получаемых дигаплоидов S . tuberosum характеризуются достаточно активным цветением, необходимым для их включения в гибридизацию. Однако использование цветущих генотипов возможно только в качестве материнских форм, так как в подавляющем большинстве эти растения мужски стерильны (20). Женская фертильность (способность завязывать ягоды и семена при опылении фертильной пыльцой совместимых опылителей), хотя и понижена у дигаплоидов S . tuberosum по сравнению с тетраплоидами, не является основным лимитирующим фактором в скрещиваниях с участием дигаплоидов (21).
Первичные дигаплоиды картофеля, формирующие жизнеспособную пыльцу (которая окрашивается, например, ацетокармином), в большинстве случаев не могут служить в скрещиваниях опылителями, так как их пыльца функционально неактивна. Фертильность, оцененная по результатам гибридизации, получила в литературе название «функциональная мужская фертильность» (ФМФ) (22). ФМФ определяется как абсолютным количеством образующейся пыльцы, так и долей в ней пыльцы, способной доставить мужские гаметы к зародышевому мешку (функционально фертильной). Для косвенной оценки функциональной фертильности пыльцы (ФФП) разработано несколько методов (23). По нашему опыту, наиболее приемлем метод, основанный на определении частоты проросших пыльцевых зерен на искусственной питательной среде за определенное время (24).
Степень проявления фертильности у дигаплоидов S . tuberosum определяется соответствующим показателем исходной тетраплоидной формы и эффектом инбридинга (8). У тетраплоидных сортов картофеля 53
практически не ведется отбор против рецессивных аллелей, неблагоприятных для мужской фертильности, так как в селекционных программах стерильные формы могут быть использованы в качестве материнских. При получении дигаплоидов они переходят в гомозиготное состояние, что проявляется в снижении фертильности и появлении стерильных форм. Гипотезу об инбридинге как основной причине мужской стерильности дигаплоидов у картофеля подтверждают оценки мужской фертильности удвоенных моноплоидов (25). Восстановлению фертильности дигаплоидов способствует их гибридизация с примитивными культурными и дикими диплоидными видами картофеля, которая устраняет эффект инбредной депрессии (8). Однако для выведения родительских линий, пригодных для использования в селекции сортов картофеля, необходимо несколько циклов скрещиваний на диплоидном уровне. Показано, что при гибридизации вторичных дигаплоидов картофеля, особенно в комбинациях с участием стерильных и низко фертильных форм, преобладают генотипы с пониженной функциональной фертильностью пыльцы (26).
Во многих случаях при гибридизации дигаплоидов S . tuberosum с диплоидными видами картофеля получают мужски стерильное потомство. Причина этого — генно-цитоплазматическая мужская стерильность, которая обусловлена взаимодействием доминантных ядерных генов, присутствующих у большинства южноамериканских диплоидных видов, и цитоплазматических генов S . tuberosum (27). Фактором, определяющим мужскую стерильность первичных дигаплоидов, может также быть их происхождение от сортов картофеля, несущих определенные типы цитоплазмы диких видов. Так, наличие у сортов цитоплазмы D (от S . demissum ) ассоциировано с функциональной стерильностью пыльцы, а цитоплазмы W/γ (например, от S . stoloniferum ) — с образованием полностью абортивной пыльцы (28).
При гибридизации дигаплоидов картофеля возможны неудачи из-за самонесовместимости, которая может быть существенным препятствием в реализации селекционных программ, связанных с использованием самоопыления, близкородственных скрещиваний, беккроссирования. Для диплоидных видов Solanum характерен гаметофитный тип самонесовмести-мости, которая проявляется в ингибировании и остановке роста пыльцевых трубок в верхней трети столбика пестика (11).
Изучение аллельного разнообразия S-генов у картофеля показало, что оно весьма ограниченно (29). Данные, полученные в нашей лаборатории, свидетельствуют, что результаты гибридизации ряда дигаплоидов S . tuberosum можно объяснить исходя из наличия у них не более четырех S-аллелей. При этом у дигаплоидов, происходящих от двух неродственных тетраплоидных сортов картофеля, может оказаться до трех общих S-аллелей (30). Решению проблемы может способствовать использование в скрещиваниях диплоидных селекционных линий с мутациями самосовместимости. Мутации самосовместимости обнаружены у дигаплоидов S . tuberosum (31), а также у некоторых диплоидных видов картофеля ( S . gоniocalix , S . kurzian-um , S . neohawkesii , S . phureja , S . pinnatisectum , S . raphanifolium , S . sanctae - rosae , S . stenotomum ) (32). Наиболее изучены в генетическом плане мутации tS1 у дигаплоидов S . tuberosum (31), а также Sli у S . chacoense (33). Гомозиготные доноры Sli были успешно использованы для селекции диплоидных инбредных линий, предназначенных в перспективе для получения на их основе гетерозисных гибридов (34).
В нашей лаборатории для решения проблемы пониженной фертильности диплоидного картофеля предложена программа создания доноров фертильности, которая включала комбинацию инбридинга, имеющего 54
целью элиминацию аллелей, неблагоприятных для фертильности, и гибридизации между высокофертильными генотипами с целью замещения неблагоприятных аллелей аллелями, которые положительно влияют на процесс развития пыльцы у картофеля (35).
В результате изучения большой коллекции вторичных дигаплоидов картофеля выделен клон IGC 203/5.7 (на основе дигаплоида сорта Полесский розовый и гибрида S . phureja ½ S . vernei ), обладающий мужской фертильностью и самосовместимостью (генетический анализ показал простое моногенное наследование этой мутации самосовместимости). Клон IGC 203/5.7 был включен в гибридизацию с высокофертильными формами IGC 92/1.1 и IGC 92/1.2, отобранными в потомстве от свободного опыления первичных дигаплоидов сорта Jubel, которые использовались в качестве доноров генов фертильности. Среди полученных гибридов и в последующих поколениях беккросса на IGC 92/1.2 отбирали наиболее фертильные формы (прежде всего, по показателям ФФП), то есть применяли мягкую форму повышения степени гомозиготности материала, так как самоопыление и сестринские скрещивания давали сильно инбредированное стерильное потомство с низкой жизнеспособностью. В результате получен ряд высокофертильных вторичных дигаплоидов, предназначенных для повышения мужской фертильности диплоидного исходного материала (35).
Установлено, что гибридизация мужски стерильных первичных дигаплоидов картофеля с полученными донорами фертильности обеспечивает получение фертильного потомства, которое может эффективно использоваться в маркер-опосредованной селекции на диплоидном уровне. Так, в случае линии IGC 01/59.11 частота высокофертильных гибридов в потомстве первичного дигаплоида сорта Альпинист составила 100 %, а в потомстве дигаплоида сорта Nortena — 53 % (36). В настоящее время в скрещиваниях с первичными дигаплоидами в качестве доноров генов фертильности мы используем высокофертильные клоны гибридной популяции IGC 10/1.n, полученной в 2010 году в результате самоопыления линии — донора фертильности IGC 01/61.40. Эти клоны обеспечивают получение фертильных гибридов как в скрещиваниях с первичными дигаплоидами (гены фертильности в гетерозиготном состоянии), так и в скрещиваниях между собой (гены фертильности в гомозиготном состоянии) (рис. 1).
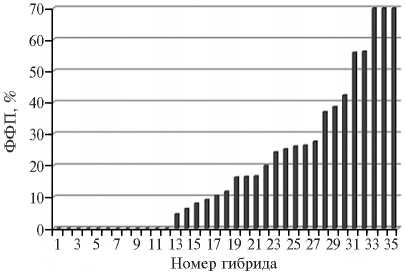
Рис. 1. Функциональная фертильность пыльцы ( ФФП) у гибридов между высокофертильными диплоидными клонами картофеля IGC 10/1.1 и IGC 10/1.23 . Несмотря на близость происхождения родительских форм (получены в результате самоопыления относительно гомозиготной диплоидной линии — донора фертильности IGC 01/61.40), более половины гибридов обладают высокой (> 10 %) ФФП.
Таким образом, гибридизация между донорами фертиль- ности и первичными дигаплоидами, отобранными по жизнеспособ- ности, продуктивности, культурным признакам и комплексу маркеров генов устойчивости к болезням и вредителям, дает исходный материал, пригодный для полноценной селекции по этим признакам на диплоидном уровне. При проведении такой селекции несложно поддерживать приемлемый уровень мужской фертильности, включая в гибридизацию генотипы с высокой ФФП. На основе лучших диплоидных линий с помощью мито- тического удвоения хромосом можно получать эффективные мультиплексные тетраплоидные родительские линии.
Для выведения диплоидных родительских линий, формирующих нередуцированную пыльцу, необходим несколько другой исходный материал: отобранные по комплексу хозяйственно ценных признаков генотипы должны быть пригодны к мейотическому удвоению хромосом (по нашему опыту, ФФП и доля 2 n -пыльцы должны быть не менее 10 %). И такие генотипы должны встречаться на заключительных этапах диплоидной селекции с достаточно большой частотой. Считается, что частота генотипов, формирующих нередуцированные гаметы, среди дигаплоидов и диплоидных видов картофеля достаточна для успешного использования мейотической полиплоидизации (8). Однако, по нашим данным многолетнего изучения большой коллекции вторичных дигаплоидов картофеля разного происхождения, доля генотипов, образующих фертильную нередуцированную пыльцу, составляла лишь 0,8 % от общего числа проанализированных форм (35), что явно недостаточно для успешной селекции по комплексу признаков.
Создание доноров генов формирования нередуцированных гамет. В результате нарушений мейоза могут формироваться нередуцированные (или 2 n ) гаметы, которые имеют число хромосом спорофита, а не гаметофита. Известно несколько видов таких нарушений, приводящих к формированию 2 n -гамет. Для селекции диплоидных родительских линий картофеля наибольшее значение имеет механизм реституции по типу первого деления мейоза — FDR (англ. first division restitution).
А
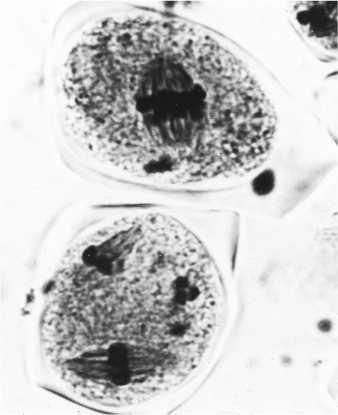
Рис. 2. Нарушения в мейозе у дигаплоидов картофеля ( Solanum tuberosum L.), связанные с формированием нередуцированных гамет FDR (first division restitution) типа: А — слившиеся веретена во втором мейотическом делении вследствие проявления мутации fs (вверху) и два веретена деления при нормальном мейозе (внизу); Б — образование диад 2 n -микроспор в результате нарушений мейоза, связанных с мутацией fs (внизу слева — нормальная тетрада n -микроспор) (фото В.Е. Подлисских).
Б
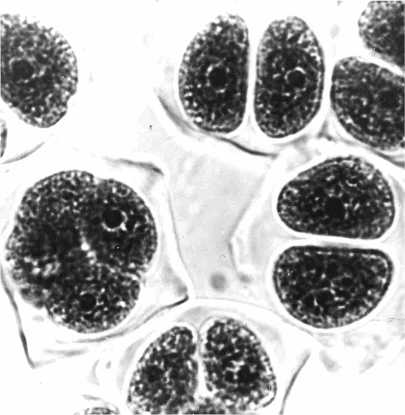
FDR связывают с появлением слившихся ( fs ) (37) или параллельных ( ps ) (39) веретен деления в метафазе II. В мутантных клетках в результате второго мейотического деления формируются только два полюса с двумя группами хромосом и лишь одна клеточная стенка эквационного деления, в результате чего образуется диада 2 n -микроспор (рис. 2). В нашей лаборатории проведено специальное исследование роли коориентации веретен в FDR. Изучение корреляций между частотой ps и частотой диад у 56
форм, продуцирующих и не продуцирующих 2 n -пыльцу, с применением математического моделирования позволило получить доказательства того, что параллельные веретена не вызваны нарушением мейоза, а образование диад связано с мутацией fs (39). При FDR хроматиды каждой хромосомы попадают в разные 2 n -споры, что обеспечивает сохранение родительской гетерозиготности в сформировавшихся гаметах (в среднем степень родительской гетерозиготности в FDR-гаметах составляет около 80 %) (40). Важно отметить, что в 2 n -споры попадает по одной хроматиде каждой из 24 хромосом генотипа, претерпевших процесс рекомбинации при первом делении мейоза. Следовательно, в каждой из них представлены все доминантные аллели генов устойчивости к болезням и вредителям, которые имелись у продуцента нередуцированной пыльцы.
Качественный признак наличия либо отсутствия слияния веретен в мейозе и формирования 2 n -пыльцы имеет количественный характер проявления. Частота формирования 2 n -пыльцы у растений в природных популяциях видов картофеля и у образцов селекционного материала варьирует в широких пределах — от долей процента до 100 % (38). Экспрессивность генов fs (частота 2 n -пыльцы) определяется генотипом, факторами среды и их взаимодействием (41). Предложена модель генетического контроля fs , где главные гены (ген) взаимодействуют с полигенами (генетическим фоном, модифицирующим экспрессию главных генов) (42). С применением методологии анализа количественных признаков показано, что частота формирования 2 n -пыльцы (вариация экспрессивности fs ) определяется действием двух-четырех локусов с равными эффектами (43). Продемонстрирована возможность повышения частоты 2 n -пыльцы при рекуррентном отборе (44). С помощью такого отбора нами созданы диплоидные линии картофеля, формирующие 80-100 % 2 n -пыльцы (35).
Наш опыт изучения генетического контроля формирования нередуцированной пыльцы при гибридизации дигаплоидов картофеля можно суммировать следующим образом (45). Во-первых, при скрещивании дигаплоидов, не образующих 2 n -пыльцу, вероятность выщепления гибридов, способных формировать нередуцированную пыльцу, крайне низкая. При этом частота диад у гибридов, образующих нередуцированную пыльцу, как правило, невысока и недостаточна для мейотического удвоения хромосом. Во-вторых, скрещивания, в которых оба родителя образуют нередуцированную пыльцу, позволяют гарантированно получить гибриды, определенная доля которых с достаточной для мейотического удвоения частотой продуцирует 2 n -пыльцу. Эта доля тем выше, чем выше частота нередуцированной пыльцы у каждого из родителей. Возможно получение гибридов, которые формируют 2 n -пыльцу с более высокой частотой, чем у лучшего по этому показателю родителя. В-третьих, гибридизация между дигаплоидами, один из которых продуцирует 2 n -пыльцу с частотой 80-100 % (донор генов этого признака), а второй ее не образует, не гарантирует получения потомства, способного формировать нередуцированную пыльцу с частотой, достаточной для мейотического удвоения хромосом. Небольшое количество таких гибридов получается только в случае, если в родословной родителя с нормальным мейозом имелись генотипы, продуцирующие 2 n -пыльцу.
Таким образом, для того чтобы на завершающем этапе диплоидной селекции выделить генотипы, обладающие (наряду с комплексом хозяйственно ценных признаков) способностью формировать функционально фертильную нередуцированную пыльцу с достаточной для мейотического удвоения хромосом частотой, необходимо, чтобы оба родителя имели гены этого признака. Следовательно, гены, ответственные за формирование 2n- пыльцы, должны быть введены в генетический пул селекционного материала на ранних этапах диплоидной селекции. Для этого необходимы линии, которые одновременно служат донорами генов фертильности и формирования нередуцированных гамет определенного типа (FDR на основе мутации fs — слившиеся веретена). Созданные нами линии — доноры фертильности практически не образуют 2n-пыльцу. С целью получения линий-доноров по генам фертильности и образованию 2n-гамет доноры фертильности были вовлечены в гибридизацию с вышеупомянутыми линиями, способными с высокой частотой (80-100 %) продуцировать FDR 2n-пыльцу благодаря мутации fs. В результате был отобран ряд гибридов с высокой ФФП и частотой 2n-пыльцы 20-40 %. Они одинаково успешно способны опылять как первичные и вторичные дигаплоиды, так и тетрап-лоидные сорта и селекционные клоны (2).
В рамках программы селекции диплоидных родительских линий, образующих фертильную нередуцированную пыльцу, доноры фертильности и 2 n -гамет были включены в гибридизацию с первичными дигаплоидами с комплексом генов устойчивости к болезням и вредителям. По результатам испытаний 2015-2016 годов отобран ряд перспективных сеянцев, сочетающих высокую продуктивность (на уровне сортов-стандартов), культурные признаки, наличие маркеров нескольких генов устойчивости к болезням и вредителям, которые при этом обладают мужской фертильностью и способны образовывать нередуцированную пыльцу. Так, в 2015 году 43,2 % отобранных по продуктивности сеянцев образовывали функционально фертильную пыльцу, а некоторые из них формировали 2n -гаметы (большинство имело в родословной продуцентов 2 n -пыльцы). Это позволило выделить достаточное количество опылителей для использования в программе маркер-опосредованной селекции родительских линий на диплоидном уровне (табл.).
Характеристика некоторых диплоидных гибридов картофеля ( Solanum tuberosum L.), отобранных для использования в качестве опылителей в маркер-опосредованной селекции родительских линий на диплоидном уровне
|
Гибрид, сорт |
Клубни с растения |
Средняя масса клубня, г |
ФФП, % |
Частота 2n -пыль-цы, % |
ДНК-маркеры генов устойчивости |
||||
|
масса, г |
число, шт. |
H1 TG689 (46) |
Gro1-4 Gro1-4 (47) |
Ryadg RYSC3 (48) |
Sen1 Nl25 (47) |
||||
|
IGC 12/42.1 |
902,3±107,8 |
18,5±2,6 |
57,9±15,3 |
25,8 |
11,0 |
0 |
1 |
1 |
0 |
|
IGC 12/43.3 |
838,2±156,4 |
14,6±1,9 |
62,1±13,3 |
49,5 |
0 |
1 |
0 |
1 |
0 |
|
IGC 12/48.2 |
802,0±122,7 |
22,0±3,2 |
36,5±1,5 |
47,2 |
0 |
1 |
0 |
1 |
1 |
|
IGC 12/42.5 |
672,0±137,9 |
16,7±3,4 |
44,5±6,2 |
44,6 |
16,0 |
0 |
1 |
1 |
0 |
|
IGC 12/42.2 |
573,7±110,9 |
14,3±2,4 |
40,0±3,7 |
11,1 |
9,0 |
0 |
0 |
1 |
0 |
|
IGC 12/42.3 |
529,5±92,3 |
10,5±1,2 |
50,9±6,6 |
56,5 |
0 |
0 |
1 |
1 |
0 |
|
IGC 12/45.11 |
453,0±95,3 |
16,5±3,3 |
28,0±2,3 |
14,0 |
5,0 |
0 |
1 |
1 |
0 |
|
IGC 12/45.12 |
379,5±54,9 |
20,2±1,9 |
19,0±2,37 |
21,3 |
10,0 |
0 |
1 |
1 |
1 |
|
Лилея |
651,6±93,1 |
6,9±0,7 |
88,8±8,3 |
1 |
1 |
0 |
1 |
||
|
Скарб |
894,3±152,8 |
14,2±1,3 |
61,3±7,7 |
1 |
0 |
0 |
0 |
||
|
Рагнеда |
1137,7±216,9 |
16,8±2,3 |
59,7±7,4 |
1 |
0 |
0 |
1 |
||
П р и м еч а ни е. Для показателей продуктивности гибридов приведены средние значения за 2015-2016 годы при 3-кратной повторности, клубни сортов-стандартов (ранний Лилея, среднеспелый Скарб и среднепоздний Рагнеда) в 2015 году высаживали одним блоком в 10-кратной повторности, в 2016 году — тремя рендомизированными блоками по три растения в каждом ( X ±SEM; экспериментальное поле Института генетики и цитологии НАН Беларуси, г. Минск). Функциональную фертильность пыльцы (ФФП) оценивали согласно описанию (24). Нередуцированной (2 n ) считали пыльцу диаметром более 26 мкм (на цитологических препаратах, окрашивание 2 % ацетокармином). У стандартов формирование 2 n -пыльцы не изучали (пропуски в таблице). «1» и «0» — маркер соответственно присутствует или отсутствует.
Таким образом, созданный селекционный материал обладает всеми необходимыми качествами для проведения маркер-опосредованной селекции на диплоидном уровне с получением на завершающем этапе диплоидной селекции ценных генотипов, пригодных для мейотического удвое- ния хромосом.
Перспективы применения маркер-опосредованного отбора на диплоидном уровне в селекции картофеля. Созданный нами диплоидный исходный материал предназначен в первую очередь для получения родительских диплоидных и тетраплоидных линий, использование которых позволит существенно повысить эффективность традиционной селекции картофеля. Однако он представляет интерес и для альтернативных перспективных направлений селекции с применением отбора на диплоидном уровне. Первое направление — выведение диплоидных сортов (лучшие диплоидные гибриды не уступают сортам картофеля по продуктивности и комплексу признаков, а селекция на диплоидном уровне намного эффективнее, чем на тетраплоидном), второе — получение гибридов или гибридных популяций, размножаемых ботаническими семенами. Это могут быть диплоидные гибриды от скрещивания гомозиготных родительских линий, гибридные диплоидные популяции от скрещивания относительно гомозиготных диплоидных родительских линий, тетраплоид-ные гибридные популяции от скрещивания формирующих нередуцированную пыльцу диплоидных родительских линий с сортами картофеля.
Размножение картофеля ботаническими семенами (англ. TPS — True Potato Seeds) предполагает, что для посадки используются не клубни, а семена, полученные в результате самоопыления (свободного опыления) какого-либо сорта (родительской линии) или в результате гибридизации специально подобранных сортов (родительских линий). Возможен также вариант двулетней культуры: в первый год от посева семян получают клубни, которые на следующий год служат посадочным материалом при производстве товарной продукции. Такие клубни занимают по сравнению с обычными семенными клубнями меньший объем и значительно более здоровые, так как не успевают накопить патогены. Это направление считается перспективным для тропических стран, где климатические условия неблагоприятны для семеноводства картофеля, а фермеры не имеют возможности ежегодно закупать импортные семенные клубни и хранить их, самостоятельно репродуцируя (49).
Задача селекции для технологии TPS — получить семена популяций, выравненных по продуктивности и наличию генов устойчивости к патогенам, прежде всего к вирусам и фитофторозу. Традиционные подходы (самоопыление сортов или межсортовая гибридизация) себя не оправдали (49). Однако, как видно из информации, представленной в настоящей статье, эти проблемы могут быть успешно решены с помощью мар-кер-опосредованного отбора на диплоидном уровне с привлечением соответствующего диплоидного исходного материала (формирующего или не формирующего нередуцированные гаметы). Следует отметить, что технология TPS на основе получения диплоидных гибридов при скрещивании гомозиготных родительских линий в последние годы рассматривается как наиболее перспективная для развитых стран, способная революционно изменить существующую систему картофелеводства (34, 50). Одна из крупнейших в мире селекционных компаний «КWS SAAT SE» (Германия) приняла решение сосредоточить свою активность в области селекции картофеля именно на развитии этой технологии (51).
Итак, для использования отбора на диплоидном уровне в селекции картофеля с целью выведения эффективных родительских линий необходимо создание специального исходного материала. Помимо широкого круга первичных дигаплоидов различного происхождения, несущих ДНК-маркеры генов устойчивости к болезням и вредителям, межвидовых ги- бридов с ценными для селекции признаками, он должен включать диплоидные линии — доноры фертильности, самосовместимости и формирования FDR (first division restitution) 2n-пыльцы. Они играют ключевую роль в диплоидной селекции картофеля, так как с их помощью можно получить популяции гибридов, обладающих фертильностью, достаточной для проведения эффективного отбора на диплоидном уровне.
Список литературы Создание исходного материала для маркер-опосредованной селекции родительских линий картофеля (Solanum tuberosum L.) на диплоидном уровне (обзор)
- Bradshaw J.E., Mackay G.R. Breeding strategies for clonally propagated potatoes. In: Potato genetics/J.E. Bradshaw, G.R. Mackay (eds.). CABI, Wallingford (UK), 1994: 109-132.
- Ермишин А.П., Свиточ О.В., Воронкова Е.В., Гукасян О.Н., Лукша В.И. Определение состава и аллельного состояния генов устойчивости к болезням и вредителям у родительских линий картофеля с помощью ДНК-маркеров. Генетика, 2016, 5: 569-578 ( ) DOI: 10.7868/S0016675816050052
- Ortiz R., Peloquin S.J., Freyre R., Iwanaga M. Efficiency of potato breeding using FDR 2n gametes for multitrait selection and progeny testing. Theor. Appl. Genet., 1991, 82(5): 602-608 ( ) DOI: 10.1007/BF00226797
- Montelongo-Escobedo H., Rowe P.R. Haploid induction in potato: сytological basis for the pollinator effect. Euphytica, 1969, 18: 116-123 ( ) DOI: 10.1007/BF00021990
- Clulow S.A., Wilkinson M.J., Waugh R., Baird E., De Maine M.J., Powell W. Cytological and molecular observations on Solanum phureja-induced dihaploid potatoes. Theor. Appl. Genet., 1991, 82: 545-551 ( ) DOI: 10.1007/BF00226789
- Hermsen J.G.Th., Verdenius J. Selection from Solanum tuberosum group Phureja of genotypes combining high frequency haploid induction with homozygosity for embryo-spot. Euphytica, 1973, 22: 244-259 ( ) DOI: 10.1007/BF00022632
- Воронкова Е.В., Лукша В.И., Гукасян О.Н., Савчук А.В., Ермишин А.П. Получение и отбор дигаплоидов S. tuberosum, перспективных для маркер-сопутствующей селекции на комплексную устойчивость к болезням и вредителям. Картофелеводство (Минск), 2011, 19: 215-226.
- Jansky S.H., Peloquin S.J., Yerk G.L. Use of potato haploids to put 2½ wild species germplasm in usable form. Plant Breeding, 1990, 104: 290-294 ( ) DOI: 10.1111/j.1439-0523.1990.tb00438.x
- Ермишин А.П., Маханько О.В., Воронкова Е.В. Использование в селекции соматических гибридов между дигаплоидами картофеля Solanum tuberosum и дикими диплоидными видами из Мексики: получение и беккроссирование дигаплоидов соматических гибридов. Генетика, 2006, 42(12): 1674-1682 ( ) DOI: 10.1134/S1022795406120088
- Ермишин А.П., Маханько О.В., Воронкова Е.В. Получение диплоидного селекционного материала картофеля на основе соматических гибридов между дигаплоидами Solanum tuberosum L. и диким диплоидным видом из Мексики Solanum bulbocastsnum Dunal. Генетика, 2008, 44(5): 645-653 ( ) DOI: 10.1134/S1022795408050086
- Ермишин А.П. Несовместимость при межвидовой и внутривидовой гибридизации диплоидного картофеля и пути ее преодоления. Известия НАН Беларуси, сер. биол. наук, 2001, 3: 105-118.
- Полюхович Ю.В., Маханько О.В., Савчук А.В., Воронкова Е.В., Ермишин А.П. Создание линий-посредников для преодоления межвидовой несовместимости у картофеля. Известия НАН Беларуси, сер. биол. наук, 2010, 2: 51-58.
- Yermishin A.P., Polyukhovich Y.V., Voronkova E.V., Savchuk A.V. Production of hybrids between 2 EBN bridge species Solanum verrucosum and 1 EBN diploid potato species. Amer. J. Potato Res., 2014, 91: 610-617 ( ) DOI: 10/1007/s12230-014-9385-9
- Полюхович Ю.В., Воронкова Е.В., Савчук А.В., Ермишин А.П. Использование Solanum verrucosum для преодоления односторонней несовместимости в скрещиваниях с аллотетраплоидными дикими видами картофеля. Картофелеводство (Минск), 2013, 21(1): 136-145.
- Воронкова Е.В., Лисовская В.М., Ермишин А.П. Диплоидные гибриды между аллотетраплоидными дикими видами картофеля Solanum acaule Bitt., S. stoloniferum Schltdl. и дигаплоидами S.tuberosum L. Генетика, 2007, 43(8): 1065-1073 ( ) DOI: 10.1134/S1022795400708008X
- Ермишин А.П. Генетические особенности аллотетраплоидных диких видов картофеля (Solanum) как объекта селекции. Известия НАН Беларуси, сер. биол. наук, 2014, 1: 23-31.
- Воронкова Е.В., Лисовская В.М., Павлючук Н.В., Савчук А.В., Ермишин А.П. Устойчивость к вирусным болезням потомства диплоидных межвидовых гибридов на основе аллотетраплоидных видов Solanum acaule и S. stoloniferum. Картофелеводство (Минск), 2008, 14: 144-152.
- Воронкова Е.В., Полюхович Ю.В., Савчук А.В., Гукасян О.Н., Ермишин А.П. Полевая устойчивость к фитофторозу гибридов, полученных путем опыления вида-посредника Solanum verrucosum и SvSv-линий на его основе диплоидными 1 EBN видами картофеля. Молекулярная и прикладная генетика (Минск), 2013, 15: 104-110.
- Yeh B.P., Peloquin S.J., Hougas R.W. Meiosis in Solanum tuberosum haploids and haploid-haploid F1 hybrids. Can. J. Genet. Cytol., 1964, 6: 393-402 ( ) DOI: 10.1139/g64-050
- Carroll C.P., Low R.J. Flowering behavior and seed fertility in dihaploid Solanum tuberosum. Potato Res., 1975, 18: 416-427 ( ) DOI: 10.1007/BF00023782
- Trognitz B.R. Female fertility of potato (Solanum tuberosum ssp. tuberosum) dihaploids. Euphytica, 1995, 81(1): 27-33 ( ) DOI: 10.1007/BF00022456
- Carroll C.P., Low R.J. Aspects of male fertility in group Tuberosum dihaploids. Potato Res., 1976, 19: 109-121.
- Trognitz B.R. Comparison of different pollen viability assays to evaluate pollen fertility of potato dihaploids. Euphytica, 1991, 56(2): 143-148 ( ) DOI: 10.1007/BF00042057
- Pallais N., Fong N., Berrios D. Research on the physiology of potato sexual seed production. Proc. Int. Conf. «Innovative methods for propagating potatoes». CIP Rep. 28th Planning Conf. Lima, CIP, 1984: 149-168.
- M'Ribu H.K., Veilleux R.E. Fertility of doubled monoploids of Solanum phureja. Am. Potato J., 1992, 69(7): 447-459 ( ) DOI: 10.1007/BF02852295
- Ермишин А.П., Воронкова Е.В. Показатели фертильности гибридного потомства вторичных дигаплоидов картофеля. Известия НАН Беларуси, сер. биол. наук, 1998, 3: 45-52.
- Grun P. Evolution of the cultivated potato: a cytoplasmic analysis. In: The biology and taxonomy of the Solanaceae. Linnean Soc. Symp. Ser. № 7. Acad. Press, London, 1979: 655-665.
- Sanetomo R., Gebhardt C. Cytoplasmic genome types of European potatoes and their effects on complex agronomic traits. BMC Plant Biol., 2015, 15: 162-178 ( ) DOI: 10.1186/s12870-015-0545-y
- Cipar M.S., Peloquin S.J., Hougas R.W. Haploidy and the identification of self-incompatibility alleles in cultivated diploid species. Can. J. Genet. Cytol., 1967, 9: 511-518 ( ) DOI: 10.1139/g67-055
- Anoshenko B.Yu. Prediction of compatibility between diploid potato varieties. Abstr. 13th Triennial EAPR Conf. Wageningen, NL, 1996: 518-519.
- Olsder J., Hermsen J.G.Th. Genetics of self-compatibility in dihaploids of Solanum tuberosum L. 1. Breeding behavior of two self-compatible dihaploids. Euphytica, 1976, 25(3): 597-607 ( ) DOI: 10.1007/BF00041597
- Cipar M.S., Peloquin S.J., Hougas R.W. Variability in the expression of self-incompatibility in tuber-bearing diploid Solanum species. Am. Potato J., 1964, 41: 155-162 ( ) DOI: 10.1007/BF02855317
- Hosaka K., Hanneman R.E., Jr. Genetics of self-compatibility in a self-incompatible wild diploid potato species Solanum chacoense. 1. Detection of an S locus inhibitor (Sli) gene. Euphytica, 1998, 99: 191-197 ( ) DOI: 10.1023/A:1018353613431
- Lindhout P., Meijer D., Schtte T., Hutten R., Visser R., van Eck H. Towards F1 hybrid seed potato breeding. Potato Res., 2011, 54: 301-312 ( ) DOI: 10.1007/s11540-011-9196-z
- Yermishin A.P. The development of initial parental material for breeding disease resistant potatoes at the diploid level. Plant Breeding and Seed Science, 2000, 44: 105-115.
- Лукша В.И., Савчук А.В., Воронкова Е.В., Ермишин А.П. Результаты отбора по признаку «высокая функциональная фертильность пыльцы» и ряду селекционно важных показателей в популяциях вторичных дигаплоидов Solanum tuberosum L. Картофелеводство (Минск), 2010, 17: 137-148.
- Ramanna M.S. A re-examination of the mechanisms of 2n-gamete formation in potato and its implications for breeding. Euphytica, 1979, 28: 537-561 ( ) DOI: 10.1007/BF00038921
- Mok D.W.S., Peloquin S.J. The inheritance of three mechanisms of diplandroid (2n pollen) formation in diploid potatoes. Heredity, 1975, 35: 295-302 ( ) DOI: 10.1038/hdy.1975.100
- Подлисских В.Е., Анкудо Т.М., Аношенко Б.Ю. Особенности формирования веретена деления и поведения хромосом в мейозе у образцов диплоидного картофеля с мутацией «слияние веретен». Цитология, 2002, 44: 996-1003.
- Hermsen J.G.Th. Mechanisms and genetic implications of 2n-gamete formation. Iowa State J. Research, 1984, 58: 421-434.
- Bani-Aameur F., Laurer F.I., Veilleux R.E. Frequency of 2n pollen in diploid hybrids between Solanum phureja and Solanum chacoense. Potato Res., 1992, 35: 161-172 ( ) DOI: 10.1007/BF02357610
- Jacobsen E. Diplandroid formation and its importance for the seed set in 4½ ½ 2½ crosses in potato. Z. Pflanzenzuchtg, 1980, 84: 240-249.
- Ortiz R., Peloquin S.J. Recurrent selection for 2n gamete production in 2½ potatoes. J. Genet. Breed., 1992, 46: 383-390.
- Mooney J., Peloquin S.J. Phenotypic recurrent selection for 2n pollen frequency. Am. Potato J., 1992, 69: 599.
- Ермишин А.П. Мейотическое удвоение хромосом в селекции картофеля с использованием отбора на диплоидном уровне. Молекулярная и прикладная генетика (Минск), 2013, 15: 39-47.
- Galek R., Rurek M., De Jong W., Pietkievicz G., Augustyniak H., Sawicka-Sienkiewicz E. Application of DNA markers linked to the potato H1 gene conferring resistance to pathotype Ro1 of Globodera rostochiensis. J. Appl. Genet., 2011, 52: 407-411 ( ) DOI: 10.1007/s13353-011-0056-y
- Gebhardt C., Bellin D., Henselewski H., Lehmann W., Schwarzfischer J., Valkonen J.P. Marker-assisted combination of major genes for pathogen resistance in potato. Theor. Appl. Genet., 2006, 112: 1458-1464 ( ) DOI: 10.1007/s00122-006-0248-8
- Kasai K., Morikawa Y., Sorri V.A., Valkonen J.P., Gebhardt C., Watanabe K.N. Development of SCAR markers to the PVY resistance gene Ryadg based on a common feature of plant disease resistance genes. Genome, 2000, 43: 1-8 ( ) DOI: 10.1139/gen-43-1-1
- Almekinders C.J.M., Chujoy E., Thiele G. The use of true potato seeds as a pro-poor technology: the efforts of an International Agricultural Research Institute to innovating potato production. Potato Res., 2009, 52: 275-293 ( ) DOI: 10.1007/s11540-009-9142-5
- Jansky S.H., Charkowski A.O., Douches D.S., Gusmini G., Richael C., Bethke P.C., Spooner D.M., Novy R.G., De Jong H., De Jong W.S., Bamberg J.B., Thompson A.L., Bizimungu B., Holm D.G., Brown C.R., Haynes K.G., Sathuvalli V.R., Veilleux R.E., Miller C., Jr., Bradeen J.M., Jiang J.M. Reinventing potato as a diploid inbred line-based crop. Crop Sci., 2016, 56: 1412-1422 ( ) DOI: 10.2135/cropsci2015.12.0740
- KWS to fully focus on hybrid potato breeding and divest its conventional seed potato business. Режим доступа: http://kws.com/aw/KWS/company-info/Potatoes/News-Articles/-hjqx/KWS-to-fully-focus-on-hybrid-potato-bree/. Без даты.
