Специфические свойства рецепторов андрогенов человека

Автор: Дегтярь В.Т., Кушлинский Н.Е.
Журнал: Сибирский онкологический журнал @siboncoj
Рубрика: Обзоры
Статья в выпуске: 3 (15), 2005 года.
Бесплатный доступ
Короткий адрес: https://sciup.org/14054237
IDR: 14054237
Текст обзорной статьи Специфические свойства рецепторов андрогенов человека
Биологическое действие стероидных гормонов в пределах физиологических концентраций в организме животных и человека обусловлено их проникновением в клетку и обязательным связыванием с внутриклеточными белками-рецепторами, а рецепторный механизм является основным и определяющим в ответе клетки-мишени на действие стероидного гормона. Рецепторы андрогенов (РА) человека изучаются более 20 лет, но наиболее впечатляющие успехи достигнуты в последнее время. Это в первую очередь связано с тем, что рак предстательной железы (РПЖ) был и остается одной из основных причин смерти мужчин пожилого и старческого возраста.
В данном обзоре обсуждаются некоторые специфические, в определенном смысле уникальные, свойства РА человека.
Специфические свойства механизма действия андрогенов и их рецепторов
РА модулируют важные нормальные клеточные процессы и действуют как транскрипционный фактор в контроле роста клеток, дифференциации, пролиферации и апоптоза в клетках-мишенях для андрогенов не только у мужчин, но и у женщин, а ген АР находится на высококонсервативном участке X хромосомы млекопитающих - Xqll -12 [19]. Как транскрипцион- ный фактор РА относится к лигандзависимому типу группы ЗС Ш класса (подсемейства) суперсемейства ядерных рецепторов [2, 3].
Действие андрогенов на органы-мишени в норме регулируется изменением концентрации гормонов в крови. Соотношение индуцируемых (стимулируемых, up-regulation) и репрессируемых (подавляемых, down-regulation) генов может быть близко единице, но изменение андрогенного сигнала обычно приводит к изменению этого соотношения, что, безусловно, очень важно" для регуляции [2,3,29].
На основании многочисленных исследований предложены следующие основные механизмы действия андрогенов на клетку-мишень:
-
1) диффузия (возможно, регулируемая) андрогена (в основном тестостерона, Т) в клетку благодаря наличию положительного градиента концентрации [10,15];
-
2) превращение Т под действием 5а-редук-тазы (5а-Р) в 5а-дигидротестостерон (ДТ) [1, 5,15];
-
3) липофильное взаимодействие ДТ (или Т) (связывание лиганда) с определенным участ ком молекулы РА, которая в цитоплазме нахо дится в составе мультибелкового комплекса, приводит к ряду физико-химических преобра зований этого мультибелкового комплекса и самой молекулы РА; конфирмационные изме нения белковой цепи РА, его диссоциация из мультибелкового комплекса, гомодимеризация,
СИБИРСКИЙ ОНКОЛОГИЧЕСКИЙ ЖУРНАЛ. 2005. №3 (15)
демаскировка определенного(ых) участка(ов) молекулы рецептора, которая в комплексе с лигандом становится более компактной и стабильной. Все эти сложные внутримолекулярные преобразования получили название «активации -РА» [26];
-
4) перенос (транслокация) активированной формы РА в ядро [10, 15];
-
5) связывание активированной формы РА с определенными акцерторными (андрогенчув-ствительными элементами - AREs) участками ДНК [10,15,29];
-
6) реализация специфического ответа - из менение экспрессии определенных генов и син теза специфических белков (ферментов, рецеп торов и т.п.) [10, 15,26].
Предложенная более 20 лет назад указанная принципиальная схема действия андрогенов остается верной, но в последнее время стали известны многие детали этой сложной молекулярной «машины», которая принимает внеклеточный андрогенный сигнал и преобразует его в ответ клетки-мишени.
Из многих природных стероидов, циркулирующих в крови организма, в соответствующих клетках-мишенях со специфическим рецептором для определенного класса стероидов с высоким сродством связывается, как правило, только один наиболее активный и проявляет свое биологическое действие как стероид-эф-фектор. Однако для андрогенов характерно важное отличие: хотя на уровне гена в мужских половых органах активным андрогеном считается ДТ [5], в пределах физиологических концентраций с диким типом РА человека связываются стехиометрически в соотношении 1:1 [11] и могут быть эффекторами два андрогена -Т и ДТ, но сродство ДТ к РА в несколько раз выше [6]. По этой причине 5а-восстановление Т (превращение Т в ДТ) в клетке-мишени некоторые исследователи называют амплификацией андрогенного сигнала [6], т.е. Т можно считать и гормоном-эффектором и прегормо-ном [1]. Указанное свойство - одно из самых необычных: именно для рецепторной системы андрогенов, так как эффекторы для других стероидов поступают в клетки-мишени уже в виде активных молекул.
Относительно более низкое сродство (или высокая степень диссоциации) Т по сравнению с ДТ связано, вероятно, непосредственно со стабильностью молекулы РА в составе комплекса с лигандом; при физиологических концентрациях (0,1 - 5 нМ) Т диссоциирует из комплекса с РА в три раза быстрее, чем ДТ , а расщепление РА идет в два раза быстрее в комплексе с Т. Видимо, по этой причине в ПЖ, в которой преобладающим андрогеном является ДТ, молекула РА более стабильна, особенно в ядрах клеток ПЖ, где концентрация ДТ может быть выше 250 нМ [5, 23].
Одна из возможных причин наличия двух андрогенов-эффекторов в крови человека - специфичное биологическое действие Т и ДТ на ранних стадиях эмбриогенеза. Т как андроген-эффектор играет исключительно важную роль в развитии и дифференцировке структур, относящихся к Вольфову протоку, а ДТ - при развитии структур урогенитального синуса [4]. Некоторые исследователи полагают, что ДТ как андроген-эффектор действует только в ПЖ и семенных пузырьках [23], хотя в большинстве органов человека и животных обнаружена относительно высокая активность 5а-Р [1]. Тем не менее строгого объяснения необходимости для организма человека двух андрогенов-эффекторов, которые образуют комплекс с РА с разными свойствами, до сих пор нет, и идентифицирован один ген AR [4, 29].
Как ни странно , в настоящее время нет единого мнения относительно того, находятся ли РА в цитоплазме клетки-мишени в виде апобелка в составе комплекса с белками теплового шока и другими шаперонами [27] и перемещаются в ядро только после связывания со стероидом, или в нормальных условиях РА постоянно находятся в ядре, а ДТ из цитоплазмы проникает в ядро, где и происходит «активация» комплекса РА-ДТ . Мнения исследователей отличаются существенно:
-
1) при физиологических условиях не только основная часть РА сконцентрирована в клеточ ном ядре, но и в отсутствие ДТ остается там [29];
-
2) свободные РА могут находиться и в ци топлазме, и в ядре, а присутствие лиганда и его связывание с РА регулирует распределение
рецептора между цитоплазмой и ядром [14,29].
Иммунохимически было обнаружено, что РА может оставаться в ядре и в отсутствие лиганда, хотя распределение свободных РА между цитоплазмой и ядром специфично для разного типа клеток [4,9]: в клетках, в которых постоянно присутствует андроген, цитоплазма является только переходным местом после неосинтеза, а специфичное внутриклеточное распределение РА зависит не только от присутствия лиганда, но и других факторов.
Большинство исследователей, однако, полагает, что в нормальной клетке свободный РА в составе мультибелкового комплекса находится именно в цитоплазме [7,29], а эффектор «заставляет» РА (в виде комплекса) перемещаться в ядро [15, 20]. Гомодимер РА также в цитоплазме связывается с импортинами, которые являются шаперонами при транслокации активированного гомодимерного комплекса РА-ли-ганд в ядерную структуру, а один из белков -HSP70 - обнаружен в ядре в составе комплекса РА-ДНК, где он, возможно, повышает связывание РА с ДНК [10, 12]. Возможно, шапероны участвуют и в «челночном» механизме переноса РА из ядра в цитоплазму: совершенно неожиданно оказалось, что молекулы РА могут несколько раз перемещаться из цитоплазмы в ядро и обратно, т.е. имеется механизм, с помощью которого одна молекула РА, вероятно, неоднократно участвует в переносе андрогенного сигнала [29].
На ряде культур клеток в динамике было показано вызываемое андрогенами перемещение РА из цитоплазмы в ядро, которое полностью заканчивалось в течение 60 мин [29]. В среде без андрогена РА возвращались в цитоплазму. В условиях полного ингибирования синтеза белка был показан четырехкратный переход РА из цитоплазмы в ядро и обратно, но, возможно , в нормальных условиях под воздействием гормонального сигнала это происходит и большее число раз [29]. Способность молекулы РА участвовать многократно в передаче андрогенного сигнала указывает на то, что диссоциация комплекса РА-лиганд и/или инактивация лиганда в ядре могут играть более принципиальную роль в прерывании (останов- ке) ответа клетки на андрогенный сигнал [29]. Важно также отметить, что независимо от того, где находится рецептор стероидного гормона без лиганда (в цитоплазме или в ядре), его конформация стабилизирована шаперонами, которые образуют с рецептором комплекс. Возможно, связывание рецепторов с определенными шаперонами препятствует активации рецептора в отсутствие лиганда, что имеет большой биологический смысл [27]. С точки зрения физиологической эффективности клетки именно в случае андрогенов и РА с их «челночным» механизмом, вероятно, имеет больший смысл держать рецептор функционирующим в течение нескольких циклов при передаче гормонального сигнала, а не расщеплять его после каждого цикла гормональной стимуляции: сам рецептор в течение определенного времени не подвергается распаду (расщеплению), а инактивируется лиганд, и его диссоциация из комплекса с РА является, по-видимому, основным фактором прерывания андрогеного сигнала [29]. Уникальный «челночный» механизм действия рецепторов пока обнаружен только для РА [29]. Возможно, это связано со специфичными механизмами инактивации ДТ в клетках [1].
Предполагается, что комплекс лиганд - рецептор в ядре локализуется в субъядерных образованиях (компартментах), природа которых не совсем ясна [29]. Эти субъядерные образования, чаще всего называемые nuclear foci, видимо, являются местами скопления (связывания) для рецепторов (NLS) и других факторов, например корегуляторов , которые являются необходимыми компонентами при экспрессии генов-мишеней под действием андрогенов [34]. Важно, что только РА, связанный с транскрипционно-активными гормонами-лигандами и частичными агонистами (например, ципроте-рон-ацетат), может импортироваться в ядро и оставаться в nuclear foci, в то время как чистые антагонисты (например, касодекс) не обладают этим свойством [34]. Вероятно, причина этого - разная топография комплекса, который образуется после взаимодействия рецептора с разными лигандами. Следовательно, транскрипционно-активный лиганд необходим не только для «активации» РА и его транслока- ции из цитоплазмы в ядро, но важен и для связывания с AREs.
Дня активации регуляторных генов гомодимер комплекса РА - лиганд соединяется с акцепторными участками [10]. В промоторных участках AREs за один акт действия молекула рецептора, вероятно, узнает только несколько генов, однако считается, что РА-зависимой сигнальной системой регулируется транскрипция сотен генов [32].
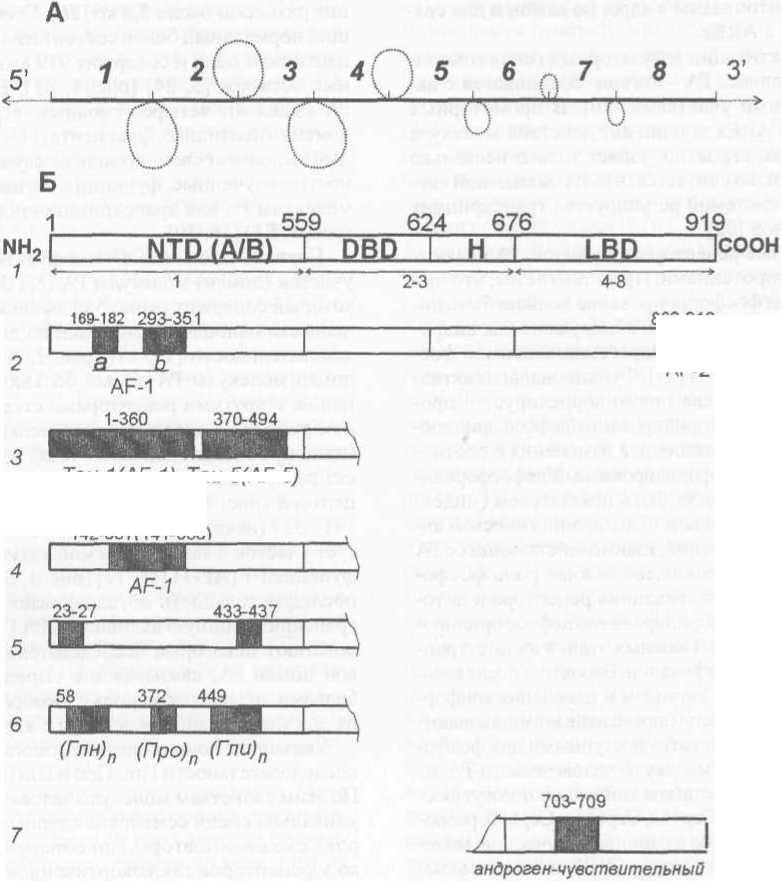
Как и все рецепторы стероидов, РА являются фосфопротеинами. Предполагается, что при активации фосфорилирование должно быть ли-ганд-зависимо, однако обнаружено как андро-гензависимое, так и андрогеннезависимое фосфорилирование РА [33]. Функциональная активность РА человека прямо коррелирует с процессами их фосфоршшрования/дефосфорилиро-вания в клетке-мишени, а изменения в соотношении форм фосфорилированный/дефосфорили-рованный РА могут быть показателем (индексом) агонистической vs антагонистической активности соединений, взаимодействующих с РА [33]. Точно установлена важная роль фосфорилирования в «активации» рецептора в цито-зоштазме, а фосфорилирование/дефосфорилиро-вание РА - один из важных этапов их посттрансляционной модификации. Вероятно , после взаимодействия РА с лигандом и изменения конформации белковой цепи определенные аминокислотные остатки становятся доступными для фосфорилирования. В молекуле человеческого РА из потенциальных остатков аминокислот могут фосфорилироваться Сер453, Сер641 и СербЗО, расположенные в районе «шарнирного участка» молекулы - рецептора - между ДНК- и лигандсвязы-вающими доменами [7] (рис. 1, Б 1).
РА человека - самый большой белок [5, 19, 24] из семейства ядерных рецепторов с хорошо охарактеризованными доменами, а специфические свойства каждого домена имеют важное значение для активности рецептора [2,3,11,13].
Ген человеческого АР (около 90 килобайс, кб) клонирован независимо несколькими группами исследователей в 1988 г. (рис. 1, А) [18, 24]. Транскрипт человеческого АР после сплайсинга и процессинга размером около 11 кб оснований содержит открытую рамку считыва- ния размером около 2,8 кб [26]. Соответствующий нормальный белок состоит из одной поли-пептидной цепи и содержит 919 аминокислотных остатков [5, 24] (рис. 1, Б1). В молекуле РА выделяют четыре основных структурных домена (пептидных фрагмента) [11,29], и каждый выполняет свои, во многих случаях не полностью изученные, функции в активности всей молекулы РА как транскрипционного фактора (рисЛ,Б1)[16,29].
Первый экзон гена АР кодирует N-концевой участок (домен) молекулы PA (NTD или А/В), который содержит около 559 аминокислот и у человека наиболее вариабелен по длине и последовательности [11, 13] (рис. 1, А; Б1). Этот домен молекулы РА самый большой по сравнению с другими рецепторами стероидов, и в нем условно выделяют более мелкие участки (transcription activation units - Taus), которые вносят разный вклад в регуляцию активности рецептора (рис. 1, БЗ) [7]. Последовательность 141 - 337 (некоторые полагают 100 - 370) образует участок с так называемой активационной функцией-1 (AF-1) [11,17] (рис. 1, Б2, Б4). Эта последовательность обусловливает основную транскрипционную активность РА [7,11]. Как полагают некоторые исследователи, N-концевой домен РА, связываясь с определенными белками, играет также роль своеобразного якоря для удерживания рецептора в ядре [22].
Указанный домен содержит гомополимерные последовательности Глн, Про и Гли (рис. 1, Б6). По этим свойствам молекула человеческого РА уникальна среди семейства ядерных рецепторов - сходные повторы Глн обнаружены только у рецепторов глюкокортикоидов (РГК) человека и крысиных РА [11, 24]. Полиморфизм N-концевого домена у здоровых индивидуумов обусловлен наличием гомополимерных последовательностей Глн (начиная с аминокислоты 58), Про (начиная с аминокислоты 372) и Гли (начиная с аминокислоты 449) [11, 24]. Число остатков Глн может колебаться от И до 35, но в большинстве случаев в норме содержится 21 остаток. Последовательность полиГли может содержать 10-31 остаток, но у основной популяции содержится 24 остатка Гли [11, 29]. Более короткая последовательность полиПро
участок
Tau-1(AF-1) Tau-5(AF-5)
142-3371141-3381
889-912
AF-2
Рис. 1. Схематическая структура гена АР, доменная структура и аминокислотная последовательность РА человека:
А - схема расположения экзонов и нитронов гена АР; петлевые структуры нитронов обозначены пунктиром;
Б - схематическое расположение доменов в молекуле РА и обозначения некоторых последовательностей аминокислот, имеющих важное значение для активности РА (подробности см. в тексте). Стрелками и цифрами внизу схемы Б,1 обозначены экзоны схемы А, которые контролируют синтез определенных аминокислотных последовательностей молекулы РА. На всех схемах цифры вверху - номера аминокислотных остатков общей последовательности молекулы белка, подписи внизу - обозначение некоторых важных последовательностей аминокислот в молекуле белка: NTD (А/В) - модуляторный домен; DBD - ДНК-связывающий домен; Н - шарнирный участок молекулы; LBD - лигандсвязывающий домен; NH2 - N-конец молекулы; СООН - С-конец молекулы содержит обычно 8 остатков и менее вариабельна, но данные о связи этого повтора с функциональной активностью человеческого РА пока отсутствуют [11].
Биологическая функция гомополимерных последовательностей указанных аминокислот РА человека не совсем ясна, однако показано, что чем длиннее последовательность полиГлн, тем ниже трансактивационная активность РА [14]. Более того , данные эпидемиологических исследований на 882 мужчинах показали, что у индивидуумов с более короткими повторами CAG в гене AR обнаруживается и более низкий уровень Т в сыворотке крови, а укорочение первого экзона гена человеческого AR на каждый элемент CAG снижает уровень Т в сыворотке крови таких индивидуумов на 0,74 ± 0,36 % [11]. Кроме того, предполагается, что полиморфность по-лиГлн РА у мужчин может быть связана с предрасположенностью к РПЖ [\ 1, 29,32].
Короткие последовательности N-концевого участка молекулы Фен23 - Глн-Асн-Лей-Фен27 и Трп433 - Гис-Тре-Лей-Фен437 обусловливают N- и С-концевое взаимодействие двух молекул РА при образовании гомодимера, что принципиально важно для общей транскрипционной активности РА (рис. 1, Б5) [17, 30]. Именно этот домен обусловливает узнавание специфической последовательности ДНК и, возможно, селективность ответа между андрогенами и глюкокортикоидами в клетках, которые экспрессируют одновременно РА и РГК, и включает основные детерминанты для трансактивации [14]. Эта часть молекулы (344 аминокислотньгх остатка) (рис. 1, БЗ) имеет тенденцию к образованию а-спиральной структуры, которая также важна для гомодимерзации РА и взаимодействия с определенными белками-регуляторами [28].
Экзоны 2 и 3 кодируют участок белковой цепи 559 - 624 (рис. 1, А; Б 1), который обусловливает связывание РА с соответствующим участком ДНК (DBD) - один из важнейших этапов процесса «запуска машины» регулировки активности андрогензависимых генов . Расположенный в середине белковой цепи РА ДНК-связы-вающий домен очень консервативен по длине, богат остатками Лиз и Apr и обязательно содержит девять остатков Цис, которые пре- допределяют третичную структуру гомодимера РА, как и других рецепторов стероидов [2, И, 13]. Консервативность указанного домена молекулы РА, вероятно, связана со значительным его влиянием на третичную структуру всей молекулы РА, поскольку даже незначительные изменения в последовательности аминокислот на этом участке белковой цепи могут оказывать очень большое влияние на активность молекулы рецептора [2]. На этом участке молекулы РА человека обнаружены основные мутации у индивидуумов с синдромом андрогенной недостаточности [11]. Более того, единственная замена - Арг608Лиз обусловливает не только андрогенную недостаточность, но и рак молочной железы (РМЖ) у мужчин: сниженная функция РА, хотя и защитная при РПЖ, может предрасполагать к РМЖ [11].
Кристаллографическими исследованиями показано, что из 9 остатков Цис этого домена 8 остатков связываются координационными связями с двумя атомами цинка с образованием двух так называемых цинковых пальцев, характерных для всех ядерных рецепторов [2, 3, И]. Пальцевые петлевые структуры, формируемые координационными связями цинка, обусловливают взаимодействие РА с основным желобом двойной спирали ДНК и стабилизацию комплекса ДНК-димер рецептора-лиганд [29]. Модуль, включающий в себя первый (начиная с N-конца молекугтьг РА) «цинковый палец» и так называемую область Р-box, непосредственно узнает полусайт AREs. которые расположены рядом с генами-мишенями [11, 32].
Димеризация комплекса РА-лиганд в гомодимер с участием второго «цинкового пальца» а-спирали Н2 возможна только после связывания с лигандом и распада мультибелкового комплекса. Образование гомодимера - результат внутри- и межмолекулярного взаимодействия между амино- и карбоксильным концами двух молекул рецептора, что формирует антипарал-лельную структуру гомодимера по принципу «голова-хвост». На важность взаимодействия концевых последовательностей молекулы для активности РА указывает следующее: единственная мутация Ала748Тре в 5-й а-спирали LBD приводит к резкому нарушению N/C-вза- имодействия в молекуле РА. Такая мутантная молекула в 5 раз быстрее распадается, чем дикая форма РА, и имеет низкое сродство к белкам теплового шока и компартментам ядра [21]. Некоторые исследователи предполагают, что для N/C-взаимодействия в молекуле РА, вероятно, очень важны аминокислотные остатки Вал716, Лиз720; Фал725 и Арг726 в 4-й а-спирали, Мет742, Гли743, Ала748, Арг752 и Фал754 в 5-й а-спирали; Вал889, Мет894, Ала896, Гли897 и Иле898 [29]. Указанные аминокислотные остатки не только участвуют в формировании связывающего «кармана» для лиганда (см. выше), обусловливают «правильную архитектуру» С-концевого лигандсвязыва-ющего домена LBD, модулируют N/C-взаимо-действие, но и важны для устойчивости комплекса РА-лиганд [31]. Нарушение структуры LBD может приводить к частичной или полной андрогенной нечувствительности - обнаружено более 70 миссенс-мутаций на этом участке белковой цепи РА [16]. В общем, во взаимодействии концов молекулы РА при димеризации, вероятно, принимают участие аминокислотные остатки 1 - 503 и 624 - 919, хотя не все они одинаково важны [16]. Было показано также, что при N/C-взаимодейств и и у РА очень важную роль играют кластеры Фал23 - Гли-Асн-Лей-Фал27 и Трп433 - Гис-Тре-Лей-Фал437 (рис. 1, Б5), липофильное и электростатическое взаимодействие которых не только обусловливает «правильную архитектуру» гомодимера, но и эффективное взаимодействие молекулы РА с корегуляторами [17].
Экзоны 4-8 кодируют последовательность аминокислот 676 - 919 белковой цепи РА, которая содержит так называемую активационную функцию-2 (AF-2) и лигандсвязывающий домен (LBD), в котором особенно важен участок 703 -709 (рис. 1, Б1; Б2 и Б7). Карбоксильный конец молекулы РА также очень консервативен - замена даже одной аминокислоты в нем часто приводит к существенному снижению функциональной активностиРА [11].
Пептидный участок LBD образует структуру из 12 ct-спиралей и играет основную роль в связывании лиганда молекулой РА [29]: 11 а-спиралей образуют гидрофобный лигандсвя- зывающий «карман», а 12-я а-спираль (самая крайняя на Оконце молекулы) является своего рода крышкой для «кармана». Принципиально важным для связывания лиганда, как полагают, считается Мет742 в 5-й а-спирали, поскольку мутация Мет742Вал приводит не только к нарушению связывания лиганда дефектной молекулой РА, но и к ослаблению ее связывания с ДНК [31]. В отсутствие лиганда 12-я а-спираль молекулы РА находится пространственно в стороне от полости «кармана», что делает «карман» доступным для лиганда [29]. Липофильное взаимодействие лиганда с рецептором - проникновение лиганда в «карман», - вероятно, вызывает определенные пространственные изменения в белковой цепи молекулы рецептора, в результате чего 12-я а-спираль перемещается в пространстве и закрывает («flips over») гидрофобный лигандсвязывающий «карман», как крышкой, которая препятствует диссоциации захваченного в «карман» лиганда. Указанные конформационные изменения приводят к образованию структурно новой поверхности молекулы РА, которая необходима для взаимодействия не только с определенным участком ДНК, но и с регуляторными белками [29]. Кристаллографический анализ человеческого РА в присутствии и в отсутствие активного синтетического агониста -R1881 показал, что в лигандсвязывающем «кармане» с лигандом взаимодействуют 18 аминокислотных остатков, образуя в основном гидрофобные связи [29]. По крайней мере две водородные связи - одна между кислородом у С-3 молекулы стероида и остатком Арг752 белковой цепи, а вторая - между 17р-гидроксильной группой и остатком Асн705 - дополнительно стабилизируют комплекс андроген-РА [29].
По конкурентному влиянию на связывание ДТ с РА человека некоторые соединения (ксенобиотики) можно разделить на две группы [29]. Первая группа соединений связывается с РА, но не способна вызывать транслокацию РА в ядро. При этом связывание РА с такими лигандами приводит к раннему блокированию андрогенного действия, вероятно, из-за невозможности «активации» - диссоциации рецептора из мульти-белкового комплекса и/или образования соответствующей «архитектуры» белка как переносчи- ка андрогенного сигнала. Вторая группа соединений может вызывать транслокацию РА, но не способна «запустить» соответствующие механизмы в самом ядре, которые обеспечивают транскрипционную активацию. При действии этой группы соединений «неправильная» ориентация в пространстве 12-й а-спирали белка препятствует взаимодействию между участками AF-1 и AF-2 молекулы РА (рис. I, Б2), что, в свою очередь, мешает рецептору связываться с коактиваторами. Точное знание всех этих процессов на каждом этапе проведения гормонального сигнала, безусловно, необходимо для развития терапевтических методов влияния антиандрогенов на РА человека [29].
Потенциальная способность других стероидов, в частности С19 стероидов надпочечников, связываться с РА, имеет важное значение при прогрессировании РПЖ, когда при гормональной блокаде сохраняется ось гипофиз -надпочечники [26]. Кроме того, показано, что при прогрессирующем РПЖ мутации гена РА могут приводить к экспрессии РА с измененной структурой, имеющей повышенное сродство к надпочечниковым и другим андрогенам [26].
Связывание рецептора стероида с ДНК, безусловно, специфично., хотя это кажется удивительным - до сих пор механизм этой специфичности полностью не ясен, Несмотря на их исключительную функциональную специфичность в физиологическом контексте, рецепторы для андрогенов, глюкокортикоидов, прогестерона и минералокортикоидов могут узнавать одни и те же последовательности ДНК, отвечающие на действие гормона. Этот парадокс уже более 25 лет остается загадкой эндокринологии и может быть решен только при дальнейшем изучении процессов трансформации хроматина и образования связанного с хроматином мультибелко-вого комплекса, характерных для каждого типа стероидного гормона [29]. Следовательно, отсутствие уникальной последовательности ДНК и наличие очень консервативного DBD в молекуле РА могут указывать на то, что специфичность связывания с ДНК и исключительно специфический ответ на действие конкретного сте-роида-эффектора - сложный механизм, для которого связывание рецептора с ДНК является необходимым, но, видимо, не достаточным условием, и в котором участвуют дополнительные (вероятно, белковые) факторы [3, 26].
В настоящее время ясно, что во взаимодействии комплекса РА-лиганд с ДНК и инициации биологического ответа совершенно необходимо участие целого ряда белков-корегуляторов и факторов транскрипции [18]. Обнаружены как не специфические регуляторы действия - общие для стероидных гормонов и общие при механизмах транскрипции, так и специфические регуляторы для отдельного типа стероидов. Некоторые корегуляторы с разными названиями являются отдельными идентичными белками [3], а некоторые образуют целое семейство белков-кофакторов [8]. Среди корегуляторов могут быть как стимуляторы активности рецепторов стероидных гормонов (коактиваторы), так и ингибиторы (корепрессоры) [3]. По крайней мере in vitro показано, что сигнальная система РА человека может также активироваться: с участием ростовых факторов, например фактора роста фибробластов и эпидермального фактора роста [10,26]. Корегуляторные белки, по-видимому, не связываются со специфическими последовательностями ДНК, а действуют как адаптеры (модуляторы) между доменами молекулы рецептора и общим транскрипционным комплексом, образуя мультибелковые комплексы и модулируя функции РА при их транс активации [3,26].
Важно отметить, что во всех без исключения случаях термин «специфический» по отношению к корегупяторам не следует понимать буквально, поскольку данные по их влиянию на активность РА получены с конкретными клеточными системами и в конкретных условиях, а в других системах эти же модуляторы могут и не обладать специфичностью действия по отношению к РА.
Многие обнаруженные коактиваторы РА являются гистон-ацетил-трансферазами, активность которых, как известно, связана с локальной модификацией белков хроматина и имеет принципиальное значение для регуляции экспрессии гена [2, 13], Это позволяет предположить, что такие коактиваторы, взаимодействуя с AF-2 рецептора, действуют и как регуляторы активности промоторов в механизме РНК-полимеразной активности [13].
Особый интерес представляет андрогенне-зависимая активация РА при развитии андрогенной рефрактерности РПЖ, поскольку такая регуляция осуществляется исключительно с участием белков-регуляторов. При этом в рефрактерных клетках транскрипционная активность РА проявляется (поддерживается) независимо от лиганда, и/или в таких клетках гены-мишени для андрогенов могут активироваться независимо от РА, хотя нормальный механизм андрогенной регуляции может оставаться функционально активным [20,26].
Оценивая роль РА как медиаторов генной экспрессии при обязательном участии корегу-ляторов, необходимо отметить, что при положительной (up-regulation) регуляции с участием указанных рецепторов необходимо их прямое связывание с участками AREs молекулы ДНК, хотя и не без участия коактиваторов и, возможно, других белков [3]. Негативная регуляция (down-regulation) осуществляется в основном при белок-белковом взаимодействии с участием киназ, возможно, уже на уровне цитоплазмы, а в ядре создаются пространственные затруднения для связывания РА с ДНК: регуляция на уровне цитоплазмы и ядра может осуществляться независимо и/или параллельно [12]. Точный состав комплекса РА с корегу-ляторами на конечном этапе регуляции экспрессии гена может зависеть от промотора и/или энхансера и типа клетки, что и обусловливает специфичность регуляции андрогенами [28,34].
В настоящее время еще очень много неясного в процессах участия корегуляторов в трансактивации и трансрепрессии стероидами клеток-мишеней, однако ясно одно: белки-ко-активаторы и белки-корепрессоры совершенно необходимые и обязательные участники процесса «приема» гормонального сигнала и ответа клетки-мишени на этот сигнал.
В клетках^шш енях активность РА изменяется при клеточном цикле: андрогены укорачивают длину фазы Gj/GQ и ускоряют вход в S-фазу, влияя на экспрессию и/или активность циклинов и циклинзависимых киназ [25]. При клеточном цикле не только изменяется активность РА, но и меняется его концентрация как белка - наименьшая в фазе Gt/S [25]. В то же время некоторые белки-регуляторы клеточного цикла могут влиять на активность РА, действуя как корегуляторы [25].
Важный вопрос - все ли время, в течение которого РА находится в ядре, он функционирует по типу «челночного» механизма. Вероятно, некоторые стадии не без участия корегуляторов временно могут прерываться. Периоды остановки и нового запуска процессов транскрипции, очевидно , коротки по времени, поскольку в отличие от цитоплазмы субъядерные компартменты не разделены мембранными структурами, что позволяет белкам «быстро двигаться» в пределах ком-партмента [3 4]. Указа! шое движение белков энергетически независимо, и этот или сходный механизм позволяет быстро «включать» в работу и «выключать» транскрипционный фактор в виде рецептора не только при действии андрогенов, но, видимо, и других стероидов [34].
Заключение
Все перечисленные выше факты позволяют считать многие детали механизма действия андрогенов уникальными в сравнении с механизмом действия других стероидов, и эта уникальность в ряде случаев объяснена в досточной степени полно. В то же время две очень важные проблемы ждут своего решения. Во-первых, дискриминатор-ные механизмы передачи андрогенного сигнала при наличии в организме двух андрогенов-лигандов - Т и ДТ, которые хоть и отличаются по активности, но имеют сравнимое сродство к РА. Одно из возможных объяснений: Т является предшественником (прегормоном) не только для андрогенов, но и для эстрогенов [1]. По-видимому, это позволяет поддерживать необходимый баланс в действии половых стероидов в разных клетках, однако это не объясняет полностью необходимость иметь в организме два активных лиганда для РА. Во-вторых, не понятен механизм специфичности действия РА при взаимодействии с AREs на ДНК по сравнению с другими рецепторами стероидов, особенно с РГК. Предполагается, что основной дискриминатор при этом - N-концевой домен молекулы РА человека, который уникален по сравнению с другими рецепторами стероидов (рис. 1, Б2; Б5 и Б6). Этот домен, по-видимому, может связываться специфично с определенными ос- нованиями в пределах коротких последовательностей ДНК, однако проблема специфичности действия андрогенов остается [29]. Не менее важно и дальнейшее исследование механизмов связи «работы» аппарата ядерных рецепторов с «работой» общего аппарата транскрипции в хроматине, поскольку основная функция рецепторов стероидов - обеспечить эффективное взаимодействие между этим аппаратом и хроматиновыми структурами, среди которых этот аппарат «работает» [8].
