Специфика формирования комплекса микромицетов в почвах островной поймы реки Илыч

Автор: Хабибуллина Флюза Мубараковна, Виноградова Юлия Алексеевна, Лаптева Елена Морисовна, Дегтева Светлана Владимировна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Землепользование
Статья в выпуске: 1-8 т.14, 2012 года.
Бесплатный доступ
Исследован комплекс микроскопических грибов в почвах островной поймы реки Илыч (приток р. Печора). Выявлено 90 видов микромицетов из 26 родов. Наиболее представительны роды Penicillium, Mortierella, Aspergillus, Trichoderma. Выделены новые виды микромицетов, не отмеченные ранее в пойменных почвах равнинных рек средней тайги. Показано, что в почвах рассмотренного ряда биотопов (бечевник → луг → лиственный лес → хвойный лес) максимальным α-разнообразием, численностью спор, длиной мицелия и их биомассой отличаются сообщества почвенных микромицетов лиственного леса. В луговых ценозах грибной мицелий концентрируется преимущественно в минеральных (гумусоаккумулятивных А1) горизонтах, в лесных сообществах – в органогенных (лесные подстилки А0) горизонтах аллювиальных дерновых почв.
Микромицеты, аллювиальные почвы, река илыч
Короткий адрес: https://sciup.org/148201006
IDR: 148201006 | УДК: 631.466.1:631.482.1(282.247.111)
Specifics of formation the micromycetes complex in soils of island flood lands at ilych river
The complex of microscopic fungi in soils of the Ilych River (a tributary of the Pechora River) island flood-lands has been studied. 90 species of micromycetes of 26 genera were identified. Most representative were the genera of Penicillium, Mortierella, Aspergillus, and Trichoderma. Some species of micromycetes were first identified for flood soils of middle taiga plain rivers. Among the studied biotopes (towpath→ meadow → deciduous forest → coniferous forest), the highest alpha-diversity, number of spores, biomass of micromycetes and longest mycelium were found out for soil micromycetes of deciduous forest. In meadow coenoses, fungal mycelium mainly concentrated in mineral (humus-accumulative – A1) horizons of alluvial soils whereas in forests it preferred organic (forest litters – A0) soil horizons.
Текст научной статьи Специфика формирования комплекса микромицетов в почвах островной поймы реки Илыч
Одним из важнейших компонентов блока почвенной микробиоты по праву считаются микроскопические грибы [9]. Они принимают непосредственное участие в деструкционных процессах, обусловливая разложение ежегодно поступающего на поверхность почвы растительного опада. Практически во всех типах почв микромицеты по биомассе превышают бактериальную составляющую комплекса микроорганизмов [6]. В настоящее время сообщества микроскопических грибов детально исследованы в зональных почвах Республики Коми [10], а также в аллювиальных почвах равнинных рек средней тайги [2]. Почвы пойменных террас рек предгорных и горных ландшафтов, в том числе и р. Илыч, в этом плане слабо изучены. В связи с тем, что большая часть бассейна Илыча приурочена к территории Печоро-Илычского государственного природного биосферного заповедника, пойменные экосистемы верхнего и среднего течения реки развиваются в режиме спонтанной динамики и характеризуются минимальным антропогенным воздействием, а почвенно-растительный покров островов, соответственно, отражает процесс естественной сукцессии растительности на аллювиальных наносах.
Цель работы: изучение комплекса микроскопических грибов, формирующихся в аллювиальных почвах островной поймы р. Илыч (Печоро-Илычский заповедник).
Объект и методы исследования. Исследования проводили в 2009-2010 гг. на территории островной поймы среднего течения р. Илыч. Вдоль топоэкологического профиля, заложенного от уреза воды к центральной части о-ва Бияизъяди, изучены состав и структура фитоценозов, охарактеризовано морфологическое строение аллювиальных почв, проведен отбор проб для изучения свойств почв (2009 г.) и комплекса микромицетов (2010 г.). Исследования проводили с применением стандартных методов геоботаники и фитоценологии [1, 4], почвоведения и почвенной биологии [3]. Суммарную численность спор микроскопических грибов, длину мицелия, их биомассу определяли методом люминесцентной микроскопии, видовое разнообразие микромицетов – методом посева почвенных вытяжек на специализированные среды [3]. Таксономическую принадлежность микромицетов идентифицировали с использованием определителей [8, 11 и др.], интерактивных «ключей» и информационного сайта интернет-ресурсов (http://www. . Характеристика комплекса микроскопических грибов дана на основе таких показателей, как обилие, частота встречаемости, коэффициент сходства Съеренсена-Чекановского. Результаты обработаны с помощью компьютерной программы «GRAPHS» [5].
Результаты и обсуждение. В пределах ключевых участков (табл. 1), отражающих сукцессионный ряд смены экосистем на аллювиальных наносах о-ва Бияизъяди, происходит закономерное преобразование состава и структуры растительности и связанное с ним изменение почвенного покрова. Наиболее специфичным видовым составом характеризуются растительные сообщества, формирующиеся в полосе экотона вода-суша (1Б). Для них свойственны минимальные показатели видовой насыщенности фитоценозов сосудистыми растениями (от 3 до 10 видов на 100 м2). На I-м уровне островной поймы начинают формироваться сообщества травянистых растений (2Б), в которых значения показателя общего проективного покрытия (ОПП) варьируют от 40 до 60%, а на пробной площади встречается в среднем 25 видов. Максимальное разнообразие травянистых растений (до 46 таксонов на 100 м2) зарегистрировано на лугах (3Б), здесь отмечены также экземпляры кустарников (3 вида). В лесных экосистемах (4Б, 5Б) показатели a-разнообразия сосудистых растений в травяном покрове несколько ниже (30 и 26 видов на 400 м2 соответственно). При этом более выражена роль растений древесной жизненной формы (по 8 видов на пробной площади). На завершающих этапах сукцессий под пологом хвойных дере-вьев-эдификаторов на фоне общего уменьшения видовой насыщенности травяного покрова из-за ухудшения освещенности появляются отчетливо выраженные доминанты из числа теневыносливых трав: Diplazium sibiricum, Athyriumfilix-femina.
Таблица 1. Характеристика пойменных биотопов о-ва Бияизъяди
|
№ уча стк а |
Высотный уровень островной поймы |
Координаты; высота над ур.м. |
Тип растительности |
Тип почвы |
|
1Б |
Бечевник |
62°32'37.1" с.ш. 58°12'55.2" в.д. h=130м |
Разреженные сообщества Petasites radiatus |
Почва отсутствует. Формирование растительности идет на мелкоземе, заполняющем межгалечниковое пространство |
|
2 Б |
I-й уровень |
62°32'36.4" с.ш. 58°13'00.9" в.д. h=134м |
Разнотравное сообщество с содоминированием Hieracium laevigatum , Antennaria dioica , Festuca rubra |
Аллювиальная дерновая примитивная песчаная* |
|
3Б |
II-й уровень |
62°32'36.9" с.ш. 58°13'06.1" в.д. h=136м |
Злаково-разнотравный луг с участием Bromopsis inermis, Thalictrum simplex , Crepis sibi-rica , Anthriscus sylvestris и др. |
Аллювиальная дерновая слоистая супесчаная** |
|
4 Б |
III-й уровень |
62°32'38" с.ш. 58°13'04.4" в.д. h=138м |
Сероольшаник высокотравновейниковый с участием Cala-magrostis purpurea , Aconitum septentrionale , Crepis sibirica , и др. |
Аллювиальная дерновая (лесная) слоистая супесчаная*** |
|
5Б |
III-й уровень, центральная часть острова |
62°32'43.1" с.ш. 58°13'04.8" в.д. h=142м |
Ельник разнотравнопапоротниковый |
Аллювиальная дерновая (лесная) оподзоленная супесчаная*** |
Примечание: подстилание крупным русловым песком с глубины: * 15-25 см; ** 50-70 см, *** 70 см.
По мере развития пойменного острова примитивные аллювиальные почвы (формула строения почвы: АдА1-D) постепенно сменяются аллювиальными дерновыми слоистыми (Адер-А1-I-й слой-...-CD), затем аллювиальными дерновыми (лесными) слоистыми (А0-А1-АВ-В-1-Й слой-...-CD) и дерновыми оподзоленными (A0-A1A2-ABg-Bg-I-й слой-...-CD) почвами. Последние, развитые под хвойными фитоценозами, составляют основу почвенного покрова острова. Минеральная часть профиля аллювиальных лесных почв слабо дифференцирована на горизонты. Их специфической особенностью является наличие горизонта лесной подстилки А0 мощностью до 5-7 см, под которой развит гумусоаккумулятивный горизонт А1 мощностью до 10-15 см. Признаки оглеения прослеживаются локально, на глубине 15-30 см. Благодаря размыванию силурийских и каменноугольных известняков, выходы которых имеются в долине среднего течения р. Илыч, на поверхность пойменной террасы ежегодно откладываются наилки с близкой к нейтральной реакцией среды (рНКС1), содержащие до 6,6 ммоль/100 г обменных оснований и до 2,8% гумуса. Соответственно, почвы, формирующиеся на ранних стадиях сукцессий, характеризуются близкими к наилкам величинами кислотности, насыщенностью основаниями и содержанием гумуса в верхних органогенных горизонтах до 1,8-2,2%. По мере развития древесной растительности, в направлении от аллювиальных примитивных к аллювиальным дерновым слоистым почвам пойменных лесов, возрастает кислотность, снижается степень насыщенности основаниями, уменьшается обогащенность лесных подстилок азотом (рис. 1).
Выщелачивание минеральной толщи почвы более ярко выражено под пологом хвойных фитоценозов. В гор. А1А2 аллювиальных лесных почв (5Б) кислотность солевой вытяжки 3,5-3,6 ед. рН, степень насыщенности основаниями 25-29%, что позволяет говорить о начальных стадиях подзолообразования в аллювиальных почвах островной поймы, формирующихся под пологом ельников травяных. Изменение экологических условий в рассмотренном ряду бечевник → луг → лиственный лес → хвойный лес (смена типа растительности и типа почвы, изменение характера поступающего на поверхность почвы растительного опада, повышение уровня пойменной террасы, снижение уровня почвенно-грунтовых вод, изменение кислотно-основных свойств почв и пр.) не могут не сказаться на состоянии остальных компонентов наземных экосистем, в том числе и на составе комплекса микроскопических грибов.
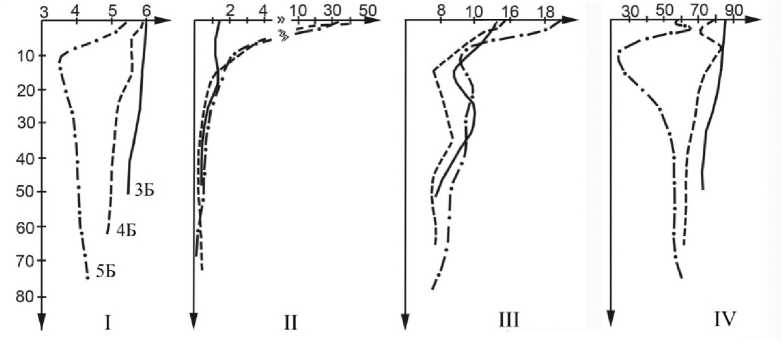
Рис. 1. Профильное распределение величины рН (I; ед.рН), Сорг. (II; %), отношения C/N (III), степени насыщенности основаниями (IV; %) в аллювиальных почвах злаково-разнотравного луга (3Б), лиственного (4Б) и хвойного (5Б) леса
Как показали проведенные исследования, видовое разнообразие микромицетов в аллювии-альных почвах о-ва Бияизъяди существенно выше по сравнению с почвами пойменных террас исследованных нами ранее равнинных рек таежной зоны – Вычегды и Сысолы [2]. В почвах острова в русле р. Илыч обнаружено 90 видов микромицетов из 26 родов, относящихся к двум отделам Zygomycota, Ascomycota и аннаморфным грибам (в том числе два «вида» стерильных форм мицелия), тогда как в почвах долин Вычегды т Сысолы – 67-71 вид мик-ромицетов. Таксономический состав почвенных микоценозов о-ва Бияизъяди, как и других пойменных почв, характеризуется обилием представителей рода Penicillium (30 видов), Mortierella (11), Trichoderma (8). Видовое разнообразие ро-дов Aspergillus , Mucor, Chaetomium, Acremonium ограничивается 4-6 видами. Роды Actinomortierella, Au-reobasidium, Cephalosporium, Chlamydomyces, Dematium, Geomyces, Geotrichum, Gliomastix, Hya-lobotrys, Monilla, Mycogone , Oospora, Phialophora, Paecilomyces, Phoma, Scopulariopsis, Sporotrichum, Stachyborys, Trichosporiella представлены 1-2 видами.
Минимальное число видов (19-25) отмечено в биотопах, формирующихся в экотоне вода-суша и характеризующихся развитием растительности на галечниковых наносах бечевников (1Б) и на примитивных аллювиальных почвах (2Б). Максимальное разнообразие (47 видов) зафиксировано в сообществах лиственного леса, представляющего собой экотонную полосу между злаково-разнотравным лугом (3Б) и хвойным фитоценозом (5Б). Следует отметить, что в подзоне средней тайги лиственные леса на водоразделах также отличаются более значительным видовым разнообразием микромицетов, по сравнению с хвойными биотопами [7].
Микоценозы исследованных сообществ характеризуются высокой специфичностью: коэффициент Съеренсена-Чекановского 30% (рис. 2). Первый кластер включает участки разнотравных сообществ на галечниках с примитивными аллювиальными почвами, второй – участки злаковоразнотравного луга, лиственного и хвойных лесов с более зрелыми почвами, относящимися к типу аллювиальных дерновых почв (рис. 2).
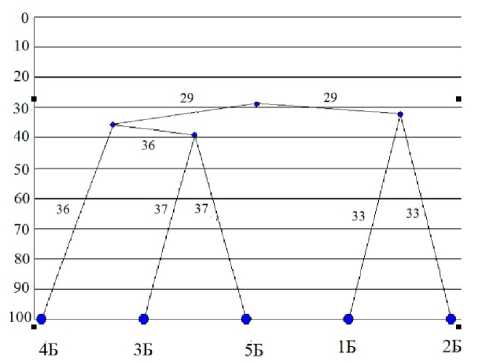
Рис. 2. Дендрограмма сходства комплекса микроскопических грибов в аллювиальных почвах о-ва
Бияизъяди. Условные обозначения по горизонтали те же, что и в табл. 1. По вертикали – коэффициент Съеренсена-Чекановского, %
Детальный анализ таксономической структуры микоценозов позволил выявить как широко распространенные виды, так и виды, приуроченные к конкретным местообитаниям. Так, например, во всех биотопах встречаются Geomyces pannorum (Link) Sigler & J.W. Carmich , Umbe-lopsis ramanniana (Möller) Linnem., Paecilomyces variotii Bainier, Рenicillium Waksmani K.M. Zallessky,
Trichoderma viride Persoon ex Fries, Mycelia sterilia (Moniliaceae), Mycelia sterilia (Dematiaceae). В то время, как для микоценозов галечников характерны Мortierella marbugensis Linnem- =-(M.verti-cillata), Oospora variabilis Linder ., Penicillium glau-cogriseum Sopp , почв разнотравного сообщества – Mucor alboater Naumov, Aspergillus candidus Link , злаково-разнотравного луга – Mortierella bira-mosa Tiegh ., Penicillium niger Tiegh (= Aspergillus niger var. Niger ), Cephalosporium charticola J.Lindau , сероольшанника разнотравно-вейникового – Mortierella humicola Ondem , Mucor hiemalis Wehmer , M. raсemosus Fresen. , Ch. spiralliforum Bainier (= Ch. spirilliferum), ельника разнотравнопапоротникового – Mortierella minutissima Tiegh. (=M.minutissima var. minutissima), Mortierella verticillata Linnem. , Aspergillus versicolor (Vuill.) Tirab ., Рenicillium chrysogenum Thom , P. martensii Biourge.
В почвах островной поймы р. Илыч выявлены виды, не отмеченные ранее в долинах рек Вычегды и Сысолы: Mortierella alpinа Peyron, M. marbugensis Linnem=(M.verticillata), Geomyces pannorum (Link) Sigler & J.W. Carmich, Oospora varia-bilis Linder, Gliomastix cerealis (P.Karst), Penicillium kapuscinskii Zaleski; Raper a. Thom (= Penicillium canescens), P. kursanovii Chalabuda (= Penicillium restrictum), Trichoderma harzianum Rifai. Обнаружение новых видов микроскопических грибов может быть обусловлено спецификой экологических условий формирования почв в условиях островной поймы р. Илыч: небольшой период затопления паводковыми водами, близкая к нейтральной реакция среды, отсутствие антропо-генного влияния на почвенно-растительный покров островной поймы.
При оценке численности и биомассы микроскопических грибов в рассмотренном сукцессионном ряду обращает на себя внимание отсутствие в субстратах участков 1Б и 2Б мицелия грибов (табл. 2). Микроскопические грибы в проанализированных образцах в основном находятся в виде спор, которые при соответствующих условиях могут активизироваться и давать рост колоний. В почвах, развитыми под остальными фитоценозами, основная доля биомассы приходится на грибной мицелий (94,7-98,3%), что свидетельствует об активном протекании здесь процессов деструкции растительного опада. В лесных сообществах (участки 4Б, 5Б) споры и мицелий грибов концентрируются преимущественно в горизонтах лесных подстилок. Вниз по профилю происходит постепенное снижение всех количественных показателей. Особое место в исследованном ряду занимает почва злаково-разнотравного луга (3Б). Здесь отмечено смещение максимальных значений показателей длины и биомассы мицелия грибов в гумусоаккумулятивный горизонт. Это может быть обусловлено наличием хорошо развитых корневых систем травянистых растений и более высоким содержанием органического вещества (Сорг. составляет 2,2%).
Таблица 2. Численность и биомасса микромицетов по данным посева на специализированные среды (I) и люминесцентной микроскопии (II)
|
Ключе-вой участок |
Горизонт |
Численность |
Биомасса |
|||
|
I |
II |
|||||
|
среда Чапека, тыс.КОЕ/г |
споры, млн. кл/г |
длина мицелия, м/г |
споры, мг/г |
мицелий, мг/г |
||
|
1Б |
I-й сл. |
55,42 |
15,37 3,5 9-27, 15 |
0 |
0,15 0,0 4-0, 27 |
0 |
|
2Б |
АдА1 |
9,50 |
3,58 0,7 9-6, 39 |
0 |
0,03 0,0 1-0, 06 |
0 |
|
3Б |
АдА1 |
152 |
12,87 11,77-13,98 |
763 697-829 |
0,13 0,12-0,14 |
2,98 2,72-3,23 |
|
А1 |
6,3 |
8,56 7,67-9,45 |
1096 915-1278 |
0,09 0,08-0,10 |
4,28 3,57-4,99 |
|
|
I-й сл. |
3,15 |
6,0 5,53-6,47 |
888 646-1130 |
0,06 0,055-0,065 |
3,46 2,52-4,41 |
|
|
4Б |
А0 |
250,43 |
27,11 25,18-29,04 |
1908,5 1299-2519 |
0,28 0,25-0,29 |
7,28 5,07-9,82 |
|
А1 |
51 |
18,96 16,61-21,31 |
810 697-851 |
0,19 0,17-0,21 |
3,4 3,0-3,32 |
|
|
АВ |
36 |
9,57 8,61-10,54 |
569 432-706 |
0,10 0,09-0,11 |
2,22 1,68-2,75 |
|
|
5Б |
А0 |
233,4 |
12,42 11,56-13,27 |
1390 1185-1595 |
0,12 0,11-0,13 |
5,42 4,62-6,22 |
|
А1А2 |
127,2 |
8,32 8,04-8,59 |
932 844-1021 |
0,08 0,07-0,09 |
3,64 3,29-3,98 |
|
|
Bg |
71,2 |
6,4 6,23-6,58 |
665 565-765 |
0,06 0,05-0,07 |
2,59 2,2-2,98 |
|
Список литературы Специфика формирования комплекса микромицетов в почвах островной поймы реки Илыч
- Ипатов, В.С. Описание фитоценоза. Методические рекомендации. -СПб., 1998. 93 с.
- Лаптева, Е.М. Разнообразие микромицетов в почвах пойменных лугов/Е.М. Лаптева, Ф.М. Хабибуллина, Ю.А. Виноградова//Микология и фитопатология. 2009. Т.43, вып.3. С. 199-206.
- Методы почвенной микробиологии и биохимии/Под. ред. Д.Г. Звягинцева. -М.: Изд-во МГУ, 1991. 304 с.
- Нешатаев, Ю.Н. Методы анализа геоботанических материалов. -Л., 1987. 192 с.
- Новаковский, А.Б. Возможности и принципы работы программного модуля «GRAPHS». -Сыктывкар, 2004. 28 с.
- Полянская, Л.М. Распределение численности и биомассы микроорганизмов по профилю зональных типов почв/Л.М. Полянская, В.В. Гейдебрехт, Д.Г. Звягинцев//Почвоведение. 1995. №3. С.322-328.
- Разнообразие почв и биоразнообразие в лесных экосистемах средней тайги. -М.: Наука, 2006. 287 с.
- Саттон, Д. Определитель патогенных и условно патогенных грибов/Д. Саттон, А. Фотергиль, М. Ринальди. -М.: Мир, 2001. 486 с.
- Терехова, В.А. Микромицеты в экологической оценке водных и наземных экосистем. -М.: Наука, 2007. 215 с.
- Хабибуллина, Ф.М. Почвенная микобиота естественных и антропогенно нарушенных экосистем Северо-Востока европейской части России: Автореф. дис…докт. биол. наук. -Сыктывкар, 2009. 40 с.
- Ramirez, C. Manual and atlas of the Penicillia. -Amsterdam; New York; Oxford: Elsevier Biomedical Press, 1982. 874 p.
