Сравнение экофизиологических показателей сосны и ели в Серебряноборском опытном лесничестве

Автор: Молчанов А. Г.
Журнал: Лесохозяйственная информация @forestry-information
Рубрика: Лес и климат
Статья в выпуске: 1, 2020 года.
Бесплатный доступ
Приведены результаты оценки влияния окружающей среды на эколого-физиологические показатели ели обыкновенной и сосны обыкновенной. Установлено, что дыхание стволов сосны и ели при расчете за сутки зависит от поступления солнечной радиации: в пасмурные дни интенсивность дыхания в 2 раза слабее, чем в малооблачные. При недостатке влаги интенсивность фотосинтеза ели снижается в большей степени, чем у сосны.
Сосна, ель, интенсивность фотосинтеза, интенсивность дыхания ствола, недостаток влаги
Короткий адрес: https://sciup.org/143170874
IDR: 143170874 | УДК: 630.161.2:630.161.32 | DOI: 10.24419/LHI.2304-3083.2020.1.11
Comparison of the ecophysiological parameters of pine and spruce in the Serebryanoborsky experimental forest district
Research conducted in Serebryanoborsky forestry Institute of Forestry RAS. To evaluate gas exchange, an original setup based on an infrared gas analyzer was used. The installation created at the Forestry Institute of the Russian Academy of Sciences made it possible to simultaneously, round-the-clock receive daily changes in the intensity of photosynthesis and respiration of adult trunks of pine and spruce trees. It was found that the respiration of pine and spruce trunks, calculated on the surface of trunks, does not differ significantly, while the intensity of photosynthesis of spruce (when calculated on the surface area of needles) is almost two times lower than that of pine. However, we believe that when calculating the area of growth, the differences will be insignificant, since the mass of spruce needles in the stand is much greater than the mass of pine needles per unit area of growth. It was found that the intensity of photosynthesis of spruce is more affected by moisture deficiency...
Текст научной статьи Сравнение экофизиологических показателей сосны и ели в Серебряноборском опытном лесничестве
Вследствие изменения климата одни регионы планеты терпят бедствие из-за увеличения количества ураганов и повышения уровня моря, другие пытаются справиться с засухой. При этом, по оценкам экспертов, по мере эскалации процесса глобального потепления площадь территорий, страдающих от засухи, может увеличиться.
При потеплении климата возрастает не только количество осадков, но и усиливается испарение с поверхности почвы, а это существенно уменьшает содержание влаги в деятельном слое почвы (глубиной 1 м). Модельные расчеты показывают, что там, где снежный покров сходит рано, влажность почвы начнет снижаться уже весной, а с наступлением лета это уменьшение становится заметным [1].
В наземных экосистемах основным источником естественного поступления углерода в атмосферу считается дыхание почвы, определяемое активностью микроорганизмов, грибов и дыханием корней. Эмиссионная составляющая углеродного цикла, или общее почвенное дыхание, представляет собой показатель, который не только позволяет дать интегральную оценку функционирования микробного сообщества почвы и экосистемы в целом, но и весьма чутко реагирует на любые изменения гидротермического режима почв, отражающего современные климатические условия в регионе [2].
Зависимость дыхания почвы от ее влажности исследовали Е. А. Давидсон и др. [3]. По их данным, скорость дыхания начинает уменьшаться, когда водный матричный потенциал почвы становится ниже -0,5 МПа. При изменении водного потенциала почвы от -0,5 до -1,5 МПа дыхание почвы снижается на 50%, а при потенциале -3,0 МПа – до 20%.
В то же время дыхание надземной нефотосинтезирующей части биомассы также играет существенную роль в углеродном балансе природных экосистем. Вклад эмиссии СО2 с поверхности стволов (дыхание стволов) в общую эмиссию надземной фитомассы лесов составляет от 5 до 22% [3–5]. В Восточной
Финляндии [6] изучены суточные изменения дыхания ствола сосны в разные промежутки вегетационного периода. Осенью интенсивность дыхания в течение суток мало изменялась и составляла около 0,3 мкмоль СО 2 /(м2с). Летом этот показатель ночью достигал 0,8–1,1, а днем – 1,0–1,7 мкмоль СО2/(м2с), т.е. в среднем около 1,2 мкмоль СО2/(м2с) (1,9 мг СО2/(дм2ч)). Получены взаимосвязи между дыханием ствола в расчете на единицу поверхности и температурой ствола, а также между уровнем дыхания стволов в насаждении и гросс-продукцией (GPP) экосистемы. На уровне древостоя значение дыхания стволов за сутки составляло 0,1–0,5 г С/(м2сут) (1,8 г СО 2 /(м2с)) земной поверхности, а годичное - около 75 г С/м2 (275 г СО 2 /м2) земной поверхности, т.е. около 8% GPP. В Словакии в насаждении ели европейской измерены сравнительные потоки эмиссии СО2 из почвы, ствола и ветвей, которые равны 448, 56 и 70 ммоль СО 2 /(м2с) соответственно [7]. Соотношение дыхания почвы и корней исследовали в сложном ельнике [8], дыхание корней составило 46%.
Эмиссия с поверхности ствола, как и интенсивность фотосинтеза, зависит от водообе-спеченности растений, которую наиболее точно отражает предрассветный водный потенциал хвои – ПВПХ. Этот показатель характеризует водообеспеченность растений [9] независимо от влияния факторов окружающей среды. Так, снижение интенсивности фотосинтеза у пихты начинается при предрассветном водном потенциале хвои, равном 0,9–1,1 МПа, а достижение компенсационной точки у пихты бальзамической происходит при ПВПХ = -2,2 МПа, у пихты великой – -4,2 МПа [10].
Цель наших исследований – установить влияние условий окружающей среды на физиологические показатели сосны обыкновенной и ели обыкновенной.
Методика и объекты исследования
Исследования проводили на участке площадью 1 км2 между Московской кольцевой дорогой
(МКАД) и Рублевским шоссе. Наши предыдущие исследования [11] показали, что состояние древостоя на этом участке практически не отличается от состояния сосняка, произрастающего в Ярославской обл. Угличского района. Таким образом, мы считаем, что наши исследования проводятся на незагрязненном участке. Измерения интенсивности фотосинтеза и дыхания стволов (с середины мая до середины июля) проводили в 140-летнем сосняке разнотравно-черничном I класса бонитета и в произрастающей рядом куртине 60-летней ели. На этом участке почва дерново-слабоподзолистая супесчаная, уровень почвенно-грунтовых вод – 2,0–2,5 м. Для исследований использовали 140-летнее дерево сосны обыкновенной ( Pinus sylvestris L.) I класса роста, высотой 27 м и диаметром 45 см и дерево 60-летней ели обыкновенной ( Picea abies L.) I класса роста, высотой 22 м и диаметром 30 см. Интенсивность фотосинтеза сосны измеряли в середине кроны на высоте 22 м, у ели – в верхней части кроны на высоте 18 м.
Измерение эмиссии СО2 с поверхности стволов и интенсивности фотосинтеза проводили с помощью метода экспозиционных камер по открытой схеме. Такую схему многие исследователи используют для оценки эмиссии СО2 из почвы [12–15]. На высоте около 1,3 м северной стороны ствола дерева устанавливали камеры с площадью основания 200–250 см2 и объемом 400–750 см3 из прозрачной полиэтиленовой пленки для защиты от перегрева при попадании на их поверхность прямых солнечных лучей [11]. Крепление камеры осуществляли с помощью пластилина толщиной 0,8–1,0 см. Входные и выходные отверстия для шлангов располагали в обрамлении камеры с разных сторон, что обеспечивало равномерное перемешивание воздуха внутри камеры. Положение камеры на дереве не менялось на протяжении всего периода наблюдений. В ходе эксперимента поддерживался постоянный проток окружающего наружного воздуха через камеры со средней скоростью 1,0 л/мин. Эмиссию СО2 с поверхности ствола рассчитывали как функцию разности концентраций СО2 между выходящим из камеры и входящим в нее наружным воздухом, скорости воздушного потока и площади поверхности камеры, прилегающей к стволу [11, 16].
Камеры для определения фотосинтеза тоже изготовлены из полиэтиленовой пленки. Для отбора воздуха из камеры в противоположный конец от входа вводили полиэтиленовый шланг. Измерения и корректировку скорости потока воздуха через камеры проводили с помощью поплавкового расходомера РС-3А. Побудителями расхода воздуха служили мембранные микрокомпрессоры (Sonic-388 Китай). Концентрацию СО2 измеряли с помощью портативного инфракрасного газоанализатора LI-820 (Li-Cor, США). Показания газоанализатора регистрировались каждые 5 с и сохранялись с 20-секундным осреднением в накопителе данных EMS (Чехия).
Автоматическая система переключения каналов обеспечивалась 3-ходовыми клапанами (D330PB32/D/111140, Италия), которые позволяют устройству осуществлять прокачку воздуха через камеры непрерывно в течение всего периода исследования, даже когда измерения газообмена не проводят.
Получение информации с камер происходило попеременно с использованием автоматического устройства по сбору данных газообмена растений. На устройстве можно проводить эксперимент на 5-ти объектах последовательно по алгоритму «опыт–контроль» с задаваемым интервалом от 15 до 35 мин на каждую камеру беспрерывно в течение несколько суток. Автоматическая система для сбора данных по газообмену разработана в Институте лесоведения РАН. Для оценки водообеспеченности и предрассветного водного потенциала хвои (ПВПХ) использовали камеру давления [16, 17].
Результаты и обсуждение
Полученные данные показывают, что среднесуточные значения интенсивности фотосинтеза и эмиссии СО2 с поверхности ствола (дыхание ствола) изменяются в зависимости от поступления солнечной радиации (рис. 1). Среднесуточные значения интенсивности фотосинтеза и дыхания ствола рассчитывались на единицу проекции поверхности хвои и единицу поверхности ствола соответственно.
В малооблачные дни при среднесуточной солнечной радиации, равной 90–150 Вт/м2, среднесуточная интенсивность фотосинтеза охвоенных побегов сосны составила 4 мкмоль СО 2 /(м2с), а в дни со сплошной облачностью (при среднесуточной солнечной радиации 30–60 Вт/м2) – только 1-3 мкмоль СО 2 /(м 2 с). Интенсивность фотосинтеза охвоенных побегов ели значительно ниже, чем у сосны: в малооблачные дни – 1,5–2 мкмоль СО 2 /(м 2 с), а в дни со сплошной облачностью -около 0,5 мкмоль СО 2 /(м2с) (рис. 1А).
Среднесуточное значение дыхания стволов также зависит от прихода солнечной радиации за сутки и в малооблачный день составляет около 3 мкмоль СО 2 /(м2с) (рис. 1Б). Различий в интенсивности дыхания ствола у сосны и ели, в отличие от фотосинтеза, практически не наблюдается.
Значения дыхания стволов ели, полученные в настоящем исследовании (см. рис. 1Б, 2Б), сопоставимы с результатами других исследований [18]. В Тверской обл. в июне интенсивность дыхания стволов деревьев ели I класса роста составляла 5,0, III класса роста – 2,8, V класса – 1,6 мкмоль СО2/(м2с). Полученные значения дыхания стволов 25-летних елей летом в Московской обл. изменялись в пределах 0,04-0,2 мг СО2/(м2с) (или 0,9-4,5 мкмоль СО2/(м2с)), что согласуется с нашими результатами [19].
Таким образом, как показали другие авторы и наши предыдущие исследования, интенсивность дыхания ствола сосны и ели изменяется в течение дня в зависимости от поступления в крону солнечной радиации. У деревьев разного класса роста интенсивность эмиссии отличается.
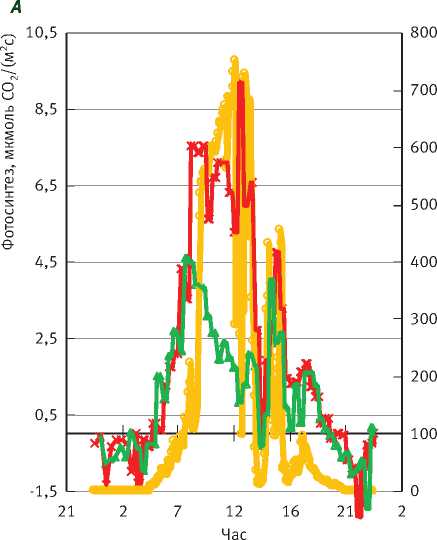
В Московской обл. на супесчаных почвах во второй половине лета возможен дефицит влаги. Обычно в таких условиях предрассветный водный потенциал хвои (ПВПХ) снижался до -0.8 МПа (исключение – 2010 г). Рассмотрим суточный ход газообмена хвои сосны и ели в условиях недостатка влаги. В малооблачные дни изменение дневного суточного хода в основном проявляется в снижении интенсивности фотосинтеза у обеих пород в первой половине дня (рис. 2А), при этом у ели депрессия фотосинтеза наблюдается несколько раньше. Сосна по сравнению с елью
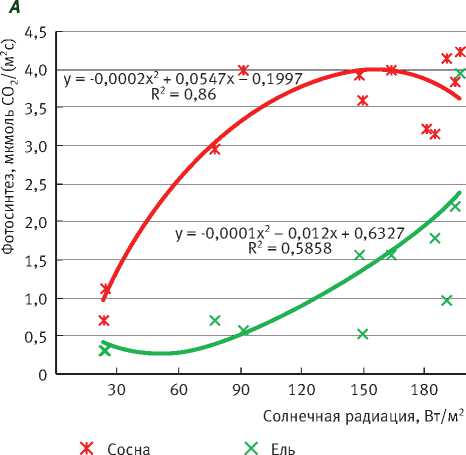
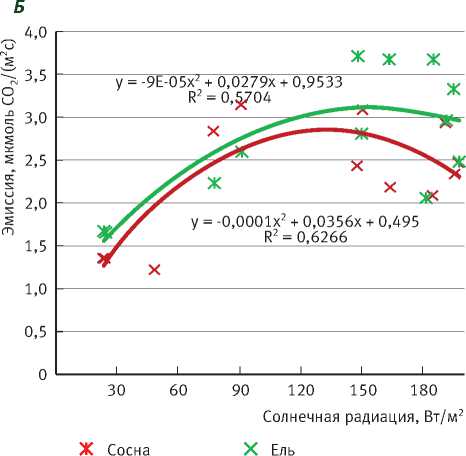
Рис. 1. Зависимость среднесуточных значений газообмена СО2 сосны и ели от средних за сутки значений солнечной радиации в дни с разной облачностью:
А – фотосинтез охвоенных побегов, Б – эмиссия СО2 с поверхности стволов
Фотосинтез сосны Фотосинтез ели Радиация
Ствол сосны
Ствол ели
Радиация
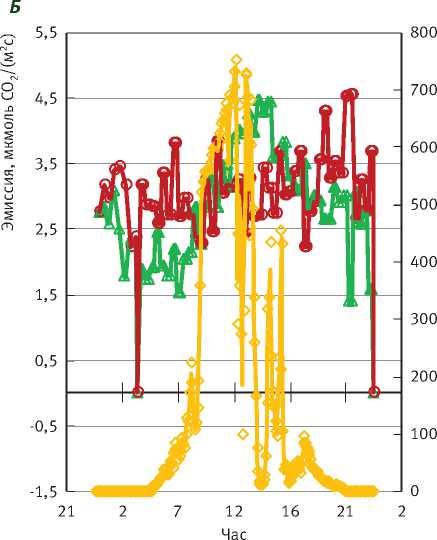
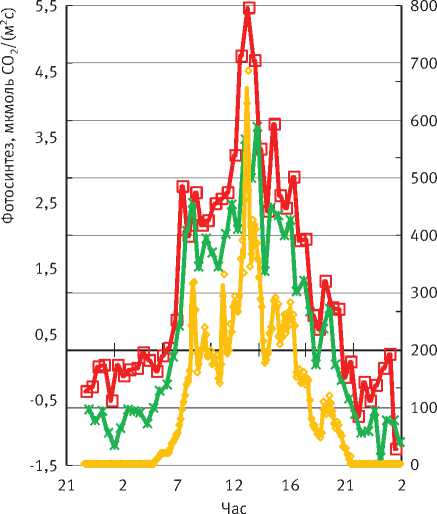
Рис. 2. Суточный ход газообмена СО2 сосны и ели и солнечной радиации в малооблачный день:
А – фотосинтез охвоенных побегов, Б – эмиссия СО2 с поверхности стволов более устойчива к недостатку влаги: она сохраняет максимальную интенсивность фотосинтеза 5 ч (с 8 до 13 ч), а ель – только 2 ч (с 7 до 9 ч). Однако во второй половине дня интенсивность фотосинтеза обеих пород становится одинаковой.
Таким образом, при ПВПХ, равном -0,8 МПа, и у сосны, и у ели в первой половине дня наблюдается депрессия фотосинтеза, т.е. такой недостаток влаги уже приводит к значительному снижению суточных значений фотосинтеза. Наши предыдущие исследования в юго-восточной лесостепи [15] также показали, что снижение водообеспеченности дуба в первую очередь влияет на длительность максимального фотосинтеза в течение суток. Чем больше дефицит влаги, тем раньше в течение дня наступает депрессия фотосинтеза.
Дыхание стволов в дни, когда наблюдалась депрессия фотосинтеза (рис. 2Б), было практически одинаковым и в среднем за сутки составляло у сосны 3,1, у ели - 2,8 мкмоль СО2/(м2с). При этом с поступлением солнечной радиации более высокой интенсивности дыхание стволов у обеих пород увеличивается. Это, скорее всего, связано с большим значением оттока из кроны полученных ассимилятов и более высокой транспирацией. Тем не менее при солнечной радиации 150 Вт/м2 за сутки у ели дыхание начинает снижаться, что, скорее всего, связано с депрессией фотосинтеза ели в малооблачные дни.
В облачный день при таком же недостатке влаги суточный ход фотосинтеза сосны и ели различается значительно меньше и изменяется синхронно (рис. 3). Интенсивность дыхания в пасмурные дни в 2 раза слабее, чем в малооблачные.
На рис. 4 представлены световые кривые фотосинтеза сосны и ели в малооблачные и
Фотосинтез сосны
Фотосинтез ели
Радиация
Рис. 3. Суточный ход фотосинтеза охвоенных побегов сосны и ели и солнечной радиации облачные дни. В пасмурные дни эти кривые различаются мало, только при солнечной радиации свыше 100 Вт/м2 значения фотосинтеза начинают существенно отличаться: у ели – 3,5 мкмоль СО2/(м2с), а у сосны - 5 мкмоль СО2/(м2с).
В малооблачные дни различие световых кривых у сосны и ели выражается значительно сильнее. С увеличением поступления солнечной радиации они начинают постепенно отдаляться друг от друга, и при повышении ее значений до 400–600 Вт/м2 интенсивность фотосинтеза этих пород различается более чем в 2 раза.
При сравнении газообмена сосны и ели установлено, что интенсивность фотосинтеза (в расчете на единицу площади поверхности хвои) у сосны примерно в 2–3 раза выше, чем у ели. Однако, как известно, лесоводственная продуктивность древостоев этих видов практически одинакова. При расчете на площадь произрастания различия в интенсивности фотосинтеза будут незначительные, так как масса хвои ели в древостое значительно больше массы хвои сосны на единицу площади произрастания.
В этой связи можно было бы предположить, что дыхание с поверхности стволов должно быть одинаковым (поверхность стволов в древостоях примерно равна). Однако наши данные показали, что весной интенсивность эмиссии СО2 с поверхности стволов ели в среднем на 50% выше, чем с поверхности стволов сосны.
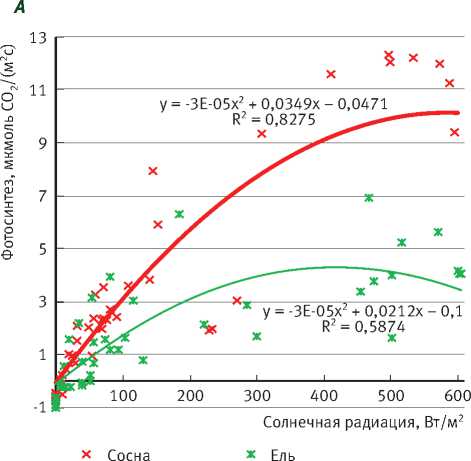
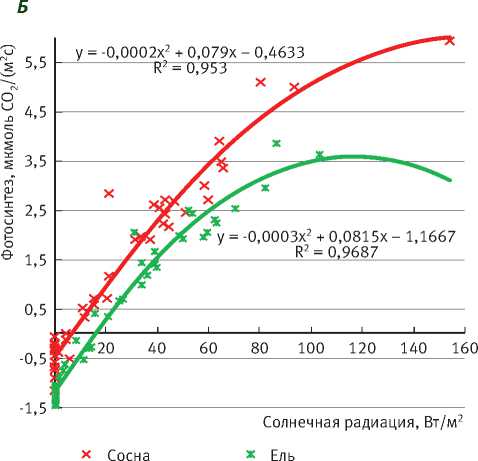
Рис. 4. Зависимость интенсивности фотосинтеза сосны и ели от солнечной радиации вмалооблачные (А) и облачные (Б) дни
Заключение
Исследования показали, что в условиях Московской обл. суточные изменения эмиссии с поверхности стволов ели обыкновенной и сосны обыкновенной одинаковы, хотя интенсивность фотосинтеза, рассчитанная на единицу площади проекции поверхности хвои, сильно различается.
При недостатке влаги интенсивность фотосинтеза ели снижается в большей степени, чем сосны. Так, в малооблачный день при ПВПХ, равном -0,8 МПа, интенсивность фотосинтеза ели снижается уже с 9 ч, а у сосны – только после 12 ч. В облачные дни, при таком же водообеспечении, практически в течение всего дня интенсивность фотосинтеза ели не снижается, и различия в световых кривых фотосинтеза сосны и ели менее значимы.
Дыхание стволов сосны и ели при расчете за сутки зависит от поступления солнечной радиации: в пасмурные дни интенсивность дыхания в 2 раза слабее, чем в малооблачные дни.
Наиболее информативным показателем, отражающим состояние древостоев, является предрассветный водный потенциал листа (хвои). Этот показатель характеризует водообеспечен-ность дерева независимо от причины, влияющей на его снижение. Дневной ход интенсивности фотосинтеза и эмиссия СО2 с поверхности стволов также являются информативными показателями состояния деревьев. Однако при сравнении двух разных по теневыносливости пород целесообразнее пользоваться показателем дыхания с поверхности стволов, так как поверхность хвои в древостоях в расчете на единицу площади произрастания значительно различается.
Список литературы Сравнение экофизиологических показателей сосны и ели в Серебряноборском опытном лесничестве
- Мелешко, В. П. Потепление климата: причины и последствия / В. П. Мелешко // Химия и жизнь. - № 4. - 2007. - С. 6-11.
- Effect of the Observed Climate Changes and Extreme Weather Phenomena on the Emission Component of the Carbon Cycle in Different Ecosystems of the Southern Taiga Zone / I. N. Kurganova, V. O. Lopes de Gerenyu, A. S. Petrov, T. N. Myakshina, D. V. Sapronov, V. A. Ableeva., V. N. Kudeyarov // Doklady Biol. Sci. - 2011. - V. 441. - P. 412-416.
- Davidsоn, E. A. Soil water content and temperature as independent or confounded factors controlling soil respiration in a temperate mixed hardwood forest / E. A. Davidsоn, E. Belk, R. D. Boone // Global Change Biology. - 1998. - V.4. - P. 217-227.
- Measurements of carbon sequestration by long-term eddy covariance: methods and a critical evaluation of accuracy / M. L. Goulden, J. W. Munger, S.-M. Fan, B. C. Daube, S. C. Wofsy // Global Change Biol. - 1996. - V. 2. - P. 169-182.
- Lavigne, M. B. Comparing nocturnal eddy covariance measurements to estimates of ecosystem respiration made by scaling chamber measurements at six coniferous boreal sites / M. B. Lavigne, M. G. Ryan, L. Anderson // Geophys. Res. -1997. - V. 102. - N. 28. - P. 977- 985.
- Law, B. E. Seasonal and annual respiration of a ponderosa pine ecosystem / B. E. Law, M. G. Ryan, P. M. Anthoni // Global Change Biol. - 1999. - V. 5. - P. 169-182.
- Seasonal and Annual Stem Respiration of Scots Pine Trees under Boreal Conditions / T. Zha, S. Kellomaki, Wang, A. Ryyppo, S. Ninisto // Annals of Botany. - 2004. - V. 94. - № 6. - P. 889-896.
- Brossaud, J. Field measurements of carbon dioxide efflux from soil and woody tissues in Norway spruce forest stand / J. Brossaud, M. V. Marek // Ecologia (Bratislava). - 2000. - V. 19. - № 3. - P. 245-250.
- Молчанов, А. Г. Предрассветный водный потенциал как показатель влагообеспеченности древостоев / А. Г. Молчанов // Проблемы экологического мониторинга и моделирования экосистем. - Т. XXXIX. - № 3. - 2018. - С. 79-92.
- Puritch, G. R. Effect of photosynthesis, respirations and transpiration of four Abies species / G. R. Puritch // Canad. J. Forest Res. - 1973. - V. 3. - № 3. - Р. 293-298.
- Молчанов, А. Г. Мониторинг эколого-физиологических показателей в экосистемах. Гл. 7 / А. Г. Молчанов // Серебряноборское опытное лесничество: 65 лет лесного мониторинга. - М.: КМК, 2010. - С. 112-130.
- Edwards, N. N. Continuous measurement of carbon dioxide evolution from partitioned forest floor components / N. N. Edwards, P. Sollins // Ecology. - 1973. - V. 54(2). -Р. 406-412.
- Maseyk, K. S. Photosynthetic responses of New Zealand Sphagnum species / K. S. Maseyk, T. G. A. Green // New Zealand Journal of Botany. - 1999. - V. 37. - P. 155-165.
- Measurements of carbon dioxide fluxes by chamber method at the Rzecin Wetland ecosystem, Poland / B. Chojnicki, M. Michalak, M. Acosta, R. Juszczak, J. Augustin, M. Drösler, J. Olejnik // Polish J. of Environ. Stud. - 2010. - V. 19. - № 2. - P. 283-291.
- Молчанов, А. Г. Баланс СО2 в экосистемах сосняков и дубрав в разных лесорастительных зонах / А. Г. Молчанов. - Тула: Гриф и К., 2007. - 284 с.
- Sap pressure in vascular plants. Negative hydrostatic pressure can be measured in plants / P. F. Scholander, H. T. Hammel, E. D. Bradstreet, E. A. Hemmingsen // Science. - 1965. - V. 148. - P. 339-346.
- Рахи, М. О. Аппаратура для исследований компонентов водного потенциала листьев / М. О. Рахи // Физиология растений. - 1973. - Т. 20. - С. 215-221.
- Молчанов, А. Г. Эмиссия СО2 стволами живых деревьев и валежом в еловых лесах юга-запада Валдайской возвышенности / А. Г. Молчанов, Ф. А. Татаринов, Ю. А. Курбатова // Лесоведение. - 2011. - № 3. - С. 14-25.
- Рост и газообмен СО2 у лесных деревьев / Ю. Л. Цельникер, И. С. Малкина, А. Г. Ковалев, С. Н. Чмора, В. В. Мамаев, А. Г. Молчанов. - М.: Наука, 1993. - 256 с.
