Сравнительная характеристика кариометрических параметров микроспороцитов и клеток тапетума сформированных пыльников длинно- и короткостолбчатых растений Jasminum fruticans L. (Oleaceae)
")
Автор: Кузьмина Т.Н.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Ботаника
Статья в выпуске: 1, 2024 года.
Бесплатный доступ
В связи с изучением функциональной активности генеративных структур при гетеростилии проведен сравнительный кариометрический анализ клеток тапетума, а также микроспороцитов сформированных пыльников длинно- и короткостолбчатых растений Jasminum fruticans L. (Oleaceae). На основании измерения площадей ядра, ядрышка, подсчета количества ядер и ядрышек в клетках вычисляли суммарные значения площадей ядерных структур. Поскольку с ядрышками связаны процессы транскрипции и трансформации рибосомальной РНК (р-РНК), то состояние ядрышковых структур в клетке отражает уровень ее функциональной активности, поэтому в качестве характеристики функциональной активности клеток использовали значение ядерно-ядрышкового отношения (ЯЯО). Установлено, что у обеих флоральных форм в сформированном пыльнике преобладают двуядерные тапетальные клетки. Число ядрышек, приходящихся на клетку, варьирует от 1 до 14. У короткостолбчатых растений число тапетальных клеток с 6-8 ядрышками больше по сравнению с длинностолбчатыми растениями. Однако данный факт не отражается на значении ядерно-ядрышкового отношения. По функциональной активности клетки тапетума пыльников длинно- и короткостолбчатых растений не имеют статистически значимого различия. Сравнение кариометрических параметров микроспороцитов показало, что площади ядерных структур больше у длинностолбчатых растений, при этом они характеризуются более высоким значением ЯЯО (17.61±0.36) по сравнению с короткостолбчатыми растениями, у которых ЯЯО составляет 15.59±0.27. Понижение значения ЯЯО у короткостолбчатых растений является признаком усиления интенсивности белкового метаболизма их микроспороцитов, а следовательно, уровня функциональной активности их геномов. Различие функциональной активности микроспороцитов длинно- и короткостолбчатых растений может быть признаком проявления пола в зависимости от флорального морфологического типа растения.
Андроцей, гетеростилия, микроспорангий, пыльник, спорогенные клетки, стенка пыльника, ядерно-ядрышковое отношение, ядро, ядрышко, jasminum fruticans, oleaceae
Короткий адрес: https://sciup.org/147243405
IDR: 147243405 | УДК: 633.811:582.931:581.182.1:611.018.13 | DOI: 10.17072/1994-9952-2024-1-22-31
Comparative characteristics of karyometric parameters of microsporocytes and tapetum cells of the formed anthers of long-styled and short-styled plants Jasminum fruticans L. (Oleaceae)
In connection with the study of the functional activity of generative structures in heterostylous plants, a comparative karyometric analysis of tapetum cells, as well as microsporocytes of the formed anthers of long-styled and short-styled plants Jasminum fruticans L. (Oleaceae) was carried out. Based on the measurement of the nucleus areas, nucleolus, counting the number of nuclei and nucleoli in cells, the total values of the areas of nuclear structures were calculated. Since the processes of transcription and transformation of ribosomal RNA (r-RNA) are associated with nucleoli, the state of nucleolar structures in a cell reflects the level of its functional activity, therefore, the value of the nuclear-nucleolar ratio (NNR) was used as a characteristic of the functional activity of cells. It was found that in both floral forms, binuclear tapetal cells predominate. The number of nucleoli per cell varies from 1 to 14. The number of tapetal cells with 6-8 nucleoli is greater in short-styled plants than in long-styled plants. However, the cells of tapetum of long-styled and short-styled plants do not have a statistically significant difference of the value of NNR. Comparison of karyometric parameters of microsporocytes showed that the areas of their nuclear structures are larger in long-styled plants, while they are characterized by a higher value of NNR (17.61±0.36) compared with short-styled plants (15.59±0.27). A decrease in the value of NNR in short-styled plants is a sign of an increase in the intensity of protein metabolism of their microsporocytes, and, consequently, the level of functional activity of their genomes. Differences in the functional activity of microsporocytes of long-styled and short-styled plants can be considered as a sign of sex manifestation of sex depending on the floral type of the plant.
Текст научной статьи Сравнительная характеристика кариометрических параметров микроспороцитов и клеток тапетума сформированных пыльников длинно- и короткостолбчатых растений Jasminum fruticans L. (Oleaceae)
Гетеростилия покрытосеменных растений – это генетически контролируемый полиморфизм цветков, который связан с системой несовместимости. Явление гетеростилии, или разностолбчатости цветков, отмечено у представителей 36 семейств покрытосеменных растений [Агаджанян, 2000; Ganders 1979]. В зависимости от особенностей строения андроцея и гинецея цветка в пределах вида выделяют, как правило, две флоральные, или морфологические, формы. Цветки различных морфологических форм дифференцируются по высоте столбика пестика, форме рыльца пестика, размерам тычинок, пыльников, пыльцевых зерен [Pailler, Thompson, 1997; Tompson, Dommée, 2000; Klein, Freitas, Da Cunha, 2009; Valois-Cuesta, Soriano, Ornelas, 2011; Keller, de Vos, Conti, 2012]. Ключевым вопросом в изучении гетеростилии является выявление функционального значения полиморфизма цветков. Наиболее распространенным объяснением его биологической роли является свойственная гетеростильным видам гетероморфная несовместимость, при которой эффективным оказывается переопыление цветков, относящихся к различным морфологическим типам, т.е. когда пыльца из короткостолбчатых цветков попадает на рыльца длинностолбчатых цветков или наоборот. Как правило, у гетеростильных видов более эффективным является опыление, которое происходит между морфами, а опыление цветков внутри одной морфлогиче-ской формы показывает слабую завязываемость семян [Stevens, Murray, 1982; Valois-Cuesta, Soriano, Ornelas, 2011; Wu et al., 2015; Gao et al., 2021]. H.G. Baker было высказано предположение, что гетеростиль-ные виды можно рассматривать как переходную форму от гермафродитных видов к однополым растениям [Baker, 1960; Жинкина, 2000]. В частности, у Mussaenda pubescens (Rubiaceae) отмечены случаи редукции пыльников у длинностолбчатых цветков, и семязачатков – у короткостолбчатых [Li et al., 2010]. Проведенные ранее исследования генезиса пыльников и семязачатков дистильного вида Jasminum fruti-cans L. (Oleaceae) не выявили различий в ходе генезиса пыльников и семязачатков, а также редукции генеративных элементов коротко- и длинностолбчатых растений [Кузьмина, 2018, 2000; Шевченко, Плугатарь, 2019]. Однако флоральные формы J. fruticans различаются по долям морфологически нормальных пыльцевых зерен [Кузьмина, 2017]. Различия по фертильности пыльцевых зерен длинно- и короткостолбчатых растений указаны и для других гетеростильных видов ( Limonium caspium (Willd.) Gams., L. gmelinii (Willd.) O. Kuntzw, Goniolimon elatum (Fisch. ex Spreng.) Boiss . (Plumbaginaceae) [Демьянова, 2014], что может свидетельствовать о различной функциональной активности генеративных элементов флоральных форм гетеростильных видов.
Известно, что стенка гнезда (микроспорангия) сформированного пыльника состоит из нескольких слоев клеток, представленных эпидермисом, эндотецием, средним слоем и тапетумом, окружающих клетки спорогенной ткани [Батыгина, Васильева, 2002]. Наиболее функционально важными тканями микроспорангия являются спорогенная ткань и тапетум. Развитие спорогенной ткани приводит к формированию микроспороцитов – материнских клеток пыльцевых зерен. Tапетум как внутренний слой стенки микроспорангия, клетки которого непосредственно прилегают к спорогенной ткани, рассматривается как наиболее активная и чувствительная к стрессовым воздействиям соматическая ткань пыльника, которая в этот период функционирует как проводящая, обеспечивающая поступление питательных веществ к микроспороцитам [Резникова, 1984; Gothandam, Kim, Chung, 2007; Yeung et al., 2011]. Известно, что нарушения микроспорогенеза и формирования гаметофита сопряжены с морфоструктурными изменениями клеток тапетума, в которых изменения затрагивают главным образом эндоплазматический ретикулум [Vi- jayaraghavan, Ratnaparkhi, 1979; Gothandam, Kim, Chung, 2007; Shi et al., 2010; Du et al., 2019].
Для определения интенсивности функционирования клеточных структур служат ядерные и ядрышковые характеристики клеток, поскольку они отражают интенсивность белкового метаболизма клетки, а следовательно, дают представление о функциональной активности генома клетки [Челидзе, Зацепина, 1988; Архип-чук, 1995]. Определение числа ядрышек и их размеров позволяет дать оценку активности генов рибосомной РНК (рРНК) на цитологическом уровне [Архипчук, 1995], что служит основой для биотестирования и определения цитогенетической стабильности видов при воздействии различных внешних факторов [Дуброва, 1989; Архипчук, 1995; Владимирова, Муратова, 2005; Калаев и др., 2009]. Кариометрические исследования репродуктивных структур высших растений [Резникова, Бугара, 1979; Романова, Третьякова, 2005; Трочин-ская, Бланковская, Тоцкий, 2010] не затрагивают вопросы гетеростилии видов.
Цель работы – сравнительная кариометрическая характеристика микроспороцитов и тапетальных клеток у сформированных пыльников длинно- и короткостолбчатых растений J. fruticans в связи с определением уровня их функционального состояния у растений различных флоральных (морфологических) форм.
Материалы и методы исследования
Jasminum fruticans L. (жасмин кустарниковый) – дистильный вид флоры стран Средиземноморья, Западной Европы, Малой Азии, Черноморского побережья Кавказа, а также Южного берега Крыма [Голубев, 1996]. Для исследования пыльников длинно- и коротостолбчатых растений J. fruticans брали терминальные бутоны длиной 1.5–2 мм. Сбор материала проводили на территории Никитского ботанического сада (г. Ялта, Республика Крым; 44°30ʹ34ʺс.ш. 34°13ʹ58ʺ в.д.) в 2019–2021 гг. Для приготовления постоянных препаратов бутоны фиксировали в смеси F. A. A. (formalin : acetic acid : alcohol 70%). Материал обезвоживали с помощью изопропилового спирта, после чего делали постоянные препараты бутонов. Препараты окрашивали гематоксилином и алциановым синим, учитывая, что гематоксилин обеспечивает визуализацию базофильных структур, к которым относятся ядра и РНК-содержащие структуры [Коржев-ский, 2007], а алциановый синий дает интенсивную окраску клеточных стенок, что позволяет четко идентифицировать границы клеток. Анализ препаратов проводили с помощью микроскопа AxioScope A.1 (Carl Zeiss, Germany) и цифровой камеры AxioCamERc5s (Carl Zeiss, Germany). Для каждой флораль-ной формы на поперечных срезах пыльников делали измерения клеток 30 пыльников. Кариометрические показатели получены с помощью программы AxioVision Rel. 4.8.2. (Carl Zeiss, Германия). Основными параметрами измерений были площадь клетки, площадь ядра и площадь ядрышка. В случае многоядерных и многоядрышковых структур определяли их суммарную площадь. Кроме того, подсчитывали количество ядер и ядрышек, приходящихся на клетку. Ядерно-ядрышковое отношение (ЯЯО) определяли как отношение площади ядра к площади ядрышка, в случае многоядерных и многоядрышковых клеток вычисления проводили, используя суммарные показатели.
Вычисления и статистическую обработку данных делали в программном приложении Statistica 10.0 (StatSoft. Ins., USA). Определение статистически значимых различий проводили на основании t-критерия Стьюдента и критерия Фишера (F-критерий) при уровне статистической значимости p < 0.05. Сравнение выборочных долей клеток по числу ядер осуществляли с помощью критерия χ2 Пирсона [Гашев, Бетляе-ва, Лупинос, 2018]. Кариометрические данные представлены в виде: Х ± S x , где X – среднее арифметическое, S x – стандартная ошибка среднего.
Результаты исследования
Развитие стенки микроспорангия J. fruticans идет в центробежном направлении, при котором деление клеток первичного париетального слоя приводит к формированию тапетума и вторичного париетального слоя. Деление последнего в свою очередь дает начало эндотецию и среднему слою. Таким образом, у J. fruticans тапетум является производным первичного париетального слоя, который в сформированном микроспорангии многослойный, секреторного типа (рис. 1). Установлено, что в тапетуме сформированных пыльников J. fruticans преобладают двуядерные клетки. Так, в тапетуме длинностолбчатых растений 73.7% клеток двуядерные, а у короткостолбчатых растений их доля составляет 69.7%, что не представляет статистически значимого различия (χ2 = 3.94; df = 1; p = 0.05).
Известно, что для многоядрышковых клеток наиболее информативным показателем оценки функциональной активности генетического аппарата считается число ядрышек в клетке [Архипчук, 1995]. Количество ядрышек в тапетальных клетках варьирует от 1 до 14, при этом преобладают двуядрышковые клетки, что объясняется наличием двух ядер в большинстве клеток данной ткани. Так, у длинностолбчатых растений количество таких клеток составляет 48.3%, а у короткостолбчатых – 46.4% (рис. 2). Однако встречаются и клетки, ядра которых содержат большее число ядрышек. Показано, что у короткостолбчатых растений доля клеток, содержащих 6–8 ядрышек, выше, чем у длинностолбчатых растений. Клетки, содержащие более 9 ядрышек, встречаются у обеих флоральных форм лишь в единичных случаях.
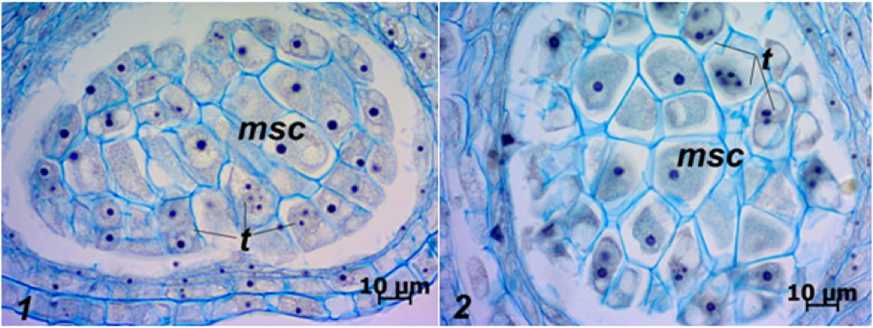
Рис. 1. Поперечные срезы микроспорангиев длинно- ( 1 ) и короткостолбчатых ( 2 ) растений J. fruticans : msc – микроспороциты; t – тапетум
[Cross section of microsporangium of long-styled ( 1 ) and short-styled ( 2 ) of J. fruticans : msc – microsporocytes; t – tapetum]
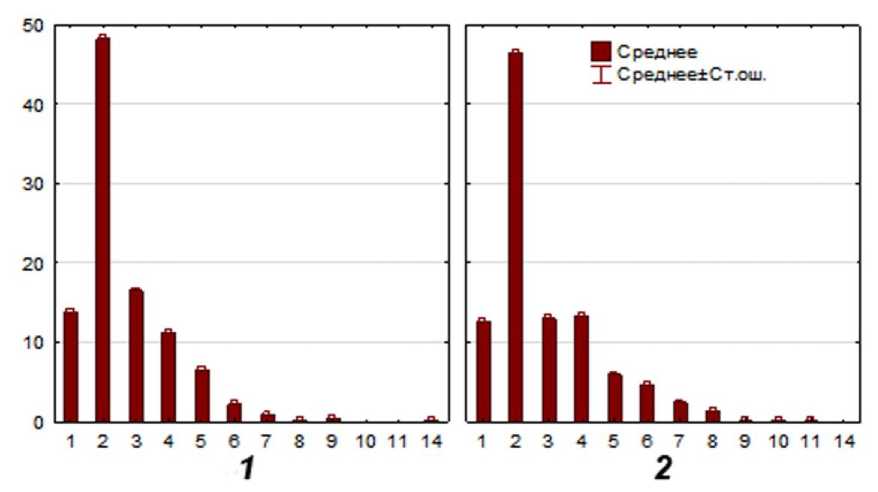
Рис. 2. Суммарное число ядрышек в клетках тапетума сформированных пыльников длинно- ( 1 ) и короткостолбчатых ( 2 ) растений J. fruticans .
Ось X – число ядрышек в ядре; Ось Y – процент клеток от общего числа проанализированных клеток
[The total number of nucleolus in tapetum cells of the formed anthers of long-styled ( 1 ) and short-styled ( 2) plants J. fruticans.
X axis – number of nucleoli in cell’s nuclear; Y axis – percentage of cells from the total number of cells analyzed]
Данные кариометрического анализа тапетальных клеток представлены в табл. 1. Клетки тапетума короткостолбчатых растений содержат более крупные ядра, суммарная площадь их ядрышек также оказывается больше по сравнению с длинностолбчатыми растениями. При этом, несмотря на статистически значимое различие клеток тапетума флоральных форм по площадям ядер и ядрышек, значения ЯЯО у них выражено слабо. Хотя можно отметить, что в сформированных пыльниках длинностолбчатых растений клетки тапетума по ЯЯО незначительно превышают аналогичный показатель короткостолбчатых. Анализ кариометрических параметров клеток тапетума сформированных пыльников длинно- и короткостолбчатых растений J. fruticans показал, что у обеих флоральных форм преобладают двуядерные клетки. В каждом ядре, как правило, содержится по одному ядрышку, а суммарное количество ядрышек на клетку равно двум. Таким образом, преобладают двуядрышковые тапетальные клетки. У короткостолбчатых растений, как упоминалось выше, чаще отмечаются клетки, содержащие более 5 ядрышек, что отражается на увеличении суммарной площади ядрышек, кроме того, они имеют более крупные ядра. Однако, в целом, это не оказывает статистически значимого влияния на показатели функциональной активности тапетальных клеток сравниваемых флоральных форм.
Таблица 1
Кариометрические параметры клеток тапетума сформированных пыльников длинностолбчатых и короткостолбчатых растений J. fruticans
[Karyometric parameters of tapetum cells of formed anthers of long-styled and short-styled plants J. fruticans ]
|
Параметр |
Длинно-cтолбчатые (n=1000) |
Короткостолбчатые (n=1000) |
t-критерий |
р-уровень |
|
Суммарная площадь ядер, мкм2 |
83.25±1.13 |
90.16±1.34 |
-3.95 |
0.001** |
|
Суммарная площадь ядрышек в клетке, мкм2 |
8.36±0.14 |
9.01±0.12 |
-3.46 |
0.001** |
|
Ядерно-ядрышковое отношение (ЯЯО) |
10.98±0.14 |
10.67±0.14 |
1.54 |
0.12ns |
Примечание. n – количество проанализированных клеток; ns – отсутствует статистически значимое различие при уровне статистической значимости p < 0.05; ** – различие статистически значимо при p < 0.05.
Микроспороциты у J. fruticans в гнезде пыльника расположены двумя–четырьмя рядами (см. рис. 1). Это крупные клетки, площадь которых варьирует в зависимости от морфотипа растения. Площадь поперечного сечения микроспороцитов в пыльниках у длинностолбчатых растений составляет 351.52±4.20 мкм2, а у короткостолбчатых – 330.27±3.92 мкм2. Микроспороциты J. fruticans содержат одно крупное ядро, занимающее центральное положение, в котором четко выражено одно ядрышко. Лишь в редких случаях отмечается образование двух–трех ядрышек. Для клеток с небольшим количеством ядрышек информативной характеристикой их функциональной активности является размер одиночных ядрышек [Архипчук, 1995]. Однако этот показатель относителен, поскольку размеры клеток и ядер у флоральных форм также различны (табл. 2).
Таблица 2
Кариометрические параметры микроспороцитов длинностолбчатых и короткостолбчатых растений J. fruticans
[Karyometric parameters of tapetum cells of formed anthers of long-styled and short-styled plants J. fruticans ]
|
Показатель |
Длинно-cтолбчатые (n=600) |
Короткостолбчатые (n=600) |
t-критерий |
р-уровень |
|
Площадь ядра, мкм2 |
134.50± 2.12 |
112.26± 1.53 |
8.52 |
0.001** |
|
Площадь ядрышка в клетке, мкм2 |
8.36± 0.11 |
7.72± 0.09 |
4.41 |
0.001** |
|
Ядерно-ядрышковое отношение (ЯЯО) |
17.61± 0.36 |
15.59± 0.27 |
4.52 |
0.001** |
Примечание. ** – различие статистически значимо при p < 0.05.
Поэтому наиболее объективную картину функциональной активности клетки демонстрирует ядерно-ядрышковое отношение (ЯЯО), свидетельствующее об интенсивности белкового метаболизма клетки [Машкин, Назарова, 1976; Хроленко и др., 2011]. У микроспороцитов короткостолбчатых растений ЯЯО меньше (15.59± 0.27) по сравнению с длинностолбчатыми растениями (17.61± 0.36).
Обсуждение результатов
Известно, что с увеличением размеров ядрышка в ядре снижается значение ЯЯО, что характерно для клеток, у которых наблюдается усиление активности белоксинтезирующей системы клетки. Следовательно, можно говорить, что у флоральных форм J. fruticans материнские клетки микроспор являются функционально неоднородными: бо́льшую интенсивность белкового метаболизма проявляют микроспоры пыльников короткостолбчатой формы. Как правило, увеличение активности ядрышковых организаторов хромосом, сопровождающееся снижением показателей ЯЯО, наблюдается при стрессовых повреждающих факторах (низких и высоких температурах. засухи. засоления. антропогенных воздействий и др.), что приводит к синтезу «стрессовых белков», являющихся ответной на цитологическом уровне адаптивной реакцией на стресс [Дуброва, 1989; Калаев и др., 2005; Вострикова, 2010; Калашник, 2013]. Таким образом, ядерно-ядрышковое отношение служит кариологическим признаком адаптивной устойчивости клеток. Как отмечалось выше, у дистильного вида J. fruticans микроспороциты короткостолбчатой формы имеют меньшее значение ЯЯО, что позволяет рассматривать их как более стабильную клеточную структуру по сравнению с аналогичными клетками длинностолбчатой формы. Характерно, что у длинностолбчатой формы дистильного вида Mussaenda pubescens (Rubiaceae) наблюдали процессы дегенерации пыльников [Li et al., 2010], что служит признаком функционального разделения полов в зависимости от морфологического типа растений. Процессы дегенерации генеративных элементов отмечены и у растений, отнесенных к группе потенциально обоеполых. Как правило, у этих растений в женских цветках деструктивные процессы в микроспорангиях проявляются в мейотический и постмейотический периоды, при этом на ранних этапах развитие и формирование клеточных слоев стенки микроспорангия, в том числе и тапетальной ткани, проходит без аномалий [Кордюм, Глущенко, 1976; Nugent et al., 2019]. В этом плане соотношение ядрышковой активности микроспороцитов и клеток тапетума пыльников длинно- и короткостолбчатых растений у J. fruticans может свидетельствовать о сходстве тенденции процессам развития пыльников, которые наблюдаются в пестичных и тычиночных цветках диэцичных растений с рудиментами органов другого пола. В частности, как пример переходной формы от гетеростилии к диэции, рассматривается Morinda parvifolia Bartl. ex DC. (Rubiaceae), у которого, несмотря на продуцирование морфологически одинаковых пыльцевых зерен растениями обеими флоральными формами, жизнеспособность пыльцы короткостолбчатых растений (S-форма) выше, чем у длинностолбчатых растений (L-форма) [Liu et al., 2012].
Заключение
В тапетуме сформированных пыльников у обеих флоральных форм J. fruticans преобладают двуядерные клетки, суммарное число ядрышек в которых равно двум. У короткостолбчатых растений по сравнению с длинностолбчатыми выше число клеток, в которых находится 6–8 ядрышек, что отражается на показателях суммарной площади ядрышек. Однако значение функциональной активности клеток тапетума длинно- и короткостолбчатых растений, оцениваемой как значения ядерно-ядрышкового отношения, не имеют статистически значимых различий.
Микроспороциты J. fruticans различаются по цитометрическим параметрам в зависимости от фло-ральной формы растения. Площади клеток и ядерных структур больше у длинностолбчатых растений.
Кариометрический анализ микроспороцитов показал статистически значимое снижение ядерно-ядрышкового отношения у короткостолбчатых растений по сравнению с длинностолбчатыми, что характеризует усиление функционирования их белоксинтезирующей системы. Данный факт можно рассматривать как проявление дифференциации функционирования мужской генеративной системы вида в зависимости от флорального (морфологического) типа растения
Список литературы Сравнительная характеристика кариометрических параметров микроспороцитов и клеток тапетума сформированных пыльников длинно- и короткостолбчатых растений Jasminum fruticans L. (Oleaceae)
- Агаджанян А.М. Распространённость и распределение гетеростилии в системе покрытосеменных растений // Успехи современной биологии. 2000. Т. 120, № 4. С. 348-360.
- Архипчук В.В. Использование ядрышковых характеристик в биотестировании // Цитология и генетика. 1995. Т. 29, № 3. C. 6-12.
- Батыгина Т.Б., Васильева В.Е. Размножение растений. СПб.: Изд-во СПб. ун-та, 2002. 232 с.
- Владимирова О.С. Муратова Е.Н. Кариологические особенности ели сибирской (Picea obovata Ledeb.) в условиях антропогенного загрязнения г. Красноярска // Экологическая генетика. 2005. Т. 3, № 1. С. 18-23.
- Вострикова Т.В. Эколого-физиологическая реакция семенного потомства древесных растений на стресс // Вестник Воронежского государственного университета. Сер. География. Геоэкология. 2010. № 1. С. 87-91.
- Гашев С.Н., Бетляева Ф.Х., Лупинос М.Ю. Математические методы в биологии. Анализ биологических данных в системе Statistica. М.: Юрайт, 2018. 208 с.
- Голубев В.Н. Биологическая флора Крыма. Ялта, 1996. 126 с.
- Демьянова Е.И. О полиморфизме некоторых гетеростильных растений лесостепного Зауралья // Вестник Пермского университета. Сер. Биология. 2014. Вып. 2. С. 10-17.
- Дуброва А.Н. Ядрышковые организаторы хромосом как адаптивный элемент вида // Журнал общей биологии. 1989. Т. 50, № 2. С.213-217.
- Жинкина Н.А. Гетеростилия // Эмбриология цветковых растений. Терминология и концепции. СПб.: Мир и семья. 2000. Т. 3. С. 82-83.
- Калаев В.Н. и др. Влияние низких активностей радона на ядерные структуры клеток апикальной меристемы корней Zebrina pendula Shcnizl (ядрышковый тест) // Вестник Воронежского государственного университета. Сер. География. Геоэкология. 2005. № 1. С. 80-85.
- Калаев В.Н. и др. Цитогенетические характеристики семенного потомства деревьев березы повислой в Хреновском бору // Лесоведение. 2009. № 3. С. 43-51.
- Калашник Н.А. Оценка функциональной активности ядрышковых организаторов хромосом у лиственницы Сукачева в условиях техногенного загрязнения // Вестник Оренбургского университета. 2013. 10 (159). С. 195-197.
- Кордюм Е.Л., Глущенко Г.И. Цитоэмбриологические аспекты проблемы пола покрытосеменных. Киев: Наукова думка, 1976. 200 с.
- Коржевский Д.Э. Применение гематоксилина в гистологической практике // Морфология. 2007. Т. 132, № 6. С. 77-82.
- Кузьмина Т.Н. Диморфизм цветков Jasminum fruticans L. // Вестник Пермского университета. Сер. Биология. 2017. Вып. 3. С. 233-239.
- Кузьмина Т.Н. Формирование мужских генеративных структур у Jasminum fruticans (Oleaecae) // Ботанический журнал. 2018. Т. 103, № 5. С. 654-663.
- Кузьмина Т.Н. Формирование семязачатка и семени Jasminum fruticans (Oleaceae) // Ботанический журнал. 2020. Т. 105, № 9. С. 919-930.
- Машкин С.И., Назарова М.Н. Сезонная динамика числа и размеров ядрышек. ядер и ядерно-ядрышковых отношений у представителей подсем. Сливовых при их интродукции // Цитология. 1976. Т. 18, № 12. С. 1438-1443.
- Резникова C.A. Цитология и физиология развивающегося пыльника. М.: Наука, 1984. 272 с.
- Резникова С.А., Бугара А.М. Функциональные изменения объема ядер в тканях развивающегося пыльника лилии // Цитология. 1979. Т. 21, № 6. С. 671-677.
- Романова Л.И., Третьякова И.Н. Особенности микроспорогенеза у лиственницы сибирской, растущей в условиях техногенного стресса // Онтогенез. 2005. Т. 36, № 2. С. 128-133.
- Трочинская Т.Г., Бланковская Т.Ф., Тоцкий В.М. Количественные признаки клеток мужских генеративных структур пшеницы, ржи и пшенично-ржаных гибридов в микроспорогенезе // Цитология и генетика. 2010. Т. 4. С. 48-54.
- Хроленко Ю.А. и др. Количественные характеристики ядрышек в клетках Panax ginseng in vivo и in vitro // Turczaninowia. 2011. T. 14, № 1. С. 104-108.
- Челидзе П.В., Зацепина О.В. Морфофункциональная классификация ядрышек // Успехи современной биологии. 1988. Т. 105, № 2. С. 252-258.
- Шевченко С.В., Плугатарь Ю.В. Исследования репродуктивной биологии семенных растений в Никитском ботаническом саду // Сборник научных трудов ГНБС. 2019. Т. 149. С. 177-198. URL: https://doi.org/10.36305/0201-7997-2019-149-177-198 (дата обращения: 19.07.2023).
- Baker H.G. Reproductive methods as factors in speciation in flowering plants // Cold Spring Harbor symp. Quant. Boil. 1960. Vol. 24. P. 177-191.
- Du K. et al. Abnormal tapetum development and energy metabolism associated with sterility in SaNa-1A CMS of Brassica napus L. // Plant Cell Rep. 2019. Vol. 38. P. 545-558. URL: https://doi.org/10.1007/s00299-019-02385-2 (дата обращения: 19.07.2023).
- Ganders F.R. The biology of heterostyly // New Zealand Journal of Botany. 1979. Vol. 17. P. 607-635. URL: https://doi.org/10.1080/0028825X.1979.10432574 (дата обращения: 19.07.2023).
- Gao S. et al. The nonreciprocal heterostyly and heterotypic self-incompatibility of Ceratostigma will-mottianum // J. Plant Res. 2021. Vol. 134. P. 543-557. URL: https://doi.org/10.1007/s10265-021-01269-5 (дата обращения: 19.07.2023).
- Gothandam K.M., Kim E.S., Chung Y.Y. Ultrastructural study of rice tapetum under low-temperature stress // J. Plant Biol. 2007. Vol. 50(4). P. 396-402. URL: https://doi.org/10.1007/BF03030674 (дата обращения: 19.07.2023).
- Keller B., de Vos J.M., Conti E. Decrease of sexual organ reciprocity between heterostulous primrose species. with possible functional and evolutionary implications // Annals of Botany. 2012. Vol. 110. P. 12331244. URL: https:// doi.org/10.1093/aob/mcs199 (дата обращения: 19.07.2023).
- Klein D.E., Freitas L., Da Cunha M. Self-incompatibility in a distylous species of Rubiaceae: is there a single incompatibility response of the morphs? // Sex Plant Reprod. 2009. Vol. 22. P. 121-131. DOI: 0.1007/s00497-009-0097-0. URL: https://pubmed.ncbi.nlm.nih.gov/20033433 (дата обращения: 19.07.2023).
- Li A.-M. et al. Cryptic dioecy in Mussaendapubescens (Rubiaceae): a species with stigma-height dimorphism // Annals of Botany. 2010. Vol. 106. P. 521-531. URL: https://doi.org/10.1093/aob/mcq146 (дата обращения: 19.07.2023).
- Liu Y. et al. Functional dioecy in Morinda parvifolia (Rubiaceae): a species with stigma-height dimorphism // Plant Systematics and Evolution. 2012. Vol. 298. P. 775-785. URL: https://doi.org/10.1007/s00606-011-0588-y (дата обращения: 19.07.2023).
- Nugent J.M. et al. Progressive programmed cell death inwards across the anther wall in male sterile flowers of the gynodioecious plant Plantago lanceolata // Planta. 2019. Vol. 249. P. 913-923. URL: https://doi.org/10.1007/s00425-018-3055-y (дата обращения: 19.07.2023).
- Pailler Th., Thompson J.D. Distyly and variation in heteromorphic incompatibility in Gaertnera vaginata (Rubiaceae) endemic to La Reunion Island // American Journal of Botany. 1997. Vol. 84(3). P. 315-327. URL: https://doi.org/10.2307/2446005 (дата обращения: 19.07.2023).
- Shi S. et al. A comparative light and electron microscopic analysis of microspore and tapetum development in fertile and cytoplasmic male sterile radish // Protoplasma. 2010. Vol. 241. P. 37-49. DOI: 10.1007/s00709-009-0100-5/ URL: https://link.springer.com/article/10.1007/s00709-009-0100-5 (дата обращения: 19.07.2023).
- Stevens V.A.M., Murray B.G. Studies on heteromorphic self-incompatibility systems: Physiological aspects of the incompatibility system of Primula obconica // Theoret. Appl. Genetics. 1982. Vol. 61. P. 245-256. DOI: 10.1007/BF00273782. URL: https://pubmed.ncbi.nlm.nih.gov/24270436/ (дата обращения: 19.07.2023).
- Tompson J.D., Dommee B. Morph-specific patterns of variation in stigma height in natural populations of distylous Jasminum fruticans // New Phytologist. 2000. Vol. 148. P. 303-314. URL: https://doi.org/10.1046/j.1469-8137.2000.00765.x (дата обращения: 19.07.2023).
- Valois-Cuesta H., Soriano P.J., Ornelas J.F. Dimorphisms and self-incompatibility in the distylous species Palicourea demissa (Rubiaceae): possible implications for its reproductive output // J. Plant Res. 2011. Vol. 124. P. 137-146. URL: https://doi.org/10.1007/s10265-010-0359-9 (дата обращения: 19.07.2023).
- Vijayaraghavan M.R., Ratnaparkhi S. Histological dynamics of anther tapetum in Heuchera micrantha // Proc. Indian Acad. Sci. 1979. Vol. 88. P. 309-316. URL: https://doi.org/10.1007/BF03046195 (дата обращения: 19.07.2023).
- Wu Xi.-Q. et al. Distyly and cryptic heteromorphic self-incompatibility in Mussaenda macrophylla (Rubiaceae) // Journal of Systematic sand Evolution. 2015. Vol. 9999. P. 1-13. URL: https://doi.org/10.1111/jse.12142 (дата обращения: 19.07.2023).
- Yeung E.C. et al. Anther, pollen and tapetum development in safflower Carthamus tinctorius L. // Sex Plant Re-prod. 2011. Vol. 24. P. 307-317. DOI: 10.1007/s00497-011-0168-x/ URL: https://pubmed.ncbi.nlm.nih.gov/21573927/ (дата обращения: 19.07.2023).
