Сравнительная оценка размерных характеристик чужеродных видов макрозообентоса Куйбышевского и Саратовского водохранилищ

Автор: Курина Екатерина Михайловна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Общая биология
Статья в выпуске: 2-1 т.20, 2018 года.
Бесплатный доступ
Получены новые данные динамики размерно-возрастной структуры массовых чужеродных видов макрозообентоса - представителей понто-каспийского и понто-азовского комплексов в водохранилищах Средней и Нижней Волги. В результате круглогодичных полевых исследований на глубоководных и прибрежных станциях Куйбышевского и Саратовского водохранилищ установлено увеличение размеров тела особей полихет Hypania invalida (Grube, 1860), двустворчатых моллюсков Dreissena bugensis (Andrussov, 1847), некоторых видов амфипод (Obesogammarus obesus (Sars, 1896), Pontogammarus robustoides (Sars, 1894)) и мизид (Katamysis warpachowskyi Sars, 1893) примерно в 1,2-1,5 раза, в сравнении с таковыми из водоемов-доноров (Каспийское море, Днепро-Бугский лиман, реки Азово-Черноморского бассейна). Наряду с этим отмечено значительное уменьшение размеров тела крупных видов мизид Paramysis ullskyi Czerniavsky, 1882 и Paramysis lacustris (Czerniavsky, 1882), а также корофиид Chelicirophium curvispinum Sars, 1895 и пиявок Archaeobdella esmonti Grimm, 1876. Популяционные различия в размерах особей из водоемов-доноров и водоемов-реципиентов, являются проявлением межпопуляционных адаптивных отношений к факторам среды, из которых температурный фактор для отдельных таксонов, вероятно, является определяющим. Выявлено, что размерная структура двустворчатых моллюсков Dreissena polymorpha (Pallas, 1771) в водохранилищах характеризуется преобладанием мелких особей размером менее 10,0 мм, что свидетельствует о высокой интенсивности их размножения. Установлено также, что после достижения D. polymorpha длины 22,0-25,0 мм (возраст 3-4 года) особи элиминируют, не достигая максимальных размеров моллюсков - 45,0 мм. Получены зависимости массы от длины тела некоторых массовых чужеродных видов донных сообществ: полихет Hypania invalida, амфипод Dikerogammarus haemobaphes (Eichwald, 1841), Pontogammarus robustoides, Stenogammarus dzjubani Mordukhay-Boltovskoy et Ljakhov, 1972, которые описываются уравнениями степенных функций.
Чужеродные виды, макрозообентос, размерные характеристики, куйбышевское водохранилище, саратовское водохранилище
Короткий адрес: https://sciup.org/148205445
IDR: 148205445 | УДК: 574.587
Comparative evaluation of dimensional characteristics of alien species of macrozoobenthos in Kuibyshev and Saratov reservoirs
New data on the dynamics of the size-age structure of massive alien species of macrozoobenthos - representatives of the Ponto-Caspian and Ponto-Azov complexes in the Kuibyshev and Saratov reservoirs of the Middle and Lower Volga are obtained. As a result of year-round research in the deep and shallow stations of the Kuibyshev and Saratov Reservoirs, it was established that the body size of Hypania invalida (Grube, 1860), Dreissena rostriformis bugensis (Andrussov, 1847), some species of amphipods (Obesogammarus obesus (Sars, 1896), Pontogammarus robustoides (Sars, 1894)) and mysids (Katamysis warpachowskyi Sars, 1893) increases in body size of individuals approximately 1.2-1.5 times, in comparison with those of the donor reservoirs (the Caspian Sea, the Dnepro-Bug estuary, rivers of the Azov Sea and the Black Sea basins). In addition, there was a significant reduction in the size of the body of large species of mysids Paramysis ullskyi Czerniavsky, 1882 and Paramysis lacustris (Czerniavsky, 1882), as well as Corophiidae Chelicirophium curvispinum Sars, 1895 and leeches Archaeobdella esmonti Grimm, 1876. Population differences in the size of individuals from donor reservoirs and recipient reservoirs are a manifestation of interpopulation adaptive relationships to environmental factors, of which the temperature factor for individual taxa is probably the determining factor. It was revealed that the dimensional structure of Dreissena polymorpha polymorpha (Pallas, 1771) in the reservoirs is characterized by the predominance of small specimens
Текст научной статьи Сравнительная оценка размерных характеристик чужеродных видов макрозообентоса Куйбышевского и Саратовского водохранилищ
уменьшение течения облегчило продвижение гидробионтов вверх по Волге и привело к увеличению числа чужеродных видов макрозообентоса [3], разнообразие и роль которых в водохранилищах Волжского бассейна продолжает возрастать [4; 5; 6; 7; 8; 9].
Известно, что важной экоморфной характеристикой бентических организмов является их размерно-массовый спектр [10]. Впервые для водохранилищ Средней и Нижней Волги подробно исследован размерный состав чужеродных видов, дана сравнительная оценка максимальных размеров тела чужеродных видов в составе донных сообществ водоемов-доноров и водоемов-реципиентов.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Материалом исследований являются пробы макрозообентоса глубоководных и прибрежных (h<3,0 м) участков Саратовского и Куйбышевского водохранилищ. Отбор проб в Саратовском водохранилище произведен в 2009-2011, 2014, 2016 гг. на 45 станциях (количество проб n=108), в Куйбышевском – в 2009-2011, 2014-2016 гг. на 40 станциях (n=113). Также включены данные ежемесячных круглогодичных сборов макрозообентоса 2009-2011 гг. и ежедекадных сборов 2012 г. на стационарной станции в районе с. Мордово Саратовского водохранилища и исследований на мелководьях левобережной части наиболее эвтрофного Приплотинного плеса Куйбышевского водохранилища, где пробы отбирались ежемесячно с мая по октябрь в 2009-2012 гг.
Количественные пробы отбирали дночерпателем Экмана-Берджи с площадью захвата 250 см2 и 400 см2 по 2 подъема на станции и дночерпателем ДАК-100 (100 см2 x 8). Качественные пробы отбирали гидробиологическим скребком с длиной ножа 20 см и драгой с длиной ножа 40 см (размер ячеи 0,23 мм). Сбор и обработка материала проведена с использованием стандартных гидробиологических методов [11; 12].
За весь период исследований в Куйбышевском и Саратовском водохранилищах было измерено более 33 тыс. особей беспозвоночных. Длина тела полихет и пиявок и высота раковин моллюсков измерялась под бинокуляром с по- мощью окуляр-микрометра с точностью до 0,5 мм и взвешивание произведено с точностью до 0,5 мг. Ракообразные измерялись с точностью до 0,1 мм (за длину тела принято расстояние от ро-струма до основания тельсона).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
В Куйбышевском и Саратовском водохранилищах зарегистрировано 35 чужеродных видов с преобладанием Crustacea: 15 видов Amphipoda, 5 – Mysidacea, 4 – Cumacea, 1 - Isopoda. Mollusca представлены 5 видами, Hirudinea и Polychaeta – по 2 вида и Oligochaeta - 1 вид. Общее число видов-вселенцев составляет примерно 20% видового состава макрозообентоса водохранилищ. Частота встречаемости >50% установлена для двух массовых видов - полихет Hypania invalida (Grube, 1860) и двустворчатых моллюсков Dreissena polymorpha (Pallas, 1771).
Размерные показатели чужеродных видов макрозообентоса в Куйбышевском, Саратовском водохранилищах и водоемах-донорах -Каспийском море; реках Азово-Черноморского бассейна и Днепро-Бугском лимане [13; 14; 15] представлены в табл. 1.
Таблица 1. Минимальная, максимальная и средняя длина тела (L) массовых чужеродных видов макрозообентоса в Куйбышевском (КВ), Саратовском (СВ) водохранилищах и водоемах-донорах (ВД)
|
Вид |
КВ |
СВ |
ВД |
||||
|
L min, MM |
L max, MM |
L cp., MM |
L min, MM |
L max, MM |
L cp., mm |
L max, MM |
|
|
Polychaeta |
|||||||
|
Hypania invalida (Grube, 1860) |
2,0 |
22,0 |
6,8±0,1 |
1,5 |
20,0 |
5,0±0,1 |
14,0 |
|
Hirudinea |
|||||||
|
Archaeobdella esmonti Grimm, 1876 |
2,9 |
27,5 |
6,9±0,3 |
1,7 |
8,0 |
2,1±0,1 |
33,0 |
|
Crustacea |
|||||||
|
Paramysis ullskyi Czerniavsky, 1882 |
3,0 |
21,5 |
“ |
3,0 |
18,5 |
8,5±0,3 |
26,0 |
|
Paramysis lacustris (Czerniavsky, 1882) |
3,0 |
19,0 |
7,9±0,2 |
25,0 |
|||
|
Katamysis warpachowskyi Sars, 1893 |
“ |
“ |
“ |
2,0 |
10,0 |
5,2±0,02 |
8,0 |
|
Pterocuma rostrata Sars, 1894) |
1,8 |
8,0 |
2,0 |
8,0 |
4,2±0,1 |
8,0 |
|
|
Pterocuma sowinskyi (Sars, 1894) |
2,0 |
9,8 |
5,5±0,1 |
2,0 |
8,0 |
4,7±0,2 |
10,5 |
|
Dikerogammarus haemobaphes (Eichwald, 1841) |
1,8 |
17,4 |
6,9±0,3 |
1,5 |
19,2 |
7,2±0,2 |
20,0 |
|
Dikerogammarus caspius (Pallas, 1771) |
“ |
“ |
“ |
2,7 |
18,0 |
7,8±0,2 |
18,0 |
|
Pontogammarus robustoides (Sars, 1894) |
2,2 |
20,2 |
7,8±0,4 |
1,5 |
19,2 |
6,2±0,3 |
18,0 |
|
Pontogammarus maeoticus (Sowinsky, 1894) |
1,8 |
11,4 |
5,0±0,1 |
1,5 |
11,0 |
5,3±0,2 |
12,0 |
|
Obesoogammarus obesus (Sars,1896) |
— |
1,2 |
11,8 |
4,9±0,2 |
8,0 |
||
|
Stenogammarus dzjubani Mordukhay-Boltovskoy et Ljakhov, 1972 |
2,0 |
7,5 |
4,9±0,1 |
1,5 |
8,5 |
3,9±0,1 |
“ |
|
Chaetogammarus warpachowskyi (Sars, 1894) |
1,5 |
7,0 |
3,6±0,1 |
6,5 |
|||
|
Shablogammarus chablensis (Carausu, 1943) |
1,5 |
6,5 |
3,4±0,1 |
3,8 |
|||
|
Chelicorophium curvispinum Sars, 1895 |
3,0 |
6,0 |
“ |
1,4 |
6,0 |
2,9±0,04 |
8,0 |
|
Mollusca |
|||||||
|
Dreissena polymorpha (Pallas, 1771) |
1,2 |
25,0 |
7,8±0,04 |
1,2 |
27,0 |
10,2±0,03 |
45,0 |
|
Dreissena bugensis (Andrussov, 1847) |
1,2 |
33,0 |
10,6±0,02 |
1,1 |
42,0 |
13,7±0,02 |
25,0 |
|
Lithoglyphus naticoides (Pfeiffer, 1828) |
1,5 |
8,8 |
5,4±0,03 |
1,3 |
10,0 |
4,6±0,03 |
10,2 |
|
Theodoxus astrachanicus (Starobogatov in Starobogatov, Filchakov, Antonova et Pirogov, 1994) |
- |
1,2 |
6,5 |
3,7±0,1 |
|||
*- нет данных
Понто-каспийские полихеты Hypania invalida в настоящее время достигли высокой популяционной плотности практически на всех биотопах водохранилищ Волги, предпочитая илистые и илисто-песчаные грунты [5; 16; 17; 18; 19].
Анализ полученных данных свидетельствует об увеличении максимальной длины тела особей полихет Hypania invalida в водохранилищах в 1,5 раза (P=0,008) по сравнению с Каспийским морем. Г.Х. Щербина на основании результатов исследования донных сообществ Горьковского водохранилища сделал вывод об увеличении размера тела полихеты более чем в 2 раза при продвижении на север [6], что подтверждается нашими исследованиями. Среди всех размерных групп в Куйбышевском водохранилище преобладают особи с длиной тела 2,1-7,0 мм, а наименьшее их количество среди наиболее мелких и наиболее крупных экземпляров (рис. 1). Отметим, что средняя длина тела полихеты в целом по водохранилищу оказалась несколько меньшей в сравнении с результатами, полученными для верхних плесов Куйбышевского водохранилища – 8,2±0,3 мм [19]. В Саратовском водохранилище отмечена тенденция уменьшения размеров тела полихет по сравнению с Куйбышевским (табл. 1). В водоеме значительно преобладают мелкие особи размером до 5,0 мм, что говорит о высокой интенсивности размножения полихет (рис. 1).
Уравнение связи между длиной тела полихеты и массой описывается уравнением степенной функции: W (мг) = 0,05 L 2,05 ( R2 = 0,97) (мм).
Хищная каспийская пиявка Archaeobdella esmonti Grimm, 1876 распространяется в волжских водохранилищах вместе с полихетами Hypaniola kowalewskii (Grimm, 1877) и Hypania invalida , которыми питается [20], и имеет высокую частоту встречаемости (до 25%). Размеры пиявки из Куйбышевского и Саратовского водохранилищ не превышали размеров особей из солоноватоводных водоемов (табл. 1), где наибольшая длина тела пиявки составляла 33,0 мм [14]. Наиболее крупные экземпляры отмечены в Рыбинском водохранилище, где фиксированные пиявки имели длину тела до 45,0 мм [21].
В Куйбышевском водохранилище среди всех размерных групп преобладают особи с длиной тела 4,1-8,0 мм (рис. 2). Крупные особи более 10,0 мм встречаются относительно редко в летний период на глубине более 6 м. Размерные показатели A. esmonti в Саратовском водохранилище существенно отличаются от таковых в Куйбышевском: значительно преобладают мелкие пиявки размером 1,5-2,0 мм, крупные особи размером более 10 мм не отмечены (рис. 2). Таким образом, для пиявок Archaeobdella esmonti, как и для массовых полихет Hypania invalida , сохраняется тенденция увеличения размеров тела от южных водохранилищ к северным.
70ЗКЗ'
Куйбышевское вдхр.
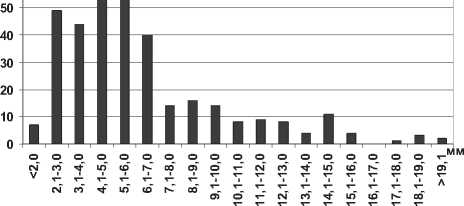
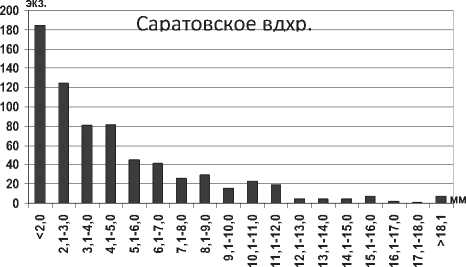
Рис. 1. Размерный состав Hypania invalida в Куйбышевском (n=353) и Саратовском (n=703) водохранилищах в 2009-2012 гг.
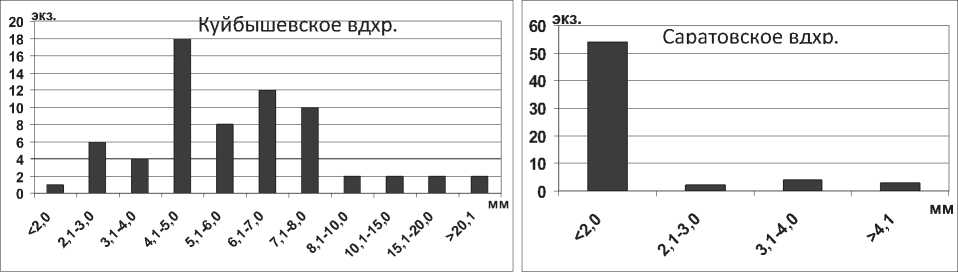
Рис. 2. Размерный состав A. esmonti в Куйбышевском (n=66) и Саратовском (n=63) водохранилищах в 2009-2012 гг.
Обитатель рек и водохранилищ Азово-Черноморского бассейна, солоноватоводная пиявка Caspiobdella fadejewi (Epstein, 1961) проникла в Волгу по Волго-Донскому каналу и в настоящее время встречается практически во всех водохранилищах Волжского каскада [22]. В связи с достаточно высокой чувствительностью пиявки к загрязнению, распределение ее в водохранилищах неравномерно и, по-видимому, имеет разорванный ареал. C. fadejewi в Куйбышевском и Саратовском водохранилищах отмечается единично, минимальная длина тела найденных пиявок - 3,0 мм, максимальная – 9,5 мм. В реках, впадающих в Черное и Азовское моря, длина самых крупных пиявок достигает 13,0 мм [15].
Широко распространенный понто-каспийский бокоплав Dikerogammarus haemobaphes (Eichwald, 1841) в волжских водохранилищах освоил практически все типы биотопов, массово встречается в консорциях моллюсков р Dreissena . Относительно крупные представители отряда Amphipoda, максимальная длина тела которых достигает 20,0 мм (табл. 1). Для популяции D. haemobaphes характерно относительно равномерное распределение особей по длине тела с преобладанием мелких экземпляров от 2,1 до 4,0 мм (рис. 3). Крупные особи бокоплава размером более 14,0 мм малочисленны и отмечены в прибрежной зоне водохранилищ.
Уравнение зависимости массы тела от его длины для D. haemobaphes имеет вид: W (мг)=0,35 L 1,81( R2= 0,95) (мм).
Понто-каспийские амфиподы Dikerogammarus caspius (Pallas, 1771) предпочитают заросшие водной растительностью биотопы и грунты с растительным детритом. В Саратовском водохранилище мелкие особи размером менее 3,0 мм встречаются крайне редко, наибольшее количество экземпляров относится к размерной группе 9,1-10,0 мм (рис. 4), среди которых основную долю составляют половозрелые самки в летний период. В Куйбышевском водохранилище D. caspius встречается единично.
Pontogammarus robustoides (Sars, 1894) – широко распространенный вид в водоемах Европы, размерные характеристики которого изучены достаточно подробно. Так, максимальная длина тела P. robustoides в Каспийском море не превышала 18,0 мм [14], в реке Висла достигала 21,0 мм [23], в водохранилищах Средней и Нижней Волги – 20,2 мм. В вегетационный сезон в водохранилищах преобладают мелкие особи до 5,0 мм, а также отмечено значительное количество экземпляров половозрелых самок размерной группы 8,1-12,0 мм в Куйбышевском водохранилище (рис. 5).
Зависимость массы тела P. robustoides от длины описывается кривой степенной функции: W (мг)=0,14 L 2,23 ( R2 =0,97) (мм).
Распространение Pontogammarus maeticus (Sowinsky, 1894) в Каспийском море и водоемах-реципиентах связано с открытыми песчаными зонами прибрежья с чистыми кварцевыми или ракушечными песками [9; 24]. Максимальная длина тела бокоплава в Каспийском море до-
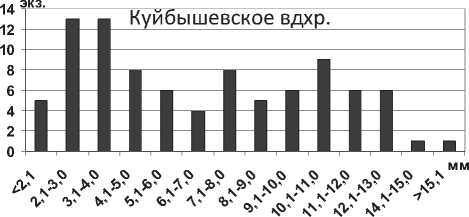
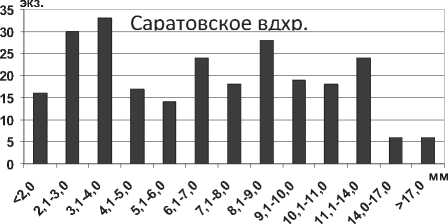
Рис.3 . Размерный состав D. haemobaphes в Куйбышевском (n=86) и Саратовском (n=237) водохранилищах в 2009-2012 гг.
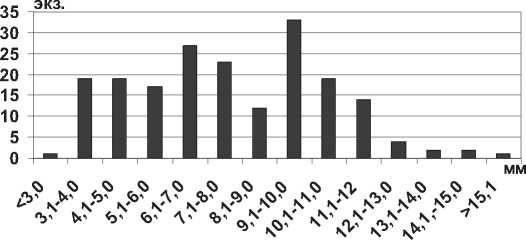
Рис. 4. Размерный состав D. caspius в Саратовском водохранилище в 2009-2012 гг.(n=193)
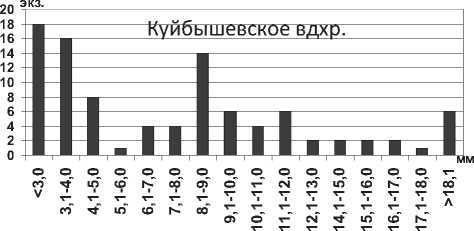
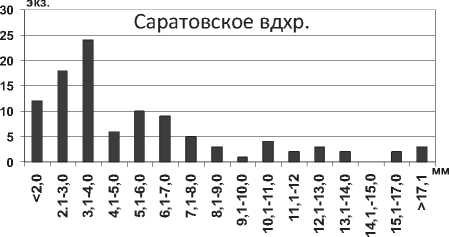
Рис. 5. Размерный состав P. robustoides в Куйбышевском (n=96) и Саратовском (n=104) водохранилищах в 2009-2012 гг.
стигает 12,0 мм, в Куйбышевском и Саратовском водохранилищах – 11,4 мм, однако крупные особи размером более 9,0 мм составляют не более 2% всех измеренных экземпляров. Среди всех размерных групп преобладают мелкие особи с длиной тела 3,1-5,0 мм (рис. 6).
Obesogammarus obesus (G.O. Sars, 1896) – относительно мелкие представители отряда Amphipoda, предпочитающих песчаные и каменистые биотопы, длина тела которых в Каспийском море находится в пределах 5-8 мм [14]. В Куйбышевском водохранилище встречаются крайне редко, длина тела обнаруженных экземпляров (самцов и самок) составляла от 9,0 до 12,0 мм, что значительно превышает размеры данного вида в водоеме-доноре.
В Саратовском водохранилище также отмечена тенденция увеличения максимальных размеров тела O. obesus по сравнению с водоемом-донором (максимальная длина тела бокоплава
- 11,8 мм). Необходимо отметить, однако, что в водохранилище значительно преобладают мелкие особи (более 78% всех измеренных экземпляров относятся к размерным группам менее 6,0 мм) (рис. 7). В летний период отмечено увеличение количества половозрелых самок размерной группы 9,1-10,0 мм.
Псаммофильные амфиподы Stenogammarus dzjubani Mordukhay-Boltovskoy et Ljakhov, 1972. были найдены только в Куйбышевском и Саратовском водохранилищах. В Каспийском море S. dzjubani не отмечается, хотя, по мнению Ф.Д. Мордухай-Болтовского и С.М. Ляхова, сделавших описание данного вида, возможно его нахождение в море, так как вид относится к роду, эндемичному для Понтокаспия [25]. В водохранилищах преобладают мелкие особи с длиной тела до 5,0 мм (рис. 8). Доля половозрелых самцов и самок размерной группы более 6,0 мм не превышает 21%.
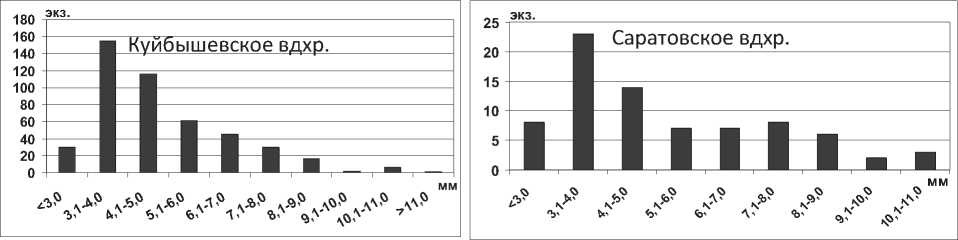
Рис. 6. Размерный состав P. maeoticus в Куйбышевском (n=462) и Саратовском (n=78) водохранилищах в 2009-2012 гг.
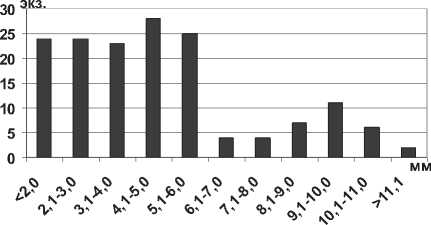
Рис. 7. Размерный состав O. obesus в Саратовском водохранилище в 2009-2012 гг.(n=158)
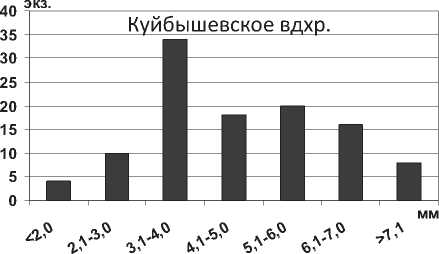
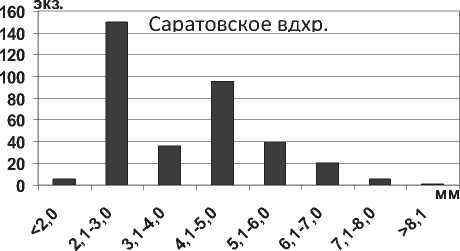
Рис. 8. Размерный состав S. dzjubani в Куйбышевском (n=110) и Саратовском водохранилищах (n=354) в 2009-2012 гг.
Зависимость массы тела от его длины для S. dzjubani описывается уравнением степенной функции: W (мг)=0,40* L 1,67 ( R 2 =0,94) (мм).
Амфиподы понто-каспийского происхождения Chaetogammarus warpachowskyi (G.O.Sars, 1894) широко распространены в Саратовском водохранилище, предпочитают малые глубины и илисто-песчаные грунты. Для бокоплава характерно равномерное распределение особей по длине тела (рис. 9), исключение составляли крупные экземпляры более 5,0 мм, единично отмеченные на свале глубин 4-6 м.
В Куйбышевском и Саратовском водохранилищах отмечен также понто-азовский бокоплав Shablogammarus chablensis (Carausu, 1943) – обитатель эстуариев рек и прибрежных озер северо-западной и западной частей Черного и Каспийского морей. Известно, что длина тела S. chablensis в реках Азово-Черноморского бассейна составляет 3,8 мм у самцов и 3,5 мм у самок [13]. В Куйбышевском и Саратовском водохранилище отмечены более крупные экземпляры этого вида, так, максимальная длина тела бокоплава в водохранилищах - 6,5 мм (табл. 1), а особи размером более 4,0 мм составляют до 45% всех измеренных экземпляров (рис. 10).
Представитель сем. Corophiidae - Chelicorophium curvispinum Sars, 1895 является одним из самых распространенных в последнее десятилетие видов понто-каспийского комплекса, инвазионная активность которого объясняется особенностями биологии и экологических характеристик вида: фильтрационно-седиментационный тип питания с возможностью переключения на потребление водорослевых обрастаний и детрита, высокая плодовитость, поливольтинность, быстрый рост, высокая конкурентоспособность, устойчивость к загрязнению среды, способность обитать в широком диапазоне экологических факторов и др. [26; 27; 28; 29]. В Куйбышевском и Саратовском водохранилищах значительно преобладают мелкие особи размерной группы 2,1-3,0 мм (рис. 11).
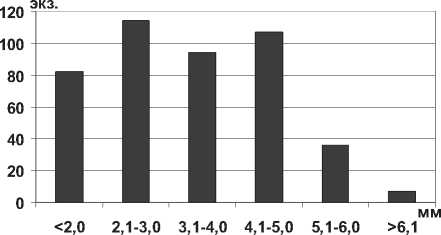
Рис. 9. Размерный состав С. warpachowskyi в Саратовском водохранилище в 2009-2012 гг. (n=440)
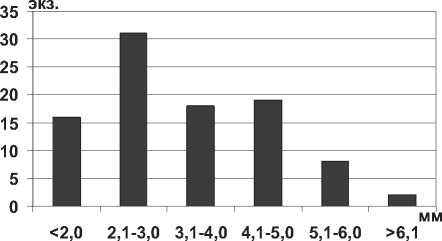
Рис. 10. Размерный состав S. chablensis в Саратовском водохранилище в 2009-2012 гг. (n=94)
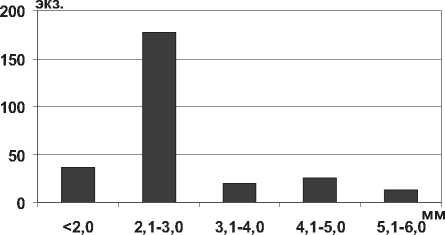
Рис. 11. Размерный состав С. curvispinum в Саратовском водохранилище в 2009-2012 гг. (n=273)
Мизиды Paramysis ullskyi Czerniavsky, 1882 в водохранилищах обитают на глубинах до 5 м, реже до 18 м, часто встречаются на песчаных и ракушечных грунтах, образуют скопления на открытых мелководьях, а также среди зарослей вышей водной растительности. Длина тела наиболее крупных экземпляров мизид P. ullskyi в Каспийском море может достигать 26,0 мм [14]. В водохранилищах Средней и Нижней Волги столь крупные особи не отмечены, максимальная длина тела P. ullskyi в Саратовском водохранилище – 18,5 мм, в Куйбышевском – 21,5 мм (табл. 1). В водоемах доминируют особи размером 5,1-9,0 мм, наименьшее количество особей среди мелких (менее 4,0 мм) и крупных (более 15,0 мм) экземпляров (рис. 12).
Paramysis lacustris (Czerniavsky, 1882) – также относительно крупный представитель отряда Mysidacea, максимальная длина тела которого в Каспийском море составляет 25,0 мм [14]. В Саратовском водохранилище максимальные размеры тела самцов не превышают 19,0 мм, крупные особи (более 15,0 мм) немногочисленны и составляют не более 3% всех измеренных экземпляров. Наибольшее количество особей относятся к размерной группе 5,1-7,0 мм (рис. 13).
Мизиды Katamysis warpachowskyi G.O.Sars, 1893 отмечены на жестких грунтах затопленной поймы Саратовского водохранилища и на пес- чаных мелководьях Куйбышевского водохранилища. Длина тела в Каспийском море не превышает 8,0 мм [14]. Длина тела в разных водоемах составляет 4,5-8,5 мм [30], однако, на пойменных участках Саратовского водохранилища обнаружены более крупные экземпляры самцов и самок с длиной тела до 9,0-10,0 мм. Вместе с тем максимального развития получили особи размерной группы 4,1-6,0 мм (рис. 14).
Кумовые ракообразные Pterocuma sowinskyi (G.O.Sars, 1894) – наиболее массовый вид Северного Каспия, обитает на глубинах 2-25 м и встречается на различных грунтах (илистых, песчанистых, ракушечных). Максимальная длина тела P. sowinskyi в Куйбышевском и Саратовском водохранилищах не достигает максимальных значений длины тела раков в Каспийском море (табл. 1). В Куйбышевском водохранилище преобладают особи размерной группы 5,1-7,0 мм (рис. 15), в целом по водоему отмечено значительное развитие мелких особей. В Саратовском водохранилище наибольшее число экземпляров имели длину тела менее 5,0 мм (67% всех измеренных экземпляров) (рис. 15).
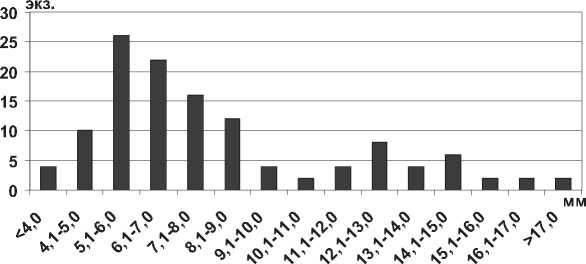
Рис. 12. Размерный состав P. ullskyi в Саратовском водохранилище в 2009-2012 гг. (n=126)
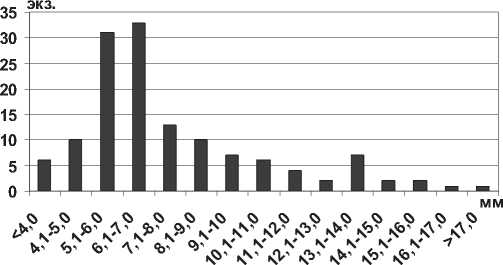
Рис. 13. Размерный состав P. lacustris в Саратовском водохранилище в 2009-2012 гг. (n=135)
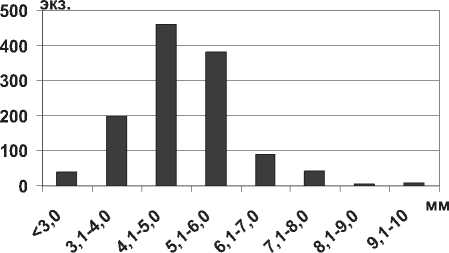
Рис. 14. Размерный состав K. warpachowskyi в Саратовском водохранилище в 2009-2012 гг. (n=1227)
там. В Саратовском водохранилище наблюдалось значительно преобладание мелких особей размером менее 5,0 мм (81% всех измеренных экземпляров) (рис. 16).
Брюхоногий моллюск Lithoglyphus naticoides (С. Pfeiffer, 1828) – представитель понто-азовской фауны, массово расселился в прибрежной зоне волжских водохранилищ. Распределение моллюсков по размерному составу в Куйбышевском водохранилище отличается большей неравномерностью, чем в Саратовском (рис. 17). В Куйбышевском водохранилище значительно преобладали моллюски размерной группы 5,1-7,0
мм, наименьшее количество экземпляров среди наиболее мелких и наиболее крупных особей. В
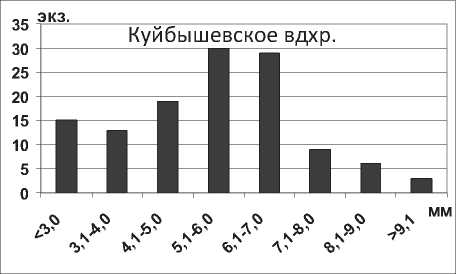
Рис. 15. Размерный состав P. sowinskyi в Куйбышевском (n=124) и Саратовском (n=66) водохранилищах в 2009-2012 гг.
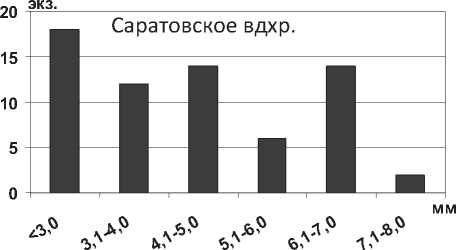
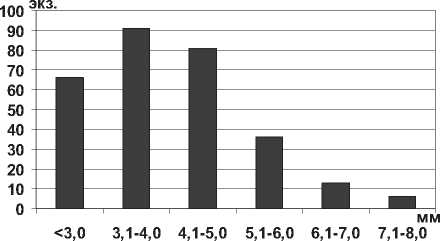
Рис. 16. Размерный состав P. rostrata в Саратовском водохранилище в 2009-2012 гг. (n=293)
целом, полученные нами данные по размерным характеристикам L. naticoides в нижних плесах Куйбышевского водохранилища согласуются с данными, полученными для верхних плесов водохранилища [32]. Авторы отмечают, что максимальные размерные показатели характерны для глубоководных районов водохранилища (глубина 5-15 м). Однако, в наших исследованиях достоверной зависимости между размерными показателями L. naticoides и глубиной не выявлено. В Саратовском водохранилище относительно равномерно представлены все размерные группы от 2,1 до 8,0 мм (рис. 17). Крупные моллюски более 8,1 мм встречаются редко и составляют менее 3% всех измеренных экземпляров.
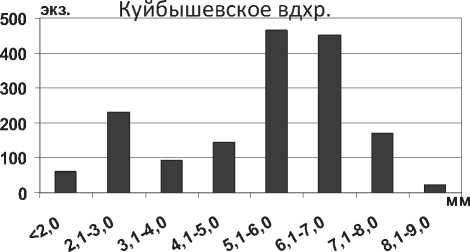
Рис. 17. Высота раковины L. naticoides в Куйбышевском водохранилище (n=1637) и Саратовском водохранилищах (n=2429) в 2009-2012 гг.
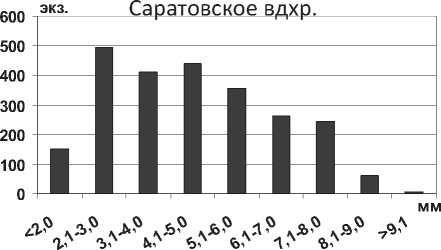
Начиная с 50-х годов прошлого века, в литературе встречаются упоминания о вселении Theodoxus pallasi Lindholm, 1924 из дельты р. Волга в водохранилища Нижней Волги [33; 34]. Отмеченный вид, по-видимому, следует относить к Theodoxus astrachanicus (Starobogatov in Starobogatov, Filchakov, Antonova et Pirogov, 1994), обитающему в дельте Волги и выделенному в отдельный вид, в отличие от T. pallasi, именно в качестве пресноводной формы [35]. T. astrachanicus в Саратовском водохранилище имеет высоту раковины от 1,2 мм до 6,5 мм, ширину – от 1,5 до 10,2 мм. Высота раковины близкого по происхождению вида Theodoxus pallasi в Каспийском море может достигать 5,5 мм, ширина раковины - до 7,5 мм. В Саратовском водохранилище преобладают особи размерной группы 4,1-5,0 мм (рис. 18), крупные особи размером более 6 мм немногочисленны, отмечены в прибрежной зоне водохранилища на каменистом грунте.
Представители рода Dreissena – Dreissena polymorpha (Pallas, 1771) и D. bugensis (Andrusov, 1897) - единственная группа пресноводных моллюсков, способных прикрепляться к твердому субстрату (камням, древесине, раковинам других моллюсков) и образовывать массовые скопления в виде друз, как это делают многие морские виды. Дрейссены часто образуют кон-
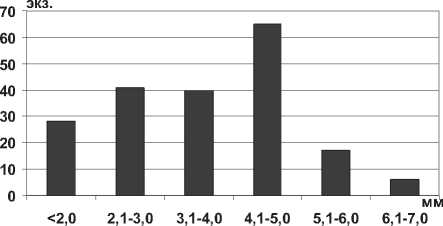
Рис. 18. Высота раковины T. astrachanicus в Саратовском водохранилище в 2009-2012 гг. (n=197)
сорции с определенными видами, между элементами сообществ возникают топические и трофические взаимодействия [36].
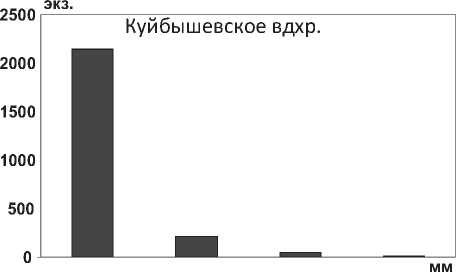
Размерная структура моллюсков D. polymorpha в водохранилищах характеризуется преобладанием особей размером <10,0 мм (рис. 19), что свидетельствует о высокой интенсивности их размножения. Установлено, что после достижения моллюсками D. p. polymorpha длины 22,0-25,0 мм (возраст 3-4 года) особи элиминируют, не достигая максимальных размеров моллюсков – 45,0 мм (табл. 1). Аналогичные данные были получены для D. p. polymorpha из Рыбинского водохранилища [37] .
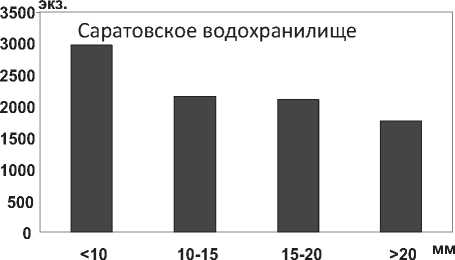
В отличие от D. p. polymorpha , максимальная длина тела моллюска D. bugensis в Куйбышевском водохранилище достигает 33,0 мм, в мелководной зоне Саратовского водохранилища – 42,0 мм, в 1,6 раза (P=0,004) превышая максимальную длину особей из Днепро-Бугского лимана. Характерно, что размерная структура популяции D. bugensis отличается преобладанием мелких особей в глубоководном Куйбышевском водохранилище и их относительно равномерным распределением на пойме Саратовского водохранилища (рис. 20). Констатируется быстрый рост моллюсков всех размерных групп на пойменных биотопах в условиях более высоких температур [38; 39], по сравнению с их развитием в глубоководных участках.
Также в водохранилищах зарегистрированы следующие редкие и единичные виды: амфиподы Dikerogammarus villosus (длина тела – 4,0-15,0 мм), Stenogammarus compressus (G.O.Sars, 1894) (2,0-7,0 мм), S. similis (G.O.Sars, 1894) (2,5-3,0 мм), Chaetogammarus ischnus (Stebbing, 1898) (6,2-7,0 мм), Chelicophium sowinskyi (2,0-6,0 мм), кумовые ракообразные Pseudocuma cercaroides G.O.Sars, 1894 (2,0-4,5 мм), Caspiocuma campylaspoides (G.O.Sars, 1897) (2,0-4,5 мм), мизиды Limnomysis benedeni (3,0-10,5 мм), двустворчатые моллюски Adacna colorata (Eichwald, 1829) (2,0-9,0 мм), полихеты Manayunkia caspica (Annenkova, 1929) (2,0-4,0 мм).
<10 10-15 15-20 >20
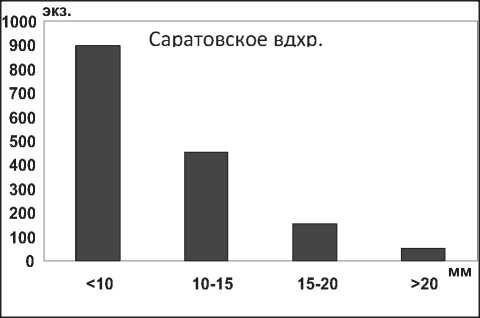
Рис. 19. Размерный состав D. p. polymorpha в Куйбышевском (n=2410) и Саратовском (n=1556) водохранилищах в 2009-2012 гг.
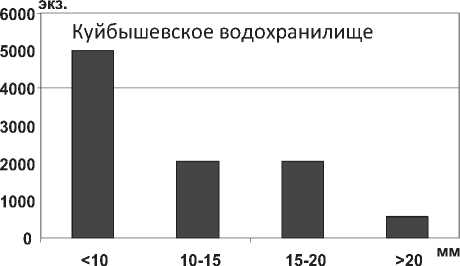
Рис. 20. Размерный состав D. r. bugensis в Куйбышевском (n=9656) и Саратовском (n=9004) водохранилищах в 2009-2012 гг.
ВЫВОДЫ
Таким образом, в водохранилищах Средней и Нижней Волги отмечена тенденция увеличения максимальных размеров тела некоторых чужеродных видов по сравнению с естественным ареалом. Так максимальная длина тела полихет Hypania invalida оказалась более чем в 1,5 раза больше, чем в Каспийском море. Максимальная длина тела половозрелых особей амфипод Obesogammarus obesus в Саратовском водохранилище составляет 11,8 мм, в Каспийском море длина тела этих рачков существенно меньше и находится в пределах 5-8 мм [14]. Также в Саратовском водохранилище наблюдалось увеличение максимальных размеров тела гамма-рид Pontogammarus robustoides и мизид Katamysis warpachowsky .
Наряду с этим отмечено значительное уменьшение размеров тела крупных видов ми-зид Paramysis ullskyi и P. lacustris . Максимальная длина тела особей обоих полов этих видов в Куйбышевском и Саратовском водохранилище составила 21,5 мм, тогда как в Каспийском море размеры мизид достигают 25-26 мм. Амфиподы Chelicorophium curvispinum, пиявки Archaeobdella esmonti и двустворчатый моллюск Dreissena polymorpha в водохранилищах также не достигают максимальных размеров, отмеченных для особей из Каспийского моря. Известно, что различия в размерах – одно из распространенных проявлений межпопуляционных различий в водоеме-доноре и водоеме-реципиенте [40]. Их адаптивное значение связано, прежде всего, с изменением условий поддержания энергетического баланса [41]. Эти различия формируются под влиянием среды, что отчетливо проявляется в популяциях чужеродных видов, обитающих в различных условиях Куйбышевского и Саратовского водохранилищ.
Список литературы Сравнительная оценка размерных характеристик чужеродных видов макрозообентоса Куйбышевского и Саратовского водохранилищ
- Бенинг А.Л. К изучению придонной жизни реки Волги//Саратов: Труды Волжской биол. станции. 1924. 398 с.
- Жадин В.И. Фауна рек и водохранилищ//Тр. ЗИН АН СССР. 1940. Т.5, вып. 3-4. С. 519-991.
- Волга и ее жизнь. Л.: Наука, 1978. 348 с.
- Зинченко Т.Д., Головатюк Л.В., Загорская Е.П., Антонов П.И. Распределение инвазионных видов в составе донных сообществ Куйбышевского водохранилища: анализ многолетних исследований//Известия Самарского научного центра РАН. 2007. Т. 10, № 2. С. 547-558.
- Филинова Е.И., Малинина Ю.А., Шляхтин Г.В. Биоинвазии в макрозообентосе Волгоградского водохранилища//Экология. 2008. №3. С. 206-210.
- Щербина Г.Х. Изменение видового состава и структурно-функциональных характеристик макрозообентоса водных экосистем Северо-запада России под влиянием природных и антропогенных факторов: Автореф. дис. … доктора биол. наук. Спб., 2009. 49 с.
- Яковлева А.В. Фауна и экология бентосных вселенцев верхней части Куйбышевского водохранилища: Автореф. дис.. канд. биол. наук. Казань, 2010. 27 с.
- Курина Е.М. Чужеродные виды донных сообществ Куйбышевского водохранилища и его притоков: структурные показатели и особенности распространения//Известия Самарского научного центра РАН. 2015. Т. 17. № 4-5. С. 925-933.
- Курина Е.М. Разнообразие, динамика распространения и структурная организация чужеродных видов бентоса Саратовского водохранилища//Росс. журнал биол. инвазий. 2016. Т. 9. № 4. С. 69-84.
- Курашов Е.А. Мейобентос как компонент озерной экосистемы. СПб: Алга-Фонд, 1994. 224 с.
- Руководство по гидробиологическому мониторингу пресноводных экосистем. СПб., Гидрометеоиздат, 1992. 318 с.
- Жадин В.И. Методы гидробиологического исследования. М.: Высшая школа, 1960. 190 с.
- Carausu S., Dobreanu E., Manolache C. Fauna Republicii Populare Romini Vol. 4: Crustacea fasc. 4: Amphipoda forme salmastre si de apa dulce. -Academia Republicii Populare Romine, 1955. 407 pp.
- Атлас беспозвоночных . М.: Пищ. пром-сть, 1968. 416 с.
- Лукин Е.И. Фауна СССР. Пиявки. Л.: Наука, 1976. 484 с.
- Калайда М.Л. Современная роль видов-вселенцев Понто-Каспийского комплекса в экосистеме Куйбышевского водохранилища//Материалы российско-американского симп. по инвазийным видам. Борок: ИБВВ РАН, 2003. С. 165-173.
- Степанова Н.Ю., Латыпова В.З., Яковлев В.А. Экология Куйбышевского водохранилища: донные отложения, бентос, бентосоядные рыбы. Казань: Изд-во АН РТ, 2004. 228 с.
- Курина Е.М. Чужеродные виды полихет в Куйбышевском и Саратовском водохранилищах: распространение, особенности натурализации и размерно-массовые характеристики//Вестник АГТУ. Серия: Рыбное хозяйство. 2016. № 2. С. 23-33.
- Яковлев В.А., Яковлева А.В. Полихета Hypania invalida (Polychaeta: Ampharetidae) в Куйбышевском водохранилище: распределение, размерно-весовые характеристики//Росс. журнал биол. инвазий. 2010. №1. С. 44-55.
- Баканов А.И. О появлении пиявки Archaeobdella esmonti (Arhynchobdella, Herpobdellidae) в волжских водохранилищах//Зоол. журн. 1993. Т. 72., вып. 6. С. 135-137.
- Перова С.Н. Структурные характеристики каспийского вселенца -пиявки Archaeobdella esmonti Grimm в Рыбинском водохранилище//Росс. журн. биол. инвазий (ISSN 1996-1499. www. sevin.ru). 2011. № 2. С. 135-141.
- Лапкина Л.Н., Свирский А.М. Пиявки Caspiobdella fadejewi (Epstein, 1961) и Acipenserobdella volgensis (Zykoff, 1903) -вселенцы в водохранилищах Верхней и Средней Волги//Американо-Российск. симп. по инвазионным видам: Тезисы докладов. Ярославль, 2003. С. 109-111.
- Bącela K., Konopacka A. The life history of Pontogammarus robustoides (G.O. Sars, 1894) -an alien amphipod species in Polish waters//Journal of Crustacean Biology. 2005. №25. P. 190-195.
- Гусейнов K.M. Ценоз и экология Pontogommarus maeoticus в Каспийском море // Актуальные вопросы экологии и охраны природы экосистем южных регионов и сопредельных территорий // Матер. XVI межресп. конф. Краснодар, 2003. С. 145 - 146.
- Мордухай-Болтовской Ф.Д., Ляхов С.М. Новый вид амфипод рода Stenogammarus (Gammaridae) в бассейне Волги//Зоол. журн. 1972. Т. 51, вып. 1. С. 21-27.
- Rajagopal S., Van der Velde G., Pfaffen B.G.P., Bij de Vaate A. Growth and production of Corophium curvispinum G.O.Sars, 1895 (Amphipoda), an invader in the Lower Rhine//The biodiversity crisis and Crustacea. Eds. J. C. von Vaupel Klein and F. R. Schram. Rotterdam: the Netherlands, 1998. P.3-33.
- Lee C.E., Bell M.A. Causes and consequences of recent freshwater invasions by saltwater animals//Trends in Ecology & Evolution. 1999. V.14, №7. P. 284-288.
- Arndt E., Fiedler S., Böhme D. Effects of invasive benthic macroinvertebrates on assessment methods of the EU Water Frame Work Directive//Hydrobiologia. 2009. Vol. 635. P. 309-320.
- Литоральная зона Ладожского озера . Спб.: Нестор-История, 2011. 416 с.
- Daneliya M.E. On the distribution of Katamysis warpachowskyi Sars 1877 (Crustacea, Mysida) in the Don river Basin//Известия ВУЗов. Северо-Кавказский регион. Естественные науки. 2001. Вып.2. С. 49-50.
- Jaume D., Boxshall G.A. Global diversity of cumaceans & tanaidaceans (Crustacea: Cumacea & Tanaidacea) in freshwater//Hydrobiologia. 2008. Vol. 595. P. 225-230.
- Яковлев В.А., Ахметзянова Е.Ш., Яковлева А.В. Встречаемость, распределение и размерно-весовые характеристики Lithoglyphus naticoides (Gastropoda, Hydrobiidae) в верхней части Куйбышевского водохранилища//Росс. журнал биол. инвазий. 2009. №1. С. 39-52.
- Кирпиченко М.Я., Ляхов С.М. О проникновении Theodoxus pallasi Lindh. (Mollusca, Gastropoda) в Нижнюю Волгу//Материалы по биологии и гидрологии Волжских водохранилищ. М.; Л.: Наука, 1963. С. 17-18.
- Пирогов В.В., Фильчаков В.А., Зинченко Т.Д., Карпюк М.И., Едский Л.Б. Новые элементы в составе бентофауны Волго-Камского каскада водохранилищ//Зоол. журн. 1990. Т. 69, № 9. С. 138-142.
- Старобогатов Я.И., Фильчаков В.А., Антонова Л.А., Пирогов В.В. Новые данные о моллюсках и высших ракообразных дельты Волги//Вестн. зоол. 1994. Т. 28. № 4-5. С. 8-12.
- Беклемишев В.Н. О классификации биоценологических (симфизиологических) связей//Бюл. МОИП. Отд. биол. 1951. Т.56, вып. 5. С. 3-30.
- Пряничникова Е.Г. Структурно-функциональные характеристики дрейссенид Рыбинского водохранилища: Автореф. дисс. … канд. биол. наук. Борок. 2012. 21 с.
- Stanczykowska A. 1976. Biomass and production of Dreissena polymorpha (Pallas) in some Masurian Lakes. Ekol. Pol.24:103-112.
- Место вида в биоценозах/А.Ю. Каратаев, В.П. Ляхнович, С.А. Афанасьев, Л.Е. Бурлакова, В.П. Закутский, С.М. Ляхов, М.П. Мирошниченко, Т.Г. Мороз, М.Я. Некрасова, С.П. Нечваленко, И.А. Скальская, Т.Г. Харченко, А.А. Протасов//Дрейссена, Dreissena polymorpha (Pall.) (Bivalvia, Dreissenidae). Систематика, экология и практическое значение . 1984. М.: Наука. С. 180-205.
- Шиганова Т.А. Чужеродные виды в экосистемах южных внутренних морей Евразии: Автореф. дис. … доктора биол. наук. Москва, 2009. 57 с.
- Винберг Г.Г. Интенсивность обмена и размеры ракообразных//Журн. общей биологии. 1950. Т. 12, №5. С. 367-380.
