Сравнительные характеристики корневых систем и корневой экссудации у синтетического, примитивного и современного сортов пшеницы

Автор: Шапошников А.И., Моргунов А.И., Акин Б., Макарова Н.М., Белимов А.А., Тихонович И.А.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Местные и селекционные сорта - факторы адаптации
Статья в выпуске: 1 т.51, 2016 года.
Бесплатный доступ
Поиск способов повышения адаптации пшеницы к засухе в настоящее время считается важнейшей задачей в селекции новых сортов этой культуры. В представляемой работе проведена сравнительная оценка генотипических различий для основных характеристик корневой системы пшеницы, способных влиять на адаптацию растений к неблагоприятным условиям окружающей среды, в том числе к засухе. Использовали три генотипа пшеницы, воспроизводящие основные этапы эволюции гексаплоидной мягкой пшеницы ( Triticum aestivum L.): синтетическую пшеницу, полученную скрещиванием сорта LEUCURUM 84693 твердой пшеницы ( Triticum durum Desf., геном АВ, Украина) и дикого злака эгилопса Тауша ( Aegilops tauschii Coss., геном D, Турция); традиционный местный турецкий сорт мягкой пшеницы Albostan (геном ABD, Турция, провинция Невшехир); сорт современной селекции Karahan (геном ABD, Турция). Между сортами выявлены различия по диаметру корней растения, биомассе стеблей и соотношению биомассы побега и корня. Удаление побега вызывало увеличение длины корней, объема и ветвления корневой системы и соотношения биомассы побег:корень у синтетической пшеницы и в меньшей степени у сорта Albostan, но снижало величину этих показателей у сорта Karahan. Средние значения десяти измеренных ростовых параметров у растений пшеницы с удаленным побегом относительно контрольных составили +28 % для синтетической пшеницы, 0 % для сорта Albostan и -37 % для сорта Karahan. Эти результаты свидетельствуют о более высокой способности к ревегетации у синтетической пшеницы по сравнению с сортом Karahan. Впервые проведен сравнительный анализ корневой экссудации аминокислот, сахаров и органических кислот у генотипов пшеницы различных уровней эволюции. Показано, что сорт Karahan наиболее активно выделяет в ризосферу триптофан (0,05 мг/г сухих корней в сутки), гистидин (0,12 мг/г сухих корней в сутки), фенилаланин (0,45 мг/г сухих корней в сутки). Общее количество выделяемых корнями сахаров (в основном фруктозы, глюкозы и мальтозы) у сорта Karahan составило 55 мг/г сухих корней в сутки и было соответственно в 5 и 3 раза выше, чем у синтетического генотипа и сорта Albastan. Количество органических кислот в экссудатах всех изучаемых генотипов оказалось примерно одинаковым и составляло около 1,8 мг/г сухих корней в сутки. Эти данные отражают низкую способность сорта Karahan контролировать выделение фотосинтатов в окружающую среду через транспортные системы корней. Возможно, что высокая активность корневой экссудации аминокислот, особенно триптофана (предшественника в биосинтезе ауксинов), и сахаров может приводить к дефициту указанных веществ и замедлять регенерацию побегов у сорта Karahan. Полученные результаты указывают на различия в функционировании корневой системы у изученных генотипов пшеницы, связанные с ревегетацией побега и корневой экссудацией. Выяснение природы обнаруженных различий требует дальнейших исследований.
Пшеница, корневая экссудация, ревегетация, устойчивость к засухе
Короткий адрес: https://sciup.org/142133660
IDR: 142133660 | УДК: 633.11:58.056:[581.1+575.1/.2 | DOI: 10.15389/agrobiology.2016.1.68rus
Comparative characteristics of root systems and root exudation of synthetic, landrace and modern wheat varieties
Finding ways to increase the wheat adaptation to drought is now considered as a major problem in breeding new varieties of this crop. This study was conducted to evaluate genotypic differences in fundamental root traits which may have effect on wheat adaptation to unfavorable environments, including drought. Three wheat genotypes representing various evolution levels of hexaploid bread wheat ( Triticum aestivum L.) were used: synthetic wheat developed by crossing variety LEUCURUM 84693 of durum wheat ( Triticum durum Desf., genome АВ, Ukraine) with wild Tausch's goat grass ( Aegilops tauschii Coss., genome D, Turkey), landrace of bread wheat Albostan (genome ABD, Turkey, province Nevşehir), and a modern cultivar Karahan (genome ABD, Turkey). The varieties differed in root diameter, shoot biomass and shoot/root ratio. The removal of above ground biomass caused increase in the root length, number of tips, number of forks, number of crosses and shoot/root biomass ratio of synthetic wheat and Albostan, but decreased these parameters in Karahan. Averaged values of ten measured growth parameters of the plants with removed above ground biomass relatively to the control plants were +28 % for synthetic wheat, 0 % for landrace Albostan and -37 % for cultivar Karahan. These results showed a higher ability of synthetic wheat but lower ability of Karahan to recover from cutting stress and to revegetate. For the first time a comparative analysis of root exudation (amino acids, sugars and organic acids) by wheat genotypes having different levels of evolution was performed. It was shown that Karahan was characterized by high exudation of tryptophan (0.05 mg/g dry roots per day), histidine (0.12 mg/g dry roots per day) and phenylalanine (0.45 mg/g dry roots per day). Total amount of sugars (mostly fructose, glucose and maltose) exuded by Karahan was 55 mg/g dry roots per day, that was 5 and 3 times higher as compared to synthetic genotype and Albastan. The quantum of organic acids in exudates of all studied genotypes was approximately similar and amounted to about 1.8 mg/g of dry roots per day. The results suggested low ability of Karahan to control the flow rate of carbohydrates from roots to the environment. We propose that high root exudation of tryptophane (precursor in biosynthesis of auxins) and sugars may result in deficit of these compounds and involved in impaired shoot regeneration of Karahan. The results demonstrated differences in the functioning of the root system of primitive hexaploid wheat (synthetic) compared to landrace and modern cultivated variety. The nature of these differences requires more investigation.
Текст научной статьи Сравнительные характеристики корневых систем и корневой экссудации у синтетического, примитивного и современного сортов пшеницы
Пшеница (Triticum spp.) — одна из основных сельскохозяйственных культур, обеспечивающих миллиарды людей продуктами питания. По имеющимся оценкам , к 2050 году производство зерна пшеницы необходимо увеличить с современных 650-700 млн т до 1 млрд т. Однако возможности мобилизации генетических ресурсов этой культуры заметно исчерпались со времен «зеленой революции». Этим (в сочетании с неблагоприятными изменениями климата) частично объясняется тот факт, что у используемых сортов практически достигнут биологический предел урожайности. Недавние исследования на озимой пшени-
∗ Работа по получению корневых экссудатов поддержана грантом Российского фонда фундаментальных исследований (15-04-09023-а). Хроматографический анализ корневых экссудатов выполнен при поддержке гранта Российского научного фонда (14-26-00094). Эксперимент в CIMMYT поддержан CRP WHEAT и Министерством продовольствия, сельского хозяйства и животноводства Турции.
це в Европе, Центральной и Восточной Азии и в США выявили повышение средней температуры воздуха в критические для развития растений периоды (1). Хотя более теплая зима, как правило, приводила к росту урожайности этой культуры, повышенный температурный фон поздней весной или летом снижал ее из-за засухи. Адаптация к засухе в настоящее время считается важнейшей задачей в селекции пшеницы, вследствие чего получила развитие всемирная программа поиска способов повышения устойчивости сортов к засухе и другим абиотическим стрессам (International Winter Wheat Improvement Program, IWWIP, .
Существуют различные методики оценки засухоустойчивости пшеницы: по урожаю и биохимическому составу зерна, а также по признакам, способствующим высокой урожайности в условиях недостатка влаги, например по строению корневой системы как важнейшего органа, ответственного за поглощение и транспорт воды в побеги (2). Показано, что эффективность поглощения влаги из засушливых почв в основном определяется величиной контактной поверхности корневой системы, которая, в свою очередь, обусловлена степенью ветвления корней и длиной корневых волосков (3).
Большинство сосудистых растений способны улучшать минеральное и водное питание, формируя на корнях и в прикорневой области (ризосфере) ассоциации и симбиозы с почвенными микроорганизмами (4-7). Основным источником углерода и энергии для таких микроорганизмов служат органические вещества (главным образом сахара, органические кислоты и аминокислоты), выделяемые корнями в ризосферу (8). Корневые экссудаты также играют важную роль в снабжении растений питательными элементами, недостаток которых может проявляться в условиях засухи (9, 10).
Величина соотношения корень:побег (по биомассе) при засухе возрастает, но общая сухая биомасса корней в сравнении с таковой в нормальных условиях увеличивается очень редко. Тем не менее, плотность корневой системы в расчете на единицу площади листьев при дефиците влаги обычно растет (11). Фотосинтаты, которые могли быть использованы для развития новых корней, расходуются на рост уже существующих, и те в результате прорастают в более глубокие слои почвы. У пшеницы осушение почвы приводило к ограничению роста корней в верхнем слое почвы (30 см) и вызывало их распространение в глубину (12). После восстановления нормального режима почвенного увлажнения у растений возобновлялся быстрый рост корней в верхнем слое почвы и прекращалось развитие корневой системы в более глубоких слоях.
Непосредственная оценка параметров корневой системы при проведении исследований и селекционных работ в полевых условиях трудоемка и требует специального оборудования. В настоящее время для определения способности корней снабжать растения питательными веществами и влагой часто используется косвенный показатель — температура растительного покрова (13). Генотипы пшеницы с хорошим водным балансом и засухоустойчивостью, как правило, имеют более низкую температуру, чем растения, находящиеся в условиях засухи. Однако измерение температуры растительного покрова, будучи простым и быстрым методом, все же требует особых условий (солнечный день без ветра), которые складываются не всегда. Еще один косвенный метод — измерение способности растений к регенерации после срезания надземной биомассы. Удаление надземной биомассы у пшеницы и других злаковых культур широко практикуется на посевах двойного назначения на южных равнинах США, в Южной Америке и Австралии, где в зимний период поля используются как пастбища, после чего посевы выращивают до созревания урожая зерна (14).
Тем не менее, с помощью этой техники до сих пор не было проведено исследований для оценки адаптации генотипов пшеницы к засухе и взаимосвязи между процессами регенерации побегов и корневой экссудацией.
Разнообразие генетических ресурсов пшеницы составляет основу для повышения устойчивости растений к абиотическим стрессам, включая засуху. Все больше исследователей и селекционеров ищут новые гены засухоустойчивости вне современных и детально описанных генотипов. Потенциальным источником таких генов служат традиционные местные сорта, столетиями культивировавшиеся в обособленных и засушливых регионах. В Турции местные примитивные сорта пшеницы до сих пор выращиваются по всей стране, особенно в отдаленных горных районах, оставаясь источником генетического разнообразия и ценных признаков. Синтетическая пшеница, полученная скрещиванием твердой пшеницы ( Triticum durum Desf.) и дикого злака эгилопс Тауша ( Aegilops taushii Coss.) также обладает рядом полезных качеств, включая засухоустойчивость (15).
Нашей целью была оценка развития корневой и стеблевой системы у трех генотипов пшеницы (примитивный, современный и синтетический сорта), представляющих разные этапы эволюции гексаплоидной пшеницы, в нормальных условиях и под воздействием стресса, вызванного удалением надземной биомассы. При этом впервые проведено сравнение корневой экссудации фотосинтатов (органических кислот, сахаров и аминокислот) такими генотипами растений с выявленными генотипическими различиями в процессах функционирования корневой системы и регенерации побегов.
Методика . Объектами исследований были генотипы пшеницы, полученные по программе IWWIP из Международного центра улучшения кукурузы и пшеницы (International Maize and Wheat Improvement Center — CIMMYT, Турция), которые соответствуют трем уровням эволюции культуры: синтетическая форма, созданная скрещиванием твердой пшеницы ( T. durum Desf.) сорта украинского происхождения LEUCURUM 84693 (геном AB) и эгилопса Тауша ( A. taushii Coss.) с геномом D; традиционный сорт гексаплоидной пшеницы ( T. aestivum L.) Albostan (геном ABD) местной селекции в турецкой провинции Невшехир (Nev s ehir ) ; засухоустойчивый современный сорт пшеницы ( T. aestivum L.) Karahan (геном ABD) турецкой селекции, выведенный в 1990-х годах стандартными методами.
Вегетационный эксперимент по изучению роста побегов и корней и реакции растений на удаление побегов выполнялся на базе турецкого отделения CIMMYT (Izmir, Турция). Растения высаживали в сосуды с 1 кг песка (2 растения в сосуде, 4 сосуда на вариант) и выращивали в теплице с естественным температурным и световым режимом (температура воздуха в колебалась от 8-12 °С ночью до 20-24 °С днем) в феврале-марте 2014 года до стадии 4-5-го листа (перед выходом в трубку). Полив осуществляли дважды в неделю (по 200 мл воды на сосуд). Питательный раствор (N20P20K20, 250 г/л, «Harmony Imports», США) вносили 3 раза: на стадии 2-3-го листа, перед кущением и через 1 нед после начала кущения. Через 5-6 нед проводили первичную оценку изучаемых параметров. Растения из двух сосудов выкапывали, отмывали корневую систему и сканировали ее с помощью Epson Perfection V700 («Epson America, Inc.», США), используя для обработки данных программу WinRHIZO («Regent Instruments, Inc.», Канада). Затем корни и побеги высушивали при комнатной температуре до постоянной массы. В двух оставшихся сосудах побеги срезали и оставляли растения для ревегетации на 3-4 нед, после чего оценивали параметры побегов и корневой системы, как описано выше.
Эксперименты с корневыми экссудатами проводили во Всероссий- ском НИИ сельскохозяйственной микробиологии с использованием гидропонной культуры. Для получения корневых экссудатов семена поверхностно стерилизовали 6 мин 0,1 % HgCl2, промывали стерильной водой и проращивали 2 сут в чашках Петри в темноте при 27 °С. Проросшие семена высаживали в стеклянные стерильные сосуды с 100 мл деионизированной воды и сетками из нержавеющей стали (10 семян на сосуд, 3 сосуда на один генотип). Проростки культивировали в климатической камере ADAP-TIS-A1000 («Conviron», Великобритания) 5 сут (освещение 200 мклк^м-2^с-1, 16-часовой фотопериод с минимальной и максимальной температурами соответственно 18 и 22 °С). Через 3 сут из каждого сосуда отбирали по 0,1 мл раствора и высевали на чашки Петри с агаризованной средой (Bacto Pseudomonas agar, «Difco International BV», Нидерланды) для контроля стерильности. Через 5 сут растения вынимали из сосудов, высушивали и определяли сухую биомассу побегов и корней. Растворы из трех сосудов объединяли, фильтровали под вакуумом через нейлоновые фильтры (0,45 мкм; «Corning, Inc.», США) и упаривали под вакуумом до объема 5 мл на ротационном испарителе BUCHI R-200 («BUCHI Labortechnik AG», Швейцария). Из полученных концентратов корневых экссудатов отбирали по 100 мкл для анализа аминокислот, а остальной раствор пропускали через колонку с ионообменной смолой DOWEX 50Wx8 («Sigma-Aldrich Co.», США) для получения очищенной фракции органических кислот и сахаров, выпаривали досуха под вакуумом и растворяли в 0,5 мл деионизированной воды. Хроматографический анализ проводили с использованием системы Waters ACQUITY UPLC H-Class («Waters», США). Сахара разделяли на колонке SUPELCOSIL LC-NH2 («Supelco Gland», Швейцария). Для обнаружения сахаров применяли рефрактометрический детектор Waters 2414 («Waters», США). Аминокислоты (кроме L-триптофана) анализировали методом Waters AccQ-Tag («Waters», США) по стандартной методике производителя с помощью флуоресцентного детектора. Количество L-триптофана в корневых экссудатах определяли, используя разделение на колонке Waters UPLC RP-18 Shield («Waters», США) с обнаружением на флуоресцентном детекторе ACQUITY UPLC («Waters», США). Органические кислоты разделяли на колонке ACQUITY CSH C18 («Waters», США) и определяли на УФ-детекторе Photodiode Array ACQUITY UPLC («Waters», США) при X = 210 нм. Стандартами для идентификации компонентов корневых экссудатов служили свежеприготовленные смеси сахаров, органических кислот, непротеиногенных аминокислот (1-аминоциклопропан-1-карбоновая, p-аланин, а-аминомасляная, р-аминомасляная, у-аминомас-ляная, N-бутирил-DL-гомосерин лактон, L-канаванин, L-цитруллин, допамин, DL-гомосерин, D-глюкозамин, L-мимозин, L-орнитин, серотонин) (analytical grade, «Sigma-Aldrich Co.», США), L-триптофан (analytical grade, «Fluka Chemie GmbH», Швейцария) и гидролизат протеиногенных аминокислот Amino Acid Hydrolysate Standard H («Thermo Fisher Scientific, Inc.», США). Эксперименты выполняли в 3-кратной повторности.
Статистическую обработку результатов (определение стандартных ошибок, t -тест Стьюдента, НСР, корреляционный анализ) выполняли в программе STATISTICA v. 7.0 («StatSoft Inc.», США) и с помощью программы для дисперсионного анализа DIANA (16).
Результаты. У синтетической пшеницы, полученной скрещиванием твердой пшеницы (T. durum Desf.) сорта LEUCURUM 84693 (геном AB) и эгилопса (геном D), образовавшийся гексаплоидный генотип (геном ABD) воспроизводит полудикую пшеницу, существовавшую 4000-6000 лет назад, когда началось ее культивирование на Ближнем Востоке в области так назы- ваемого Плодородного полумесяца. Генотип обладает устойчивостью к абиотическим и биотическим стрессам благодаря генам, полученным с геномом D. Albostan (геном ABD) — местный древний традиционный примитивный сорт гексаплоидной пшеницы (T. aestivum L.), хорошо адаптированный к засухе, который культивировался в Турции сотни лет без улучшения современными селекционными методами. Сорт пшеницы (T. aestivum) Karahan (геном ABD), полученный стандартными селекционными методами, засухоустойчив и используется в Турции при создании сортов, устойчивых к засухе.
Вегетационный эксперимент показал, что у сорта Albostan диаметр корней был больше соответственно на 29 и 48 % в сравнении с синтетическим генотипом и сортом Karahan, а биомасса побегов и соотношение по-бег:корень у сортов Albostan и Karahan были выше, чем у синтетического генотипа, соответственно на 130 и 113 % и на 25 и 100 % (табл.).
Показатели роста корней и побегов у генотипов пшеницы, представляющих разные уровни эволюции культуры, при удалении и без удаления побегов на растении (вегетационный опыт)
|
Показатель в расчете на 1 растение |
Контрольные растения |
Растения с удаленным побегом |
|||||||
|
всего |
к контролю, % |
||||||||
|
1 |
2 |
1 3 |
1 |
1 2 |
3 |
1 1 |
2 1 |
3 |
|
|
Длина корней, см |
823a |
923a |
1373a |
1274a |
1404a |
1052a |
+55 |
+52 |
- 23 |
|
Площадь корней, см2 |
144a |
206a |
204a |
160a |
153a |
103a |
+11 |
- 26 |
- 50 |
|
Объем корней, см3 |
2,1ab |
3,7ab |
2,4ab |
1,6ab |
1,3ab |
0,8a |
- 22 |
- 64 |
- 67 |
|
Средний диаметр корня, мм |
0,55c |
0,71d |
0,48bc |
0,40ab |
0,35ab |
0,31a |
- 27 |
- 51 |
- 34 |
|
Число кончиков корней |
1596a |
2022a |
2311ab |
2365ab |
3398b |
2534ab |
+48 |
+68 |
+10 |
|
Число разветвлений корней |
6650a |
9723a |
11069a |
9529a |
10268a |
6299a |
+43 |
+6 |
- 43 |
|
Число пересечений корней |
926a |
1213a |
2121a |
1963a |
2318a |
1618a |
+112 |
+91 |
- 24 |
|
Сухая масса корней, г |
0,29ab |
0,53b |
0,31ab |
0,19a |
0,17a |
0,12a |
- 35 |
- 69 |
- 63 |
|
Сухая масса побега, г Отношение массы побега |
0,56a |
1,30b |
1,19b |
0,64a |
0,54a |
0,43a |
+15 |
- 59 |
- 64 |
|
к массе корня |
2,0a |
2,5ab |
4,0b |
3,5ab |
3,2ab |
3,7b |
+78 |
+32 |
- 8 |
|
Прим еч ани е. 1 — синтетическая форма, созданная скрещиванием твердой |
пшеницы |
( Triticum durum |
|||||||
|
Desf.) сорта украинского происхождения |
LEUCURUM 84693 (геном |
AB) и |
эгилопса |
Тауша ( Aegilops |
|||||
|
taushii Coss.) (геном D); 2 — |
традиционный сорт гексаплоидной пшеницы ( T. aestivum L.) Albostan (геном |
||||||||
ABD) местной турецкой селекции; 3 — засухоустойчивый современный сорт пшеницы (T. aestivum L.) Karahan (геном ABD) турецкой селекции. Приведены средние данные одного эксперимента в двух по- вторностях по каждому варианту. Неодинаковыми латинскими буквами отмечены достоверные различия между вариантами для каждого параметра (тест НСР, P < 0,05).
Удаление побега привело к уменьшению диаметра корней на 27-50 % у растений всех генотипов, к увеличению числа новых корней на 68 % у сорта Albostan, к снижению биомассы стеблей в 2,4 и 2,8 раза соответственно у сортов Albostan и Karahan, к снижению биомассы корней в 3,1 раза у сорта Albostan (см. табл.). Значения некоторых параметров корневой системы (длина корней, число корней, число ветвлений, отношение побег:корень) при удалении побега изменялись в зависимости от генотипа пшеницы, а именно возрастали у синтетической формы и растений сорта Albostan, но снижались у сорта Karahan (см. табл.). Эти данные можно рассматривать как свидетельство того, что даже у сортов современной селекции с эффективной корневой системой ее восстановление после стресса, вызванного удалением побега, все же затруднено. В контрольном варианте биомасса стеблей у современного сорта Karahan была в 4,0 раза выше, чем биомасса корней (см. табл.), тогда как у синтетической пшеницы и традиционного местного сорта Albostan — только соответственно в 2,0 и 2,5 раза. При удалении побега корневая система у синтетической пшеницы и сорта Albostan показала высокую эффективность в процессе ревегетации и способствовала приросту новой стеблевой биомассы до значений, которые в 3,2-3,5 раза превышали величину корневой биомассы. Сравнение средних значений показателей роста у подвергнутых стрессу и контрольных растений подтвердило, что эффект от удаления побега существенно зависит от генотипа пшеницы (рис. 1). У синтетической пшеницы по семи из десяти представленных в таблице параметров реакция растений на удаление стеблевой биомассы была положительной, у сорта Albostan под воздействием стресса возрастали значения пяти параметров, тогда как у современного сорта Karahan — только одного.
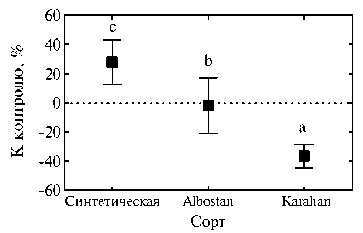
Сорт
Рис. 1. Средние значения ростовых параметров у растений пшеницы с удаленным побегом относительно контрольных (%). Изучены формы, представляющие разные уровни эволюции культуры: синтетическая, традиционный местный сорт Al-bostan и сорт современной турецкой селекции Karahan. Данные и описание сортов приведены в таблице. Вертикальные отрезки указывают стандартную ошибку средней. Неодинаковыми буквами отмечены достоверные различия между вариантами (тест НСР, P < 0,05).
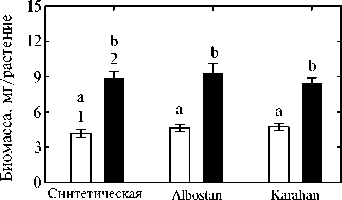
Рис. 2. Сухая биомасса корней (1) и побегов (2) у растений пшеницы в гидропонной культуре. Изучены формы, представляющие разные уровни эволюции культуры: синтетическая, традиционный местный сорт Albostan и сорт современной турецкой селекции Karahan. Приведены средние данные трех экспериментов в одной повторности по каждому варианту. Вертикальные отрезки указывают стандартную ошибку средней. Неодинаковыми буквами отмечены достоверные различия между вариантами (тест НСР, P < 0,05).
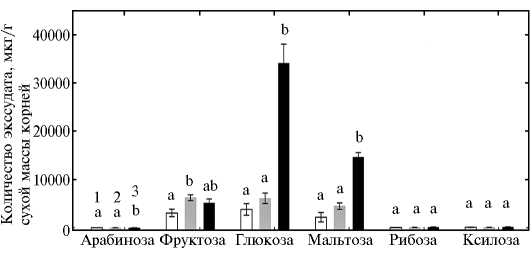
Рис. 3. Корневая экссудация сахаров у растений пшеницы в гидропонной культуре: 1, 2, 3 — формы, представляющие разные уровни эволюции культуры (соответственно синтетическая пшеница, традиционный местный сорт Albostan и сорт современной турецкой селекции Karahan). Приведены средние данные трех экспериментов в одной повторности по каждому варианту. Вертикальные отрезки указывают стандартную ошибку средней. Неодинаковыми буквами отмечены достоверные различия между вариантами (тест НСР, P < 0,05).
В условиях гидропоники изучаемые генотипы пшеницы практически не различались по биомассе корней или побегов (рис. 2), что позволяет предположить отсутствие влияния биомассы растений на интенсивность корневой экссудации. Анализ сахаров в корневых экссудатах показал, что у всех трех генотипов корни растений выделяли в основном фруктозу, глюкозу и мальтозу (рис. 3). Интенсивность экссудации сахаров была минимальной у синтетической пшеницы, а современный сорт Karahan секретировал существенно больше глюкозы и мальтозы, чем два других генотипа. В корневых экссудатах всех трех генотипов также обнаруживалось незначительное количество арабинозы, рибозы и ксилозы (см. рис. 3).
Состав органических кислот был сходным у всех генотипов (рис. 4). Основными компонентами оказались лимонная, молочная, яблочная и пропионовая кислоты. Генотипические различия выражались в повышенной корневой экссудации уксусной кислоты у растений синтетической пшеницы и пировиноградной кислоты у сорта Karahan в сравнении с сортом Albostan.
В корневых экссудатах изучаемых генотипов пшеницы были обна- ружены 17 протеиногенных аминокислот, основную долю которых составляли аргинин, аспарагиновая и глутаминовая кислоты, гистидин, лейцин, фенилаланин, серин, триптофан и валин (см. рис. 4), и только три из 13 анализируемых непротеиногенных аминокислот — α-аминомасляная, γ-амино-масляная кислоты и L-орнитин. Синтетическая пшеница отличалась от остальных форм повышенной экссудацией α-аминомасляной кислоты, аргинина и глицина, тогда как для современного сорта Karahan оказалось характерно интенсивное выделение гистидина, фенилаланина и триптофана.
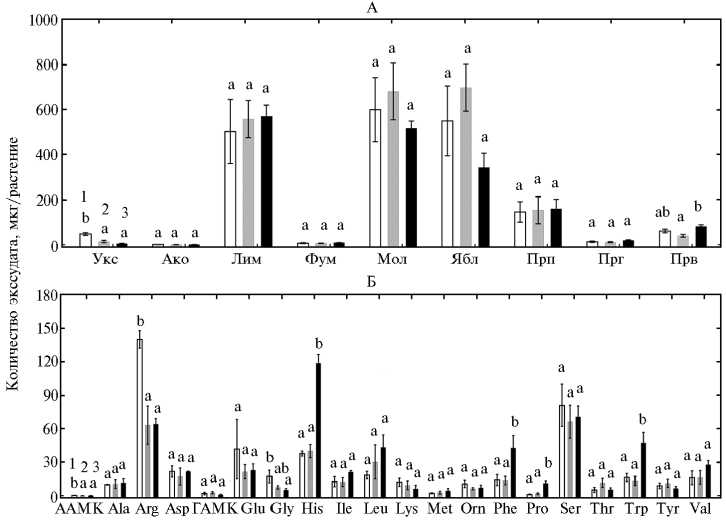
Рис. 4. Корневая экссудация органических кислот (А) и аминокислот (Б) у растений пшеницы в гидропонной культуре: 1, 2, 3 — формы, представляющие разные уровни эволюции культуры (соответственно синтетическая пшеница, традиционный местный сорт Albostan и сорт современной турецкой селекции Karahan); Укс, Ако, Лим, Фум, Мол, Ябл, Прп, Прг, Прв — соответственно уксусная, аконитовая, лимонная, фумаровая, молочная, яблочная, пропионовая, пироглутамовая, пировиноградная кислоты; ААМК и ГАМК — α -аминомасляная и γ -аминомасляная кислоты, для протеиногенных аминокислот указаны общепринятые аббревиатуры. Приведены средние данные трех экспериментов в одной повторности по каждому варианту. Вертикальные отрезки указывают стандартную ошибку средней. Неодинаковыми буквами отмечены достоверные различия между вариантами для каждой кислоты или аминокислоты (тест НСР, P < 0,05).
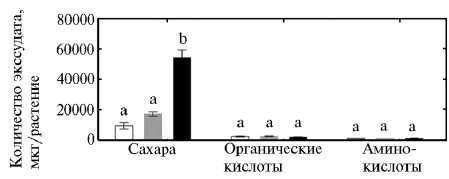
Рис. 5. Суммарное количество сахаров, органических кислот и аминокислот в корневых экссудатах у растений пшеницы в гидропонной культуре: 1, 2, 3 — формы, представляющие разные уровни эволюции культуры (соответственно синтетическая пшеница, традиционный местный сорт Albostan и сорт современной турецкой селекции Karahan). Приведены средние данные трех экспериментов в одной повторности по каждому варианту. Вертикальные отрезки указывают стандартную ошибку средней. Неодинаковыми буквами отмечены достоверные различия между вариантами (тест НСР, P < 0,05).
Суммарное количество сахаров, секретируемых корнями растений сорта Karahan, было соответственно в 5 и 3 раза выше, чем у синтетической пшеницы и сорта Albostan, но по суммарной экссудации органических ки- слот и аминокислот генотипических различий мы не выявили (рис. 5). Сравнение суммарных количеств в трех анализируемых фракциях корневых экссудатов показало, что у всех генотипов их основу составляют сахара.
Таким образом, основной исходной гипотезой при проведении вегетационного эксперимента стало предположение о том, что удаление побега растений представляет собой стресс, приводящий к мобилизации регенерационных процессов. При этом скорость и степень регенерации зависит от способности корневой системы снабжать растение водой и питательными элементами. Следовательно, более активное функционирование корневой системы, вероятно, приведет к ускоренной и эффективной регенерации растений. Эксперимент с удалением побега показал повышенную устойчивость к стрессу у синтетической пшеницы по сравнению с сортами Albostan и Karahan. Соответственно, эффективность регенерации, определяемая биомассой новых побегов и отношением параметров роста растений в стрессовом и контрольном вариантах, оказалась лучше у синтетической пшеницы. В то же время современный сорт Karahan в контрольном варианте продуцировал в 2 раза больше надземной биомассы в расчете на единицу массы корней. Можно предположить, что продолжительная селекция пшеницы косвенно способствовала повышению эффективности функционирования ее корневой системы. Наши результаты показали, что сорт Karahan был неспособен к увеличению отношения побег:корень в ответ на воздействие стрессового фактора. Более того, средняя величина этого отношения у подвергнутых стрессу и контрольных растений в случае сорта Karahan была значительно ниже, чем у синтетической пшеницы и сорта Albostan (см. рис. 1). Корневые системы синтетического генотипа и сорта Albostan проявили более высокую стабильность в ответ на стресс, вызванный удалением побега. Существует вероятность, что у этих генотипов засухоустойчивость выше, чем у сорта Karahan, и в настоящее время проводятся полевые опыты для проверки такого предположения. Ранее исследования, в которых сравнивали корневые системы старых и современных сортов пшеницы, показали, что у современных форм масса корней и размеры корневой системы в период цветения несколько меньше (17). Поедание побегов озимой пшеницы животными существенно не влияет на развитие корневой системы у зрелых растений (18). Однако сравнивать эти данные с результатами наших исследований затруднительно, так как мы анализировали параметры корневой системы до начала выхода в трубку. Не исключено, что эффективный ответ корневой системы синтетической пшеницы на удаление побега уникален и может быть связан с механизмами адаптации к абиотическим стрессам.
Значительные сортовые различия в корневой экссудации сахаров, органических кислот и аминокислот были описаны для разных видов растений, включая томат (19), горох (20) и картофель (21). Это указывает на высокую вариабельность генотипов по названным признакам. Характерной особенностью сорта Karahan оказалась очень активная экссудация сахаров (фруктозы, глюкозы и мальтозы), составлявших основное количество углеводов, выделяемых в ризосферу. У сорта Karahan, как уже отмечалось, оно соответственно в 5 и 3 раза выше, чем у синтетической пшеницы и сорта Albostan (см. рис. 5). Ранее нами было показано, что современные сорта гексаплоидной пшеницы (T. aestivum L.) выделяют сахара в ризосферу активнее, чем диплоидные генотипы T. boeoticum Boiss. и T. monococcum L. (22). На основании этих данных можно предположить, что современный сорт Karahan обладает низкой способностью контролировать интенсивность выделения сахаров через транспортные системы корней в окружающую среду. Правомерно также допустить, что при удалении побега высокая интенсивность экссудации сахаров у сорта Karahan способна привести к снижению транспорта источников углерода из корня в надземную часть и этим замедлять процесс ревегетации. Поэтому сравнение корневой экссудации у изучаемых сортов с удаленными побегами, а также количества и состава ксилемного сока, поступающего в регенерируемый побег, представляет интерес. В частности, у устойчивого к кадмию мутанта гороха SGECdt нами было показано усиление экссудации ксилемного сока (23) и активный транспорт питательных элементов из корня в побег (24), что приводило к лучшему росту растений при кадмиевом стрессе (23). Таким образом, усилением транспорта питательных и энергетических ресурсов из корней в побег может, по крайней мере частично, объясняться высокий ревегетационный потенциал синтетической пшеницы.
В представляемой работе мы впервые сравнили корневую экссудацию органических веществ (сахаров, органических и аминокислот) у генотипов пшеницы, соответствующих основным уровням ее эволюции. Наиболее важным результатом стало обнаружение существенных различий в интенсивности экссудации сахаров. Характер экссудации органических кислот у изученных сортов был сходен, но имелись некоторые генотипические особенности в экссудации аминокислот. Так, корни синтетической пшеницы выделяли больше аргинина, тогда как у современного сорта Karahan отмечали повышенную экссудацию гистидина, фенилаланина и триптофана. Триптофан известен как предшественник в биосинтезе ауксинов (25) — фитогормонов, играющих важную роль во многих физиологических процессах у растений, включая закладку и рост боковых корней, клеточное деление, формирование тканей ксилемы, регенерацию побегов и транспорт сахаров в меристему стеблей (26, 27). Потребность в триптофане и ауксинах возрастает в стрессовых условиях, вызванных потерей верхушки побега (в нашем эксперименте этот стресс был вызван удалением побегов), и эти соединения необходимы для снятия апикального доминирования и генерации нового побега (27). Активная экссудация триптофана у сорта Karahan может уменьшать его содержание в тканях, приводя к снижению биосинтеза ауксинов и их транспорта к регенерирующим побегам. Кроме того, дефицит ауксинов способен оказывать негативное воздействие на транспорт сахаров из корней в регенерирующие побеги. Для проверки этой гипотезы необходимы более детальные исследования, включая определение концентраций ауксинов и сахаров в тканях и ксилемном соке.
Итак, полученные результаты указывают на различия в функционировании корневой системы у трех изученных генотипов пшеницы (примитивный, современный и синтетический сорта), представляющих разные этапы эволюции гексаплоидной пшеницы, которые связаны с ревегетацией побега и корневой экссудацией. Выяснение природы обнаруженных различий требует дальнейшего изучения.
Список литературы Сравнительные характеристики корневых систем и корневой экссудации у синтетического, примитивного и современного сортов пшеницы
- Morgounov A., Haun S., Lang L., Martynov S., Sonder K. Climate change at winter wheat breeding sites in central Asia, eastern Europe, and USA, and implications for breeding. Euphytica, 2013, 194: 277-292 ( ) DOI: 10.1007/s10681-013-0968-1
- Blum A. Plant breeding for water-limited environments. Springer, 2011.
- White R.G., Kirkegaard J.A. The distribution and abundance of wheat roots in a dense, structured subsoil -implications for water uptake. Plant Cell and Environment, 2010, 33: 133-148 ( ) DOI: 10.1111/j.1365-3040.2009.02059.x
- Porcel R., Ruiz-Lozano J.M. Arbuscular mycorrhizal influence on leaf water potential, solute accumulation, and oxidative stress in soybean plants subjected to drought stress. Journal of Experimental Botany, 2004, 55: 1743-1750 ( ) DOI: 10.1093/jxb/erh188
- Aroca R., Porcel R., Ruiz-Lozano J.M. How does arbuscular mycorrhizal symbiosis regulate root hydraulic properties and plasma membrane aquaporins in Phaseolus vulgaris under drought, cold or salinity stresses? New Phytologist, 2007, 173: 808-816 ( ) DOI: 10.1111/j.1469-8137.2006.01961.x
- Belimov A.A., Dodd I.C., Hontzeas N., Theobald J.C., Safronova V.I., Davies W.J. Rhizosphere bacteria containing 1-aminocyclopropane-1-carboxylate deaminase increase yield of plants grown in drying soil via both local and systemic hormone signalling. New Phytologist, 2009, 181: 413-423 ( ) DOI: 10.1111/j.1469-8137.2008.02657.x
- Groppa M.D., Benavides M.P., Zawoznik M.S. Root hydraulic conductance, aquaporins and plant growth promoting microorganisms: a revision. Applied Soil Ecology, 2012, 61: 247-254 ( ) DOI: 10.1016/j.apsoil.2011.11.013
- Bais H.P., Weir T.L., Perry L.G., Gilroy S., Vivanco J.M. The role of root exudates in rhizosphere interactions with plants and other organisms. Annual Review of Plant Biology, 2006, 57: 233-266 (doi: 10.1146/annurev.arplant.57.032905.105159).
- Farrar J.F., Jones D.L. The control of carbon acquisition by roots. New Phytologist, 2000, 147: 43-53 ( ) DOI: 10.1046/j.1469-8137.2000.00688.x
- Plant roots. The hidden half/Y. Waisel, A. Eshel, U. Kafkafi (eds.). NY, Marcel Dekkers Inc., 1996.
- Blum A., Ritchie J.T. Effect of soil surface water content on sorghum root distribution in the soil. Field Crops Research, 1984, 8: 169-176 ( ) DOI: 10.1016/0378-4290(84)90060-1
- Asseng S., Ritchie J.T., Smucker A.J.M., Robertson M.J. Root growth and water uptake during water deficit and recovering in wheat. Plant and Soil, 1998, 201: 265-273 ( ) DOI: 10.1023/A:1004317523264
- Reynolds M., Mujeeb-Kazi A., Sawkins M. Prospects for utilising plant-adaptive mechanisms to improve wheat and other crops in drought-and salinity-prone environments. Annals of Applied Biology, 2005, 146: 239-259 ( ) DOI: 10.1111/j.1744-7348.2005.040058.x
- Bell L.W., Harrison A.M., Kirkegaard B. Dual-purpose cropping -capitalising on potential grain crop grazing to enhance mixed-farming profitability. Crop and Pasture Science, 2015, 66: i-iv.
- Ogbonnaya F.C. Development, management and utilization of synthetic hexaploids in wheat improvement. In: The world wheat book: a history of wheat breeding/A.P. Bonjean, W.J. Angus, M. van Ginkel (eds.). Lavoisier, France, 2011, V. 2: 823-849.
- Воробьев Н.И., Проворов Н.А., Свиридова О.В. Программа для однофакторного дисперсионного анализа рендомизированных биологических данных. Свидетельство о государственной регистрации программы для ЭВМ ¹ 2013615092. М., 2013.
- Siddique K.H.M., Belford R.K., Tennant D. Root:shoot ratios of old and modern, tall and semi-dwarf wheats in a mediterranean environment. Plant and Soil, 1990, 121: 89-98 ( ) DOI: 10.1007/BF00013101
- Kierkegaard J.A., Lilley J.M., Hunt J.R., Sprague S.J., Ytting N.K., Rasmussen I.S., Graham J.M. Effect of defoliation by grazing or shoot removal on the root growth of field-grown wheat (Triticum aestivum L.). Crop and Pasture Science, 2015, 66: 249-259.
- Шапошников А.И., Азарова Т.С., Кравченко Л.В., Бажанова А.А., Бажанов Д.П., Бабак О.Г., Некрашевич Н.А., Кильчевский А.В. Корневые выделения генотипов томата (Solanum lycopersicum L.), отличающихся отзывчивостью на бактеризацию. Молекулярная и прикладная генетика, 2012, 14: 63-68.
- Кузмичева Ю.В., Шапошников А.И., Азарова Т.С., Петрова С.Н., Наумкина Т.С., Борисов А.Ю., Белимов А.А., Кравченко Л.В., Парахин Н.В., Тихонович И.А. Состав корневых экзометаболитов высокосимбиотрофного сорта гороха Триумф и его родительских форм. Физиология растений, 2014, 61(1): 121-128 ( ) DOI: 10.7868/S0015330314010084
- Belimov A.A., Dodd I.C., Safronova V.I., Shaposhnikov A.I., Azarova T.S., Makarova N.M., Davies W.J., Tikhonovich I.A. Rhizobacteria that produce auxins and contain ACC deaminase decrease amino acid concentrations in the rhizosphere and improve growth and yield of well-watered and water-limited potato (Solanum tuberosum). Annals of Applied Biology, 2015, 1: 11-25 ( ) DOI: 10.1111/aab.12203
- Кравченко Л.В., Шапошников А.И., Макарова Н.М., Азарова Т.С., Львова К.А., Костюк И.И., Тихонович И.А. Видовые особенности состава корневых выделений растений и его изменение в ризосфере под влиянием почвенной микрофлоры. Сельскохозяйственная биология, 2011, 3: 71-75.
- Belimov A.A., Dodd I.C., Safronova V.I., Malkov N.V., Davies W.J., Tikhonovich I.A. The cadmium tolerant pea (Pisum sativum L.) mutant SGECdt is more sensitive to mercury: assessing plant water relations. Journal of Experimental Botany, 2015, 66(8): 2359-2369 ( ) DOI: 10.1093/jxb/eru536
- Tsyganov V.E., Belimov A.A., Borisov A.Y., Safronova V.I., Georgi M., Dietz K.-J., Tikhonovich I.A. A chemically induced new pea (Pisum sativum L.) mutant SGECdt with increased tolerance to and accumulation of cadmium. Annals of Botany, 2007, 99: 227-237 ( ) DOI: 10.1093/aob/mcl261
- Ljung K. Auxin metabolism and homeostasis during plant Development. Development, 2013, 140, 5: 943-950 ( ) DOI: 10.1242/dev.086363
- Sassi M., Vernoux T. Auxin and self-organization at the shoot apical meristem. Journal of Experimental Botany, 2013, 64: 2579-2592 ( ) DOI: 10.1093/jxb/ert101
- Motte H., Vereecke D., Geelen D., Werbrouck S. The molecular path to in vitro shoot regeneration. Biotechnology Advantages, 2014, 32: 107-121 ( ) DOI: 10.1016/j.biotechadv.2013.12.002
