Сравнительный анализ функций населения ногохвосток лесной подстилки

Автор: Пинаева О.Н.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Наземные экосистемы
Статья в выпуске: 2 т.10, 2008 года.
Бесплатный доступ
В статье освещены некоторые функциональные особенности влияния жизнедеятельности ногохвосток на трансформацию органического вещества лиственного опада в зависимости от ценотической орга- низации населения коллембол.
Короткий адрес: https://sciup.org/148198110
IDR: 148198110 | УДК: 595.713:595.423
Contrastive analysis functions of population springtails carpet of leaves
This article deals with the problem of some functional perculiarities of springtails influence on the transformation of organic substance of leaf tall, which depends on the cenotic organization of the inhabitance of collembols.
Текст научной статьи Сравнительный анализ функций населения ногохвосток лесной подстилки
Российский государственный университет туризма и сервиса, г. Самара
В статье освещены некоторые функциональные особенности влияния жизнедеятельности ногохвосток на трансформацию органического вещества лиственного опада в зависимости от ценотической организации населения коллембол.
Лесная подстилка населена разнообразным и стабильным на основе сложных внутренних связей сообществом микроорганизмов и животных. Интегральный эффект воздействия этого сообщества напрямую сказывается на характеристиках лесной почвы, находящихся в прямой зависимости от скорости и направления биотических круговоротов.
Микроартроподы, и коллемболы в первую очередь, являются активными участниками процессов трансформации органического вещества растительных остатков. Высокая насыщенность растительного опада микроарт-роподами ставит вопрос об их роли в биоценозах. Многочисленные расчеты и эксперименты показывают, что на долю микроарт-ропод приходится относительно небольшая часть энергии, заключенная в химических связях веществ растительного опада [1]. Однако, исключение их деятельности резко снижает скорость разложения опада [2].
Микроартроподы, как представители второго гетеротрофного трофического уровня, выступают регуляторами микробиальной активности [3] [4], что влияет на соотношение процессов минерализации и гумификации [5]. Микробиологическая обстановка растительного опада, его положение в подстилке, структура сообщества микроартропод должны сказываться на продуктивности и стабильности лесных экосистем [6].
Раздельное влияние коллембол различных жизненных форм на процесс трансформации растительного опада
В стеклянные сосуды объемом 1 литр на чистый речной песок, промытый водой, HCl, NaOH и еще раз водой, помещали по 10 г (ВСВ) смесь лиственного опада (дуб, липа, лещина).
Лиственный опад подвергался температурной обработке (60-700С в течение 1-2 часов) для уничтожения кладок и диапазирую-щих беспозвоночных. Опад увлажняли до 60 % от полной его влагоемкости. Варианты эксперимента предусматривали деструкцию опада в следующих режимах: а) при участии микроорганизмов;
-
б) при участии микроорганизмов и видов ногохвосток, относящихся к пионерной группе мелких членистоногих – Entomobrya sp., Isotoma viridis;
-
в) при участии микроорганизмов и видов постпионерной группы – Isotoma gr. olivacea, помещенных в опад спустя 4 месяца с начала эксперимента;
-
г) при участии микроорганизмов и видов ферментативного слоя лесной подстилки -Isotoma notabilis, Folsomia quadrioculata, Pseudosinella alba, инокулированных спустя 6 месяцев с начала эксперимента;
-
д) при участии микроорганизмов и видов почвенного горизонта - Folsomia fimetaria, Onychiurus gr. armatus, помещенных в опад с 10 месяцев с начала эксперимента;
-
е) при участии микроорганизмов и комплекса всех указанных выше видов ногохвосток.
Опад инокулировали коллемболами, извлекая их из естественной подстилки методом флотации с последующим переносом насекомых в соответствующие сосуды препа- ровальными иглами. Начальная численность ногохвосток в вариантах – 10 экз/г опада.
Микробиальная обстановка лиственного опада на различных этапах деструкции ока- зывает существенное влияние на интенсивность физиологических процессов ногохвосток, что отразилось на динамике их численности в экспериментальном опаде (рис. 1).
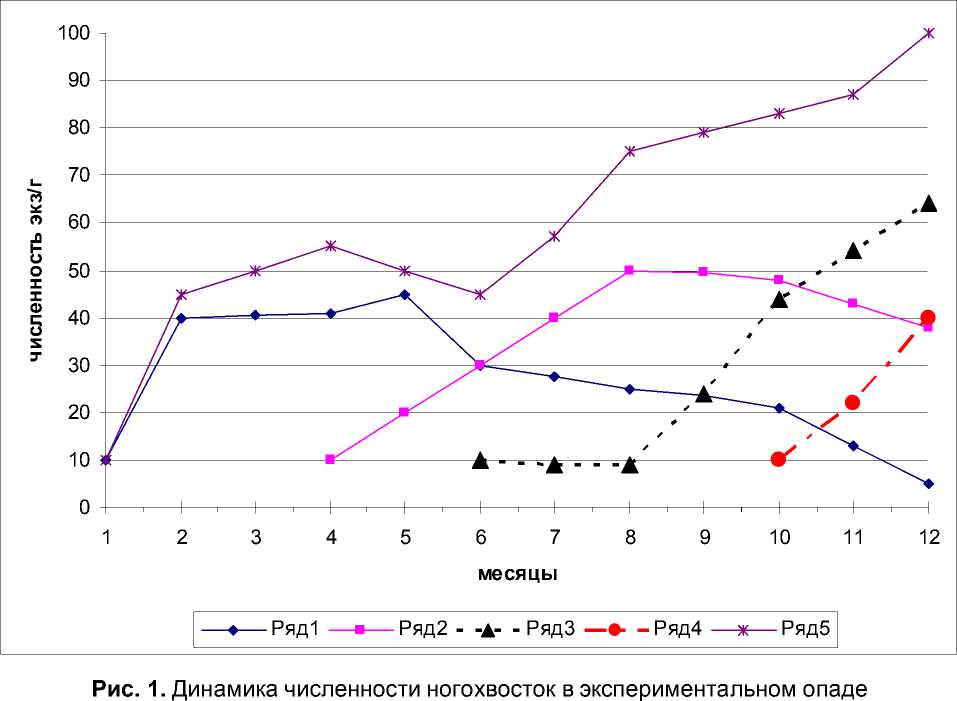
Суммарная численность Entomobrya sp.и Isotoma viridis имела максимальное значение на 2 – 4 месяцы, что вполне соответствует положению атмобионтных видов начальных этапов сукцессии населения коллембол лиственного опада. В дальнейшем обилие ногохвосток данной жизненной формы неуклонно снижалось и к концу эксперимента упало до 5 экз.
Отсутствие в экспериментальном опаде предыдущих группировок ногохвосток (по вариантам эксперимента) негативно сказывалось на интенсивности размножения видов последующих экологических групп коллем-бол. Так максимальное обилие Isotoma gr. olivacea отмечено на 8 – 10 месяцы с последующим спадом численности, гемиэдафичес-кие виды имели некоторое снижение численности к 8 месяцу с последующим ростом чис- ленности на 10 и 12 месяцы экспозиции, а эудафические виды начали увеличивать свою численность с момента инокуляции.
Из приведенных результатов видно, что микробиальная обстановка экспериментального опада оказывает наибольшее влияние на гемиэдафические виды, которые являются основными регуляторами микробиальной активности и от которых в большей степени зависит соотношение процессов минерализации и гумификации органического вещества растительных остатков.
Кроме того, в экспериментальных микрокосмах не сработало одно из основных правил видовой структуры сообщества: чем меньше видов, тем больше численность. Вспышка численности отельных видов в обедненных группировках не отмечена.
Комплексная группировка ногохвосток (ва- риант е) функционировала в режиме, обычном для достаточно сложных и устойчивых сообществ микроартропод [1].
Темпы размножения коллембол оказывали специфическое воздействие на скорость разложения опада (рис. 2).
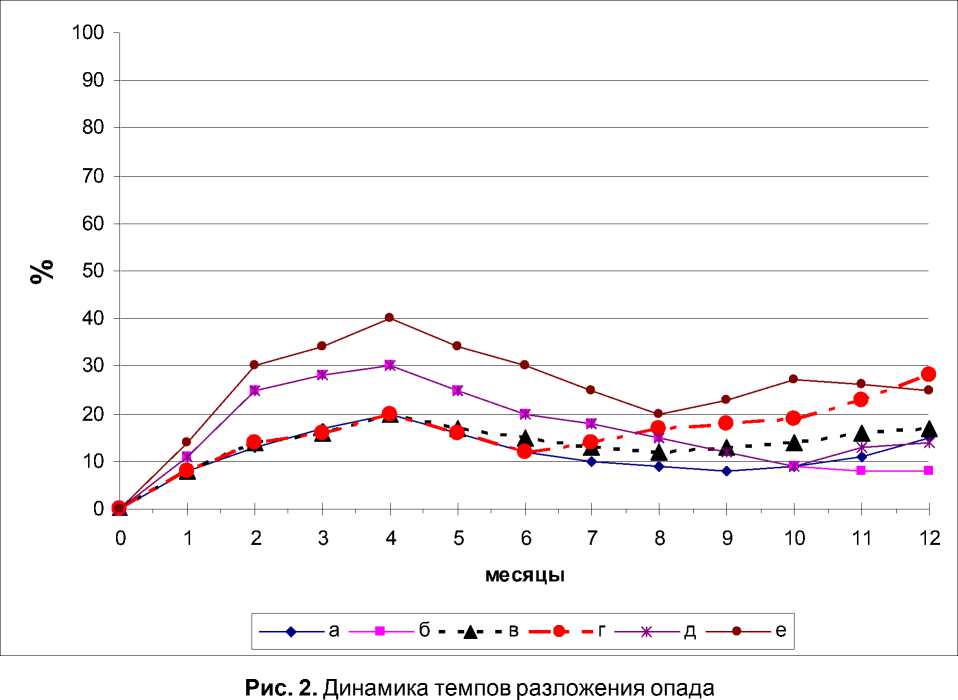
Кривые темпов разложения опада во всех вариантах эксперимента, кроме полного комплекса ногохвосток, повторяли по форме таковые динамики численности коллембол экспериментального опада (рис.1). Более значительная скорость разложения растительных остатков в вариантах с ногохвостками указывает на их оптимизирующую роль процесса деструкции [7].
Отсутствие начальной и последующих группировок ногохвосток в экспериментальном опаде в вариантах опыта приводит к тому, что темпы потери веса опадом увеличиваются к концу эксперимента в отличие от варианта с полным комплексом коллембол.
Общеизвестно, что темпы разложения растительных остатков постепенно падают. В лабораторных экспериментах кривые темпов разложения с участием полного комплекса микроартропод имеют 2 пика [7], характери- зующие интенсивность и последовательность преобладающих процессов минерализации и гумификации.
Органические вещества опада, находящиеся под воздействием неполных группировок ногохвосток, и как следствие, в несвойственных условиях деструкции (степень активности микроорганизмов, видовой состав микроорганизмов и микроартропод, и т.д.), не разлагаются с той интенсивностью, которая отмечается в вариантах с полным комплексом микроартропод. Эти «невостребованные» вещества начинают интенсивно разлагаться при инокуляции опада последующей группой ногохвосток, что и сказывается на повышении темпов разложения к концу эксперимента.
Таким образом, нарушенные группировки коллембол, оказываясь в несвойственных им условиях (степень разложения опада, видо- вой состав микроорганизмов и микроартро-под) не могут в полной мере компенсировать отсутствие тех видов ногохвосток, которые должны предшествовать им согласно сукцессионной смены видов первичнобескрылых насекомых.
Различия в условиях деструкционных про- цессов и в степени влияния разрозненных экологических групп ногохвосток на эти процессы должны некоторым образом сказаться на соотношении процессов минерализации и гумификации органического вещества лиственного опада.
Таблица 1. Содержание органического углерода и соотношение ГК/ФК в пирофосфатных вытяжках из листьев
|
№ |
Биоагенты |
2 |
4 |
6 |
8 |
10 |
12 |
||||||
|
С орг |
ГК/ ФК |
С орг |
ГК/ ФК |
С орг |
ГК/ ФК |
С орг |
ГК/ ФК |
С орг |
ГК/ ФК |
С орг |
ГК/ ФК |
||
|
1 |
Микроорганизмы |
5,2 |
0,6 |
5,1 |
0,7 |
5,5 |
0,8 |
6,9 |
1,1 |
7,8 |
1,1 |
9,6 |
1,7 |
|
2 |
Микроорганизмы и пионерная группа |
6,7 |
1,1 |
6,3 |
1,3 |
6,1 |
1,5 |
7,1 |
1,2 |
6,6 |
1,2 |
9,1 |
1,1 |
|
3 |
Микроорганизмы и постпионерная группа |
7,1 |
1,2 |
7,4 |
1,3 |
8,5 |
1,3 |
10,5 |
1,5 |
||||
|
4 |
Микроорганизмы и гемиэдафическая группа |
8,5 |
1,5 |
10,3 |
1,7 |
11,2 |
2,1 |
||||||
|
5 |
Микроорганизмы и эудафическая группа |
9,0 |
1,1 |
12,1 |
1,3 |
||||||||
|
6 |
Микроорганизмы и комплекс микроартропод |
7,21 |
1,6 |
9,0 |
2,5 |
10,1 |
1,9 |
14,8 |
2,6 |
15,5 |
2,8 |
16,1 |
3,1 |
Анализируя данные таблицы 1 можно сделать следующие выводы:
-
1. Деятельность микроартропод в комплексе с микроорганизмами положительным образом сказывается на процессах трансформации органического вещества лесного опада.
-
2. Деятельность нарушенных группировок ногохвосток, представленных 1-2 видами каждой экологической группы в раздельных вариантах эксперимента, отрицательно сказывается на течение процессов гумификации и минерализации по сравнению с полным комплексом ногохвосток.
-
3. Деятельность отдельных видов ногохвосток в процессе деструкции лиственного опада всецело обусловлена подготовительной деятельностью микроорганизмов, т.е. соответствующей микробиологической обстановкой растительных остатков.
-
4. Отсутствие видов-предшественников в поэтапном процессе деструкции лиственного опада негативно сказывается на микробиальной активности опада и на процессе
-
5. Выход ГВ в варианте с микроорганизмами в целом соответствует данным, имеющимся в литературе [1].
-
6. Позитивная роль пионерной группировки ногохвосток наиболее ощутимо отмечена лишь в конце годичного срока экспозиции.
-
7. Интенсивность влияния ногохвосток на соотношение процессов минерализации и гумификации усиливается в направлении от верхнеподстилочных к эудафическим.
трансформации органического вещества в целом.
Трансформация органического углерода растительных остатков при нарушении сукцессионной смены видов ногохвосток
Снижение видового разнообразия микро-артропод снимает зависимость пространственного распределения от этапа разложения органического субстрата.
Однако, горизонтальная и вертикальная стратификация видов в обедненных группировках мелких членистоногих не может не сказаться на механизме множественного обеспечения функций комплекса микроартро-под в деструкции опада.
В модельных опытах изучали влияние различных видов ногохвосток, относящихся к разным экологическим группам, на процесс деструкции при нарушении хода сукцессионной смены видов.
Подготовка экспериментальных сосудов аналогична предыдущему опыту.
Варианты эксперимента предусматривали трансформацию органического вещества лиственного опада при участии:
-
а) микроорганизмов
-
б) микроорганизмов и пионерной группы ногохвосток - Entomobrya sp., Isotoma viridis;
-
в) микроорганизмов, пионерной и постпионерной группы - Isotoma gr. olivacea; последняя группировка помещена в опад спустя 4 ме-
- сяца с начала эксперимента;
-
г) микроорганизмы, пионерная и гемиэдафичес-кая группа - Isotoma notabilis, Folsomia quadrioculata, Pseudosinella alba; последняя группировка инокулирована с 6 месяцев с начала эксперимента;
-
д) микроорганизмы, пионерная и эудафическая группа - Folsomia fimetaria, Onychiurus gr. armatus; последняя группировка помещена в опад с 10 месяцев с начала эксперимента;
-
е) микроорганизмы и комплекс указанных выше видов ногохвосток.
Нарушение микробиальной активности опада, связанное с выпадением отдельных видов ногохвосток в сукцессионном ряду сказалось на динамике численности коллембол (рис. 3).
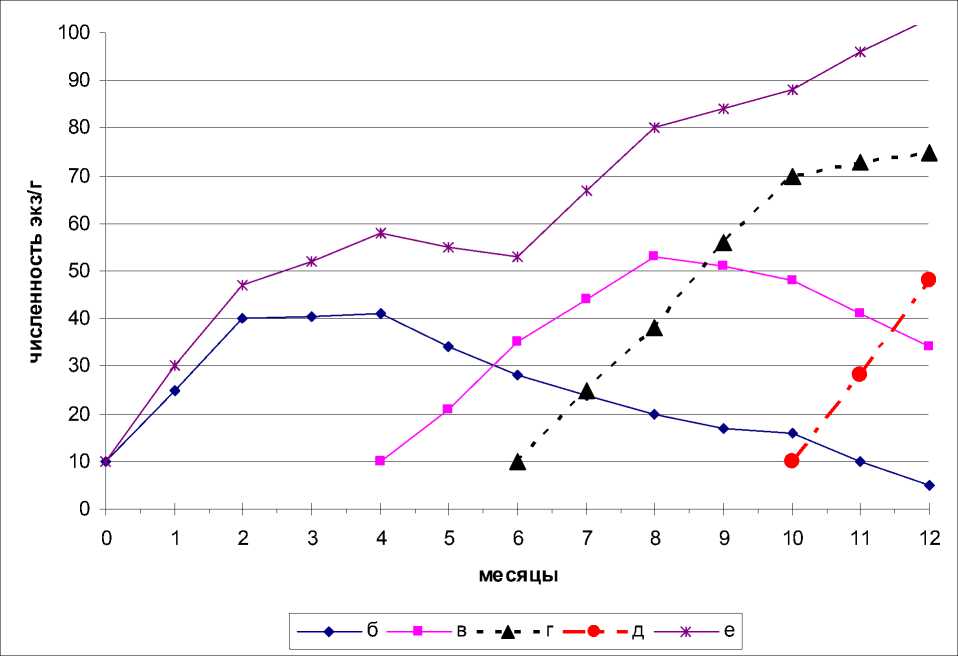
Рис. 3. Динамика численности отдельных видов ногохвосток и их общей численности в вариантах эксперимента
Кривая динамики численности пионерной группировки аналогична изменениям численности ногохвосток предыдущего эксперимента (см. рис.1).
Ход кривых динамики численности дру гих экологических групп разнятся от подоб ных кривых тех же экологических групп пре дыдущего эксперимента (рис. 1 и рис. 3). Раз личия касаются не столько сроков максимальных значений численности, сколько количества особей соответствующей экологической группы.
Наиболее существенное различие в численности ногохвосток в сравнении с предыдущим вариантом отмечены в варианте с ге-миэдафическими видами.
Несмотря на то, что деятельность пионерной группировки создала комфортную микробиологическую обстановку для постпионерной группы, численность последней неуклонно снижается.
Численность видов эудафической группы выше, чем в предыдущем эксперименте, од- нако отсутствие видов-предшественников тормозило сукцессионную смену микроорганизмов, что негативно сказалось на степени «готовности» опада к деятельности настоящепочвенных видов.
В целом, выпадение видов-предшественников отрицательно сказалось на динамике численности ногохвосток по сравнению с таковыми показателями полного комплекса коллембол.
Изменения в динамике численности ногохвосток и микробиологической активности лиственного опада сказалось и на темпах разложения (рис. 4).
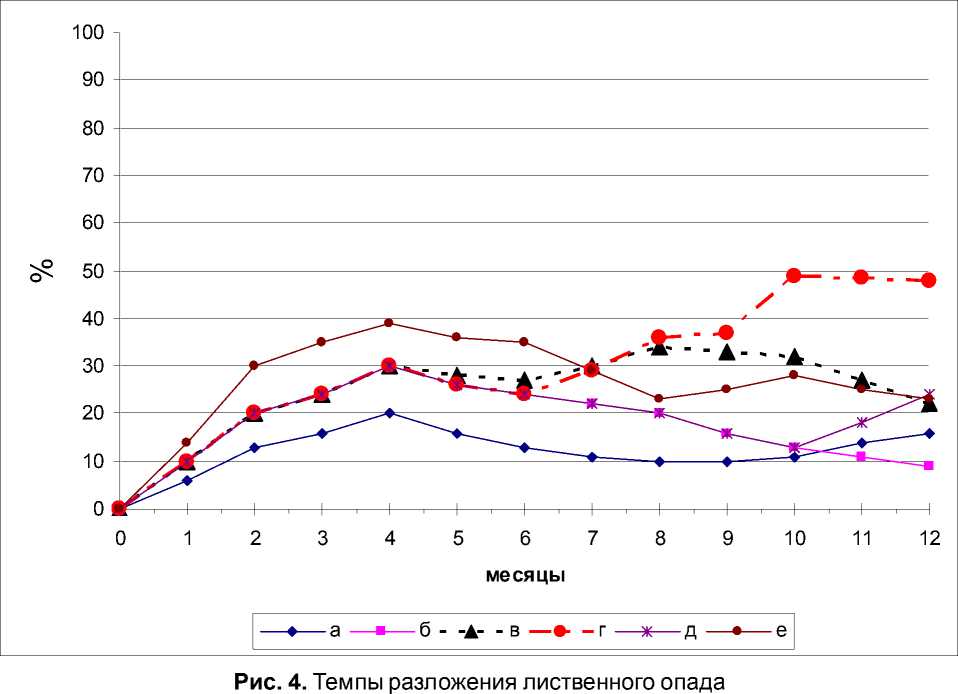
Темпы разложения опада в варианте с микроорганизмами аналогичны предыдущим экспериментам: в течение года максимум наблюдался на 3-4 месяце экспозиции, а второй пик активности отмечается к 12 месяцу.
Деятельность пионерной группировки увеличивает по значению, но не смещая по времени 1 волну потери органического вещества лиственным опадом по сравнению с микроорганизмами. Однако, отсутствие ногохвосток других жизненных форм, влияющих на микробиальную активность, делают жизненную обстановку не вполне комфортной, что сказывается на снижении темпов разложения к концу эксперимента.
Сравнительная деятельность пионерной и постпионерной группировок начиная с 4 месяцев, дает «типичный» [7] второй пик ак- тивности, приходящийся на 8 – 10 месяцы деструкции опада.
Совершенно очевидно, что последовательная деятельность двух экологических сукцессионных групп позитивно сказывается на увеличении темпов разложения растительных остатков.
Деятельность гемиэдафической группы ногохвосток, подключенной к пионерной группе с 6 месяцев так же положительно сказывается на динамике темпов деструкции: появление второго пика приурочено к 8 – 10 месяцу, однако его значение гораздо выше предыдущего. Это говорит о том, что деятельность основной группы микроартропод в гу-мификационном процессе переоценить невозможно.
Наконец, иная динамика темпов разложения опада регистрируется при совокупном воздействии начальной и эудафической групп: отсутствие предыдущих групп микроартро-под, не свойственность микробиальной обстановки срокам экспозиции негативно сказывается на деятельности почвенных видов – темпы разложения лиственного опада «нео- хотно» взрослеют.
Как было показано ранее [7] 1 пик темпов разложения характеризует минерализирующие процессы, тогда как второй пик «гуми-фикационный», он ниже первого и характеризует степень участия микроартропод в процессах накопления гумусовых веществ.
Во всех вариантах эксперимента, где участвовали две экологические группы ногохвосток, отмечено увеличение темпов разложения, т.е. второй пик, вопреки логике, выше чем в варианте с полным комплексом ногохвосток. Очевидно, это объясняется тем, что в обедненных сообществах деструкторов процесс разложения идет не полноценно, и на долю включающихся экологических групп приходится значительная остаточная часть легкоразлагающегося органического вещества, а то время как в варианте с полным набором экологических групп коллембол процесс минерализации и гумификации протекает по известным законам.
Подтверждение сказанному мы регистрируем в данных по выходу и накоплению ГВ (табл. 1).
Таблица 1. Содержание С орг. Растительных остатков в пирофосфатной вытяжке (мгС/г опада)
|
№ |
Варианты |
Сроки (месяцы) |
|||||
|
2 |
4 |
6 |
8 |
10 |
12 |
||
|
1 |
Микроорганизмы (контроль) |
0,6 |
0,7 |
0,8 |
1,1 |
1,1 |
1,7 |
|
2 |
Начальная с 0 месяцев |
1,1 |
1,3 |
1,5 |
1,2 |
1,2 |
1,1 |
|
3 |
Начальная+постпионерная с 4 месяцев |
1,1 |
1,2 |
1,4 |
1,3 |
1,5 |
1,3 |
|
4 |
Начальная+гемиэдафическая с 6 месяцев |
1,1 |
1,2 |
1,5 |
1,5 |
1,8 |
2,3 |
|
5 |
Начальная+эудафическая с 10 месяцев |
1,1 |
1,2 |
1,5 |
1,4 |
1,4 |
1,3 |
|
6 |
Полный комплекс |
1,6 |
2,5 |
1,9 |
2,6 |
2,6 |
3,1 |
Как следует из анализа кривых динамики выхода и накопления ГВ совместная деятельность двух экологических групп ногохвосток положительно сказывается на процессах гумификации органического вещества: во всех вариантах эксперимента отмечено увеличение выхода С орг гумусовых веществ (рис. 5).
Деятельность пионерной и постпионерной группировки характеризуется спокойным течением процесса деструкции: увеличении выхода ГВ происходило постепенно, что обусловлено отсутствием увеличением поздних функциональных групп ногохвосток.
Деятельность пионерной и гемиэдафичес-кой групп значительно увеличивало выход ГВ, однако кривая динамики также спокойная, что отражает неполный механизм поэтапного включения органического вещества растительных остатков в процессах минерализации и гумификации.
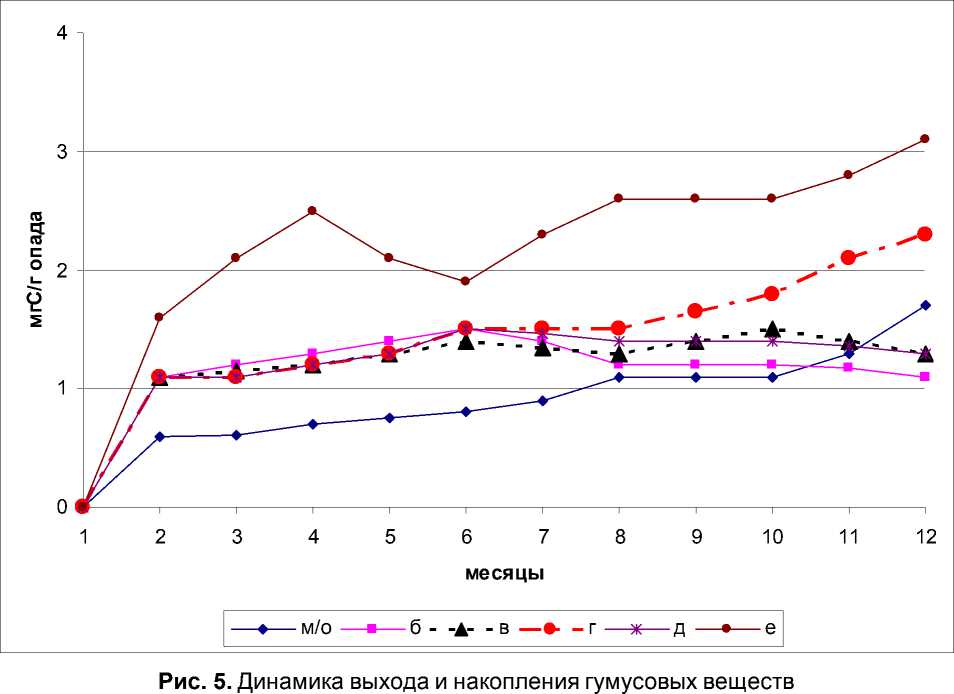
Кривая динамики выхода ГВ в варианте с эудафической группой ногохвосток указывает, что опад не готов к «приему» настоящей почвенной группировки, т.к. в течение 10 месяцев отсутствовали практически все промежуточные экологические группировки но- гохвосток.
Список литературы Сравнительный анализ функций населения ногохвосток лесной подстилки
- Симонов Ю.В. Раздельное влияние коллембол различных жизненных форм на процесс гумификации растительных остатков.//Изучение и охрана биологического разнообразия ландшафтов. Пенза, 1999.
- Паников Н.С., Симонов Ю.В. Влияние микроартропод на скорость разложения растительного опада.//Экология, № 4. 1986.
- Симонов Ю.В., Борисова В.Н. Экспериментальный анализ взаимоотношения микроартропод с гифомицетами лесной подстилки.//Экология микроартропод лесных почв. М.: Наука, 1988.
- Симонов Ю.В., Добровольская Т.Г. Воздействие коллембол и орибатид на бактериальные клетки разлагающегося опада//Экология, № 5. 1994.
- Симонов Ю.В. Количественная оценка участия микроартропод в гумификации растительных остатков//Докл. АнСССР, 1984. Т.277. № 4.
- Чернова Н.М., Кузнецова Н.А., Симонов Ю.В. Ценотическая организация и функции населения микроартропод лесной подстилки.//Механизмы биотической деструкции органического вещества в почве. М.: Наука, 1989.
- Симонов Ю.В. Общие закономерности влияния микроартропод на трансформацию органического вещества почвы.//Исследования в области биологии и методики ее преподавания: Межкафедральный сборник научных трудов. Вып.1. Самара: Издательство СГПУ, 2002.
