Сравнительный анализ структуры подпологовых популяций ели в березняках южной тайги и смешанных лесов Русской равнины

Автор: Дерюгин А.А., Глазунов Ю.Б.
Журнал: Лесохозяйственная информация @forestry-information
Рубрика: Лесоведение и лесоводство
Статья в выпуске: 2, 2019 года.
Бесплатный доступ
Приведены результаты изучения демутационного процесса в 105-летних березняках с подпологовой популяцией ели в условиях хвойно-широколиственного и южно-таежного лесных районов. Рассмотрены возрастная и вертикальная структуры популяции ели, сделан вывод об отсутствии принципиальных различий в развитии подпологовой популяции ели в районах исследований.
Южная тайга, хвойно-широколиственные леса, березняки, ель под пологом, структура древостоя
Короткий адрес: https://sciup.org/143169920
IDR: 143169920 | УДК: 630.182.21: | DOI: 10.24419/LHI.2304-3083.2019.2.02
Comparative analysis of the under-canopy spruce populations structure in birch forests in southern taiga subzone and mixed forests zone of the Russian plain
Researches of recovery processes in natural post-disturbance birch forests with under-canopy spruce population are actual in connection with an increase in the area of such stands, a decrease in the volume of tending felling in young stands, and insufficient knowledge of the process at the later stages of development of birch forests. The purpose of this study is to make a comparative analysis of the under-canopy spruce populations in birch forests, the age of which approaches age of natural maturity, for the conditions of the southern taiga and the mixed forests. The objects of research are birch forests at the age of 105 years with a under-canopy population of spruce. The works were carried out on permanent trial plots in the Moscow region, Mozhaisk district, and in the Yaroslavl region, Rybinsk district. For all the spruce trees age was determined, biometric characteristics were measured, the presence of rot was determined selectively when taking cores. It is established that the regeneration of spruce under the canopy of birch forests in the southern taiga and coniferous-broadleaved forest areas occurs according to similar scenarios...
Текст научной статьи Сравнительный анализ структуры подпологовых популяций ели в березняках южной тайги и смешанных лесов Русской равнины
Для ссылок:
Дерюгин, А. А. Сравнительный анализ структуры подпологовых популяций ели в березняках южной тайги и смешанных лесов Русской равнины [Электронный ресурс] / А. А. Дерюгин, Ю. Б. Глазунов // Лесохоз. информ. : электрон. сетевой журн. – 2019. – № 2. – С. 15–24. URL:
Современное состояние земель лесного фонда в южно-таёжном и хвойно-широколиственном (смешанном) лесных районах европейской части России характеризуется значительным участием (около 30% площади земель, занятых лесными насаждениями) производных березовых насаждений. За 2003–2013 гг. в рассматриваемых регионах площадь таких насаждений возросла почти на 4%, что обусловлено, главным образом, сокращением объемов рубок ухода в молодых насаждениях березы. За 10-летний период в смешанных лесах площадь рубок ухода уменьшилась почти на 60%, в южной тайге – на 30%. На значительной части площади под пологом березняков развивается популяция ели. В южной тайге доля таких березняков может достигать 75%, в смешанных лесах – 52% [1]. В таких насаждениях демутационные процессы протекают спонтанно. Изучение этих процессов с целью прогнозирования будущих древостоев и оценки их хозяйственной ценности является актуальной проблемой.
Восстановительно-возрастная динамика подпологовой ели в березняках европейской части России была объектом внимания многих исследователей [2-5]. Однако в большинстве работ результативность этого процесса оценивается гипотетически. Наиболее полно демутационные процессы в березняках с популяцией ели под пологом изучены в южной тайге [6–8].
Цель настоящей работы – провести сравнительный анализ периода возобновления, возрастной и вертикальной структур подпологовых популяций ели в березняках, возраст которых приближается к возрасту естественной спелости для условий южной тайги и смешанных лесов. В березняках такого возраста можно определить перспективу формирующихся ельников.
Объекты и методика исследований
Исследования выполнены Институтом лесоведения РАН в южно-таёжном лесном районе (Ярославская обл., Рыбинский район, координаты: 58°14’038’’ N, 38°57’513’’ E и в хвойно-широколиственном (смешанном) лесном районе (Московская обл.,, Можайский район, координаты: 55°40’746’’ N, 35°31’390’’ E). Объект исследований – подпологовая популяция ели в производных березняках. Работы проводили на постоянных пробных площадях (ПП), заложенных в высокополнотных, Ia и I классов бонитета, березняках-кисличниках 105-летнего возраста:
ПП-1БС – район хвойно-широколиственных (смешанных) лесов, Московская обл. – Поречье;
ПП-17 – южно-таежный район, Ярославская обл. – Косково.
По составу первого и второго ярусов древостоев ПП отличаются несущественно. Некоторые различия наблюдаются в густоте подроста и производительности древостоев. Последнее связано с географическим расположением пробных площадей и расхождением в возрасте (около 20 лет) деревьев ели первого яруса (табл. 1).
Различия таксационных характеристик насаждений на постоянных ПП не исключают возможности проведения сравнительного анализа результата динамики подпологовой популяции ели в березняках, находящихся в конце стадии «старения», т.е. в возрасте естественной спелости [8]. Следует отметить, что вся ель под пологом – это последующее возобновление, появившееся после рубки коренных ельников.
На ПП установлены возраст, биометрические характеристики, выполнены работы по картированию каждого дерева; при взятии кернов определяли наличие гнили. В процессе обработки данных и анализа использованы методы, описанные в ранее опубликованных работах [7, 9, 10].
Результаты и обсуждение
В южной тайге и смешанных лесах основная часть подпологовой популяции ели возобновляется в березняках, достигших 20–30-летнего возраста [11–14]. Это регламентирует возрастной диапазон популяции – около 30 лет. О том, что в условиях южной тайги восстановительный период ели после рубки насаждений может
Таблица 1. Таксационная характеристика насаждений на пробных площадях
Однако некоторые исследования возрастной структуры подпологовой ели свидетельствуют о ее более широком возрастном диапазоне. Так, по данным В. И. Алексеева, в сомкнутых лесах Подмосковья подрост ели под пологом лиственных насаждений, возникших на вырубках, представлен непрерывным возрастным рядом от 3–5 до 50–60 лет [17].
Возобновление ели под пологом березняков в южной тайге может проходить в ранние и поздние сроки. При раннем возобновлении, наиболее распространенном в регионе, формирование популяции ели происходит, главным образом, за счет особей, появившихся в березняках до 30-летнего возраста. При отсутствии генеративных деревьев поселение ели под пологом березы может наблюдается в более поздние сроки [8].
Процесс возобновления подпологовой еловой популяции может характеризоваться периодом возобновления (Tw) ели, сохранившейся под пологом березняков. Величина этого периода определяется разницей между средним возрастом березы и возрастом ели. В настоящей публикации рассматриваются березняки, относящиеся к насаждениям с ранним возобновлением ели.
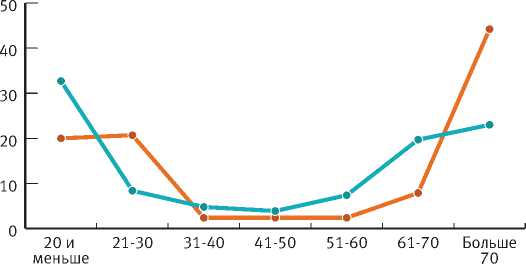
На рассматриваемых объектах распределение деревьев ели по периоду возобновления однотипно и имеет вид U-образной кривой, на которой можно выделить три области (рис. 1).
Первая область (левая ветвь) кривой включает 30-летний период, в течение которого появляется значительная часть деревьев ели. На ПП-1БС и ПП 17 на это возрастное поколение ели приходится примерно равная доля (около 40%) общей численности популяции.
Вторая область (центральная часть) кривой охватывает период возобновления 31–60 лет. Как показали исследования в южной тайге [8], к этому времени численность подпологовой популяции ели достигает максимальных значений. В березняках в возрасте 50 лет начинает формироваться второй ярус ели, обостряется внутривидовая конкуренция. Особи ели, появившиеся в это время, попадают под полог первого поколения (Tw≤30 лет) ели и большей частью погибают. Это объясняет небольшое участие деревьев этого поколения (Tw=31–60 лет) в популяции: ПП-1БС – 16%, ПП-17 – 7%.
В процессе формирования второго яруса ели наблюдается значительный отпад елового подроста, сокращается общая численность популяции. Через 70–80 лет после возобновления березы под ее пологом создаются условия для выживания ели с периодом возобновления больше 70 лет. Происходит пополнение популяции ели (правая часть) – третья область кривой. В популяции доля поколения с периодом возобновлении Tw>60 лет составляет около 43% в Московской обл. (ПП-1БС) и 52% в Ярославской обл. (ПП-17).
Доля в популяции, % Доля в популяции, %
TW, лет
Ярославская обл., ПП-17
Московская обл., ПП-1БС
1. Распределение деревьев ели по периодам возобновления
Возраст ели, лет
Ярославская обл., ПП-17
Московская обл., ПП-1БС
Рис.
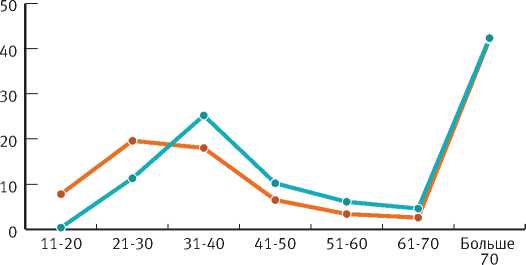
2. Возрастное распределение деревьев ели
Период возобновления определяет возрастную структуру подпологовой популяции ели. Возрастная дифференциация деревьев, как и распределение по Tw, на объектах исследований практически однотипны (рис. 2). Наиболее представлены деревья в возрасте более 70 лет (ПП-1БС – 42%, 471 шт./га; ПП-17 – 41%, 608 шт./ га). Это поколение, появившееся в березняках до 30-летнего возраста, является основой формирующегося ельника и определяет перспективу его развития. В вертикальной структуре двух изучаемых объектов деревья этого возраста относятся к первому и второму ярусам древостоя. В формировании этих ярусов небольшое участие принимают деревья 61–70-летнего возраста с Tw=35-45 лет. Деревья других возрастных групп относятся к подросту и преимущественно находятся в угнетенном состоянии.
Распределение деревьев по периоду возобновления и возрасту на объектах практически идентично, о чем свидетельствуют коэффициенты асимметрии и эксцесса (табл. 2). Они не отвечают закону нормального распределения (критерии достоверности Тex>3). Эти распределения в том и другом случае могут быть аппроксимированы уравнением вида: lny=a+bx+cx2 +dx3 +ex4.
Оценка типа возрастной структуры подполо-говой популяции ели, проведенная по значению стандартного отклонения возрастных рядов (G) [18], показала, что ельники, формирующиеся под пологом 105-летних березняков в условиях смешанных лесов и южной тайги можно отнести к условно одновозрастным (G=28 лет).
Для оценки сопоставимости изучаемых объектов проанализирована вертикальная структура подпологовой популяции ели. Средние характеристики ярусов древостоев свидетельствуют о схожести объектов (см. табл. 1). Это подтверждается данными о высотном распределении деревьев ели. На определенную тождественность распределения деревьев на ПП по высотным группам указывают почти равные значения коэффициентов и критериев достоверности асимметрии и эксцесса. Ряды распределения отличаются лишь максимумами и амплитудами высоты (см.
Таблица 2. Статистические характеристики рядов распределения
32,1-34,0
^■•^* Ярославская обл., ПП-17
Московская обл., ПП-1БС
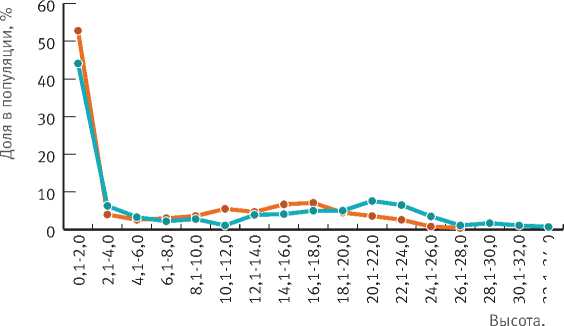
Рис. 3. Распределение деревьев ели по высотным группам
м
На обеих ПП наиболее распространены деревья высотой до 2 м. Их доля достигает 44% на ПП-1БС и 52% на ПП-17 (рис. 3). Различие в представленности деревьев других высотных групп не превышает 4,5% (высотные группы деревьев второго яруса 10,1–12,0 и 20,1–22,0 м).
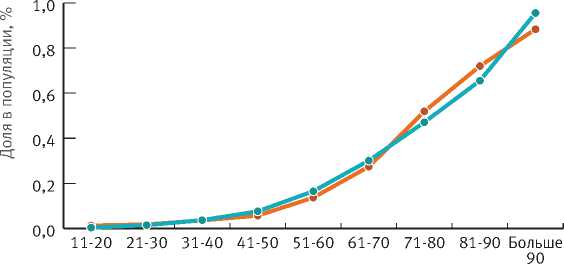
Кроме того, для каждой 10-летней возрастной группы нами рассчитано среднее значение относительной высоты: отношение средней высоты в возрастной группе к средней высоте деревьев ели 100-летнего возраста в нормальных насаждениях II класса бонитета (рис. 4). Данный подход обусловлен тем, что такого возраста достигли деревья – лидеры подпологовой популяции, а ельники такой производительности наиболее распространены в регионах исследований.
Возраст ели, лет
Ярославская обл., ПП-17
—*^— Московская обл., ПП-1БС
Рис. 4. Изменение относительной высоты деревьев ели в зависимости от возраста
Установлено, что кривые, отображающие связь относительной высоты с возрастом деревьев подпологовой ели, для южной тайги и смешанных лесов практически одинаковы. Это в очередной раз свидетельствует о схожести процессов формирования ельников в изучаемых березняках.
В регионах исследований к одному из негативных явлений, влияющих на формирование ельников подпологовой популяции, относится поражение деревьев стволовой и комлевой гнилями.
По данным исследований, в южной тайге доля таких деревьев в первом и втором ярусах с возрастом увеличивается. Поражаемость гнилью деревьев старше 75-летнего возраста может достигать 54% [19]. На изучаемых объектах в составе первого и второго ярусов доля таких деревьев составляет 11% (60 шт./га) в Московской обл. (ПП-1СБ) и 21% (120 шт./га) в Ярославской обл. (ПП-17). Эти деревья в сильной мере подвержены ветролому и ветровалу, поэтому численность части популяции, возобновившейся в первые 30 лет после поселения березы, уменьшается, а также снижается возможность смены березы первым поколением ели с периодом возобновления до 30 лет. После распада березняков из этого поколения с высокой долей вероятности сформируются низкополнотные III–IV классов бонитета ельники. Восстановление коренных ельников возможно за счет поколения ели с периодом возобновления более 80 лет.
Выводы
Возобновление ели под пологом березняков в южно-таежном и хвойно-широколиственном лесных районах происходит по схожим сценариям. Основа формирующихся под пологом березы ельников – поколение, появившееся в березняках до 30-летнего возраста. Другое представительное поколение ели появляется при достижении березняками возраста более 70 лет.
К началу распада березняков (120 лет) под-пологовые еловые популяции в обоих лесных районах характеризуются примерно одинаковыми возрастной и вертикальной структурами.
Несмотря на различия в абсолютных значениях некоторых показателей, вызванные климатическими условиями произрастания, де-мутационный процесс в березняках южной тайги и смешанных лесов протекает по одинаковым закономерностям, обусловленным, прежде всего, биологическими особенностями и совместным развитием популяций березы и ели. Климатические условия имеют при этом второстепенное значение.
В условиях южной тайги и смешанных лесов после распада березняков с высокой долей вероятности можно спрогнозировать формирование низкополнотных и низкопроизводительных ельников.
Список литературы Сравнительный анализ структуры подпологовых популяций ели в березняках южной тайги и смешанных лесов Русской равнины
- Писаренко, А. И. Лесовосстановление/А. И. Писаренко. -М.: Лесн. пром-сть, 1977. -256 с.
- Мелехов И. С. Лесоведение./И. С. Мелехов. -М.: Лесн. пром-сть, 1980. -408 с.
- Побединский, А. В. Рубки главного пользования/А. В. Побединский. -М.: Лесн. пром-сть, 1980. -192 с.
- Таёжное лесоводство/В. Г. Чертовской, И. С. Мелехов, Г. В. Крылов . -М.: Лесн. пром-сть, 1974. -232 с.
- Орлов, А. Я. Почвенно-экологические основы лесоводства в южной тайге/А. Я. Орлов. -М.: Наука, 1991. -104 с.
- Рубцов, М. В. Возрастная структура популяции ели под пологом березняков южной тайги/М. В. Рубцов, А. А. Дерюгин, А. П. Никитин//Лесоведение. -2000. -№ 4. -С. 28-34.
- Рубцов, М. В. Возрастная динамика морфоструктуры и рост популяции ели под пологом березняков южной тайги/М. В. Рубцов, А. А. Дерюгин//Идеи биогеоценологии в лесоведении и лесоразведении. -М.: Наука, 2006. -С. 63-81.
- Рубцов, М. В. Динамика возрастной структуры популяции ели под пологом южно-таёжных березняков Русской равнины/М. В. Рубцов, А. А. Дерюгин//Хвойные бореальной зоны. -2013. -XXXI. -№ 1-2. -С. 9-14.
- Рубцов, М. В. Возрастная динамика морфоструктуры и состояния популяции ели под пологом южнотаёжных березняков/М. В. Рубцов, А. А. Дерюгин//Комплексные стационарные исследования в лесах южной тайги (Памяти М. В. Рубцова). -М.: Тов-во науч. изд. КМК, 2017. -С. 125-153.
- Рубцов, М. В. Восстановительно-возрастная динамика популяции ели под пологом южно-таёжных березняков при демутационном процессе/М. В. Рубцов, А. А. Дерюгин//Продукционный процесс и структура лесных биогеоценозов: теория и эксперимент (Памяти А. И. Уткина). -М.: Тов-во науч. изд. КМК, 2009. -С. 206-228.
- Белов, С. В. Лесоводство/C. В. Белов. -М.: Лесн. пром-сть, 1983. -352 с.
- Чупров, Н. П. Зональные особенности возрастной структуры березово-еловых древостоев Европейского Севера/Н. П. Чупров//Лесоведение. -1988. -№ 3. -С. 11-20.
- Яковлев, Г. В. Особенности последующего возобновления ели в подзоне южной тайги/Г. В. Яковлев//Возобновление и формирование лесов на вырубках: сб. ст. -М., 1975. -С. 120-130.
- Яруткин, И. А. Оптимальный возраст лиственных древостоев для возобновления ели обыкновенной в подзоне хвойно-широколиственных лесов/И. А. Яруткин//Вопросы повышения продуктивности лесов. -Йшкар-Ола: Марийское кн. изд-во, 1976. -С. 10-13.
- Кравчинский, Д. М. О последствиях сплошных рубок в еловых и лиственных лесах средней и северной тайги/Д. М. Кравчинский. -СПб, 1901. -31 с.
- Кайрюкштис, Л. Формирование елово-лиственных молодняков/Л. Кайрюкштис. -Каунас, ЛитННИЛХ, 1959. -245 с.
- Алексеев, В. И. Возобновление ели на вырубках/В. И. Алексеев. -М.: Наука, 1978. -130 с.
- Лебков, В. Ф. Дендрометрические основы структурно-динамической организации древесных ценозов сосны: автореф. дисс. в форме науч. докл. … д-ра биол. наук; 06.03.03./В. Ф. Лебков. -М.: ИЛ РАН, 1992. -43 с.
- Рубцов, М. В. Поражение гнилью ели в процессе формирования её популяции под пологом южно-таёжных березняков/М. В. Рубцов, А. П. Никитин, А. А. Дерюгин/Строение, свойства и качество древесины -2004: тр. Междунар. симп.; 13-16 октября 2004, СПбГА. -Т. II. -СПб, 2004. -С. 456-457.
