Сравнительный фенетический анализ популяций двух видов грызунов (Apodemus, Mus) в условиях Центрального Кавказа
 в условиях Центрального Кавказа")
Автор: Амшокова Альбина Хасмановна, Кучинова Екатерина Александровна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Общая биология
Статья в выпуске: 5-2 т.19, 2017 года.
Бесплатный доступ
Исследована изменчивость 20 неметрических признаков черепа 197 экземпляров малой лесной и 204 экземпляров домовой мыши из 4 географических точек Центрального Кавказа с учетом двух факторов - высоты местности и антропогенного загрязнения. Выявлена видоспецифическая реакция на действие, как антропогенного загрязнения, так и фактора высоты, что проявляется в несходной картине изменения частот фенов у исследуемых видов грызунов. По величине фенетических дистанций, установлено, что антропогенные и контрольные, а также среднегорные и предгорные выборки малой лесной мыши дифференцированы в большей степени, чем таковые домовой мыши. Предполагается, что меньший уровень дифференциации выборок домовой мыши обусловлен более сглаженными условиями существования и, как следствие, меньшей подверженностью средовым факторам. В качестве второй возможной причины может быть и то, что популяции домовой мыши в большей степени подвержены генетическому обмену за счет периодического завоза зверьков из других районов, что вероятно приводит к нивелировке эпигенетических различий.
Малая лесная мышь, домовая мышь, неметрические признаки черепа, предгорья, среднегорья, фенетические дистанции, центральный кавказ
Короткий адрес: https://sciup.org/148205361
IDR: 148205361 | УДК: 575.2
Comparative phenetical analysis of populations of two rodent species (Apodemus, Mus) in the Central Caucasus
Variation of 20 nonmetrical cranium characters in 197 specimens of the lesser common field mouse and in 204 specimens of the house mouse from 4 geographical sites of the Central Caucasus have been studied considering two factors: anthropogenic pollution and altitude of locality. A species-specific reaction to the effect of anthropogenic pollution and the altitude of locality factor was revealed. This is reflected in a dissimilar pattern of changes in the frequency of phenes in the studied rodent species. Based on phenetic distances, it is found that the anthropogenic and control, as well as middle highland and foothill samples of a field mouse are distinguished largely than those of a house mouse. It is suggested that the lesser differentiation level in the samples of the house mouse is caused by more favourable living conditions, and therefore by lesser susceptibility to the environmental factors. The second possible reason may be as follows: the populations of the house mouse are more susceptible to genetic exchange through periodic invasion from other regions, which probably leads to a leveling of epigenetic differences.
Текст научной статьи Сравнительный фенетический анализ популяций двух видов грызунов (Apodemus, Mus) в условиях Центрального Кавказа
Работа выполнена при частичной финансовой поддержке гранта РФФИ №15-04-03981.
Изучение состояния природных популяций живых организмов, находящихся под воздействием комплекса естественных и антропогенных факторов среды является одной из важнейших задач экологии. Во многих регионах Центрального Кавказа основной формой антропогенной деятельности выступает сельскохозяйственная трансформация территорий, связанная с интенсивным применением минеральных удобрений, ядохимикатов и т.д. В связи с этим исследование особенностей морфологической изменчивости мелких млекопитающих, обитающих на антропогенных территориях весьма актуально и представляет интерес для понимания адаптивных стратегий организма в меняющихся условиях среды.
Для оценки состояния популяций животных широко используются методы феногенетического мониторинга, основанные на изучении эпигенетических и феногенетических особенностей сравниваемых групп, индивидуальных и групповых характеристик морфогенеза. Как отмечают А.Г. Васильев и И.А. Васильева [1], феногенетический мониторинг включает в себя оценку состояния среды как в пространстве (выявление размаха феногенетических различий
между населением различных местообитаний), так и во времени (определение изменений в состоянии населения животных в одном местообитании в течение ряда лет.
Цель – на основе анализа неметрических признаков черепа провести сравнительную оценку влияния двух факторов – антропогенного загрязнения и высоты местности на популяции широко распространенных видов грызунов A. uralensis и М. musculus .
МАТЕРИАЛ И МЕТОДЫ
Материал по обоим видам собирался одновременно в одни и те же годы в предгорьях Центрального Кавказа: окр. г. Нальчик – 500 м над ур. м.; окр. с. Псынадаха – 700 м над ур. м.; окр. с. Бедык – 1000 м над ур. м. Первая точка относится к естественным биотопам, а последние две к антропогенным (агроценозы). Для оценки влияния фактора высоты местности в анализ включена дополнительная выборка из среднегорий Центрального Кавказа – п. Эльбрус – 1800 м над ур. м. Объем изученного материала составил по малой лесной мыши –197, а по домовой мыши – 204 экз. черепов зверьков.
При проведении фенетического анализа (выделении, описании фенов, обработке результатов) использовали методы, разработанные и предложенные А.Г. Васильевым [2, 3]. Поиск и учет фенов проводили для каждого вида отдельно с помощью микроскопа Carl Zeiss (Stemi
2000C). Большая часть фенов, используемых в работе, взята из литературных источников [2, 4, 5]. За основу принята латинизированная система кодировки фенов, разработанная И.А. Васильевой [6]. Связь проявления фенов с полом, возрастом, друг с другом оценивали на основе расчета непараметрических коэффициентов корреляции Спирмена. Признаки, проявившие сильную значимую корреляцию с указанными факторами, исключили из дальнейшего анализа, что уменьшает вероятность получения артефактов за счет смещения оценок, обусловленных названными причинами, и повышает надежность косвенной генетической интерпретации фенетических различий [3].
Учитывая, что часть признаков, характерных для малой лесной мыши, имела иное состояние признака у домовой мыши, с целью сопоставимости данных фенетический анализ проводили по 20 неметрическим признакам черепа, встречающимся у обоих видов. Перечень 20 признаков, оставшихся после первичной выбраковки, приведен в таблицах 1, 3, а их расположение показано на рисунке 1.
Фенетические дистанции (MMD) между выборками и их средние стандартные отклонения (MSD) рассчитывали по формулам, предложенным С. Хартманом [7]. Различия статистически значимы на уровне р < 0.05 при MMD > 2 MSD. Меры средней фенетической уникальности (MMU) для
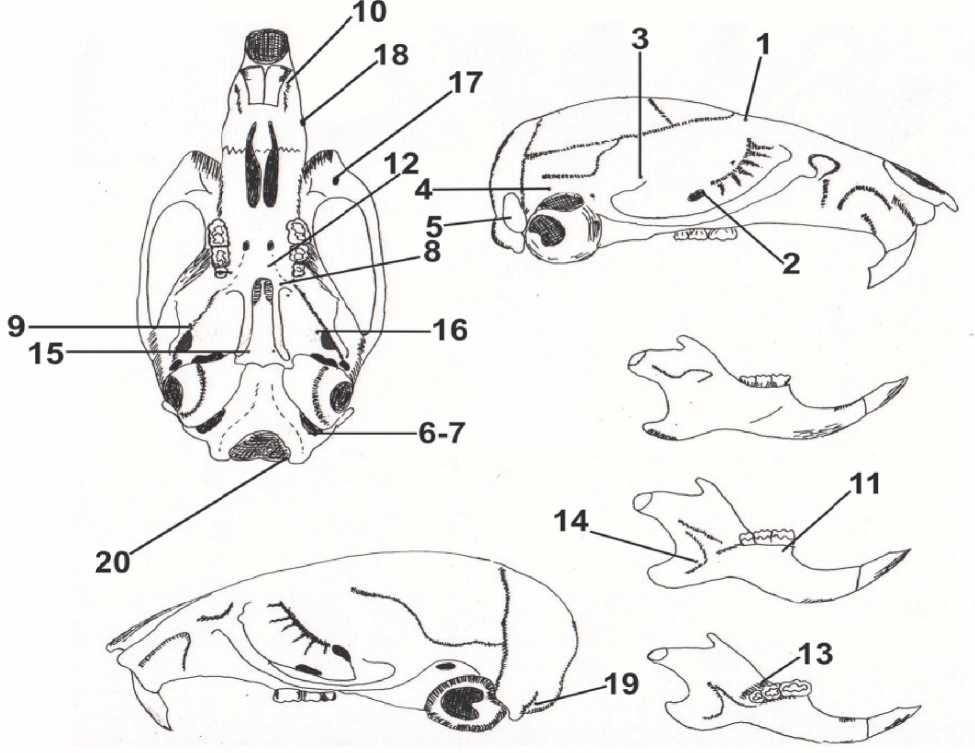
Рис. 1. Расположение фенов неметрических признаков на черепе малой лесной мыши:
1 – отсутствие лобного отверстия FFracan; 2 – одиночное нижнеглазничное отверстие FIO; 3 – одиночное переднее отверстие чешуйчатой кости FTmacan; 4 – одиночное отверстие височного хода MeTm; 5 – отсутствие окна на сосцевидной кости FeMs; 6 – одиночное подъязычное отверстие FHg; 7 – утроенное подъязычное отверстие FHgrt; 8 – одиночное отверстие в основании крыловидного отростка FPT; 9 – удвоенное отверстие на перегородке между овальным и круглым отверстиями FLTIdu; 10 – переднее вентральное предчелюстное отверстие FPmvean; 11 – удвоенное отверстие с лингвальной стороны альвеолы М1 FMtlgdu; 12 – одиночное дополнительное небное отверстие FPl; 13 – удвоенное отверстие с лингвальной стороны альвеолы М3 FPCdu;
14 – дополнительное отверстие в крыловидной ямке нижней челюсти (в сочленовно-угловой вырезке) FMbmst; 15 – боковые отверстия на вентральной поверхности основной клиновидной кости FBsla;
16 – дополнительные отверстия в области овального отверстия с вентральной стороны в крыловидной ямке Fasac; 17 – боковое верхнечелюстное отверстие на скуловом отростке FMxzmla; 18 – переднее боковое верхнечелюстное отверстие FPmla; 19 – множественные отверстия на затылочной кости в сочленовной ямке FFsOC (mx);
20 – наличие мыщелкового отверстия FCS
каждой выборки подсчитывали как сумму MMD данной выборки со всеми остальными, отнесенную к числу сравниваемых пар выборок [8].
Уровень внутрипопуляционного разнообразия оценивали с помощью показателя μ по Л.А. Животовскому [9].
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Первым этапом было проведено сравнение предгорных выборок малой лесной мыши из антропогенных и естественных биотопов. При сравнении контрольной (г. Нальчик) и антропогенной выборок (с. Псынадаха) различия выявлены по трем неметрическим признакам черепа: FIO, Fasac, FPmvean, из которых встречаемость первых двух выше в псынадахской выборке.
Аналогичное сравнение зверьков из контрольной и загрязненной территории (Наль-чик–Бедык) выявило достоверные различия по семи признакам, из которых четыре – FTmacan, FPT, FPmvean, FBsla значимо чаще встречаются в нальчикской, а три – FMbmst, Fasac и FFsOc(mx) в бедыкской выборке. Между парой выборок Псы-надаха–Бедык достоверные различия выявлены по частоте встречаемости 4 признаков из 20, что составляет 20% от изученного числа признаков (табл.1). Частота встречаемости отмеченных признаков в большинстве случаев выше в псы-надахской выборке, за исключением признака – дополнительное отверстие в крыловидной ямке нижней челюсти FMbmst, встречающегося с высокой частотой в бедыкской выборке.
В таблице 2 приведены фенетические дистанции, рассчитанные для отмеченных пар выборок малой лесной мыши. Дистанции между всеми сравниваемыми парами: Нальчик – Псынадаха, Нальчик – Бедык, Бедык – Псынадаха, оказываются статистически высоко достоверными.
При этом максимальная дистанция получена между выборками Бедык – Нальчик (0,056±0,007; d.f.=19; p<0,001), промежуточное положение занимают значения MMD между выборками Бедык – Псынадаха (0,040±0,006; d.f.=19; p<0,001) и минимальные значения отмечены между выборками Псынадаха – Нальчик (0,021±0,007; d.f.=19; p<0,01).
Результаты сравнения антропогенных и контрольных выборок домовой мыши в условиях предгорий отличны от полученных при сравнении таковых малой лесной мыши. Так, выборки домовой мыши из Бедыка и Нальчика различаются только по 1 признаку (табл.3).
Между псынадахской и бедыкской выборками достоверные различия выявлены по трем признакам – FHg, FBsla, Fasac, из которых
Таблица 1. Частота встречаемости фенов неметрических признаков черепа в выборках малой лесной мыши Центрального Кавказа, %
|
Номер и код признака |
1 |
2 |
3 |
4 |
1-2 |
1-3 |
2-3 |
1-4 |
2-4 |
3-4 |
|
N=82 |
N=94 |
N=10 8 |
N=11 0 |
Хи - квадрат |
||||||
|
1. FFracan (-) |
1,22 |
2,1 |
1 |
1,8 |
||||||
|
2. FIO |
65,9 |
84,0 |
60,2 |
75,2 |
||||||
|
3. FTmacan |
9,8 |
6,4 |
2,8 |
9,1 |
||||||
|
4. MeTm |
14,6 |
24,5 |
16,7 |
11,8 |
* |
|||||
|
5. FeMs (-) |
7,3 |
10,6 |
12,0 |
29,4 |
||||||
|
6. FHg |
40,2 |
50,0 |
52,9 |
41,8 |
||||||
|
7. FHgtr |
2,44 |
5,3 |
5,7 |
4,6 |
||||||
|
8. FPT |
28,1 |
32,3 |
7,4 |
10,9 |
*** |
*** |
** |
*** |
||
|
9. FLTIdu |
18,3 |
28,7 |
27,8 |
42,7 |
||||||
|
10. FPmvean |
14,6 |
5,3 |
1 |
12,0 |
||||||
|
11. FMtlgdu |
0 |
2,1 |
0 |
2,7 |
||||||
|
12. FPl |
46,3 |
36,2 |
40,7 |
31,5 |
||||||
|
13. FPCdu (M 3 ) |
40,2 |
28,7 |
26,9 |
33,3 |
||||||
|
14. FMbmst |
0 |
1,1 |
6,5 |
7,41 |
||||||
|
15. FBsla |
53,7 |
54,3 |
33,3 |
41,8 |
||||||
|
16. Fasac |
23,2 |
39,4 |
39,8 |
60,9 |
||||||
|
17. FMxzmla |
0 |
0 |
0 |
1 |
||||||
|
18. FPmla |
26,8 |
33,3 |
26,9 |
25,5 |
||||||
|
19. FFsOc(mx) |
15,9 |
27,7 |
29,6 |
46,4 |
||||||
|
20. FCS |
55,6 |
59,6 |
54,7 |
64,6 |
||||||
Примечание: 1 – г. Нальчик, 2 – с. Псынадаха, 3 – с. Бедык, 4 – п. Эльбрус; N-число изученных сторон черепа. Уровень значимости межгрупповых различий по отдельным признакам: * p<0,05; **p<0,01; *** p<0,001
Таблица 2. Фенетические дистанции (MMD±MSD) между популяциями малой лесной и домовой мыши. Верхняя треугольная матрица - данные по малой лесной мыши, а нижняя треугольная матрица по домовой мыши
|
Выборки |
г. Нальчик |
с. Псынадаха |
с. Бедык |
п. Эльбрус |
MMU |
|
|
малая лесная мышь |
домовая мышь |
|||||
|
1 |
0,021±0,007 |
0,056±0,007 |
0,099±0,007 |
0,059 |
0,020 |
|
|
2 |
0,016±0,007 |
0,040±0,006 |
0,050±0,006 |
0,037 |
0,027 |
|
|
3 |
0,004±0,006 |
0,023±0,006 |
0,046±0,006 |
0,047 |
0,028 |
|
|
4 |
0,041±0,007 |
0,041±0,006 |
0,057±0,006 |
0,065 |
0,046 |
|
Таблица 3. Частота встречаемости фенов неметрических признаков черепа в выборках домовой мыши Центрального Кавказа, %
|
Номер и код признака |
1 |
2 |
3 |
4 |
1-2 |
1-3 |
2-3 |
1-4 |
2-4 |
3-4 |
|
N=90 |
N=98 |
N=118 |
N=102 |
Хи - квадрат |
||||||
|
1. FFracan (-) |
2,2 |
5,1 |
6,78 |
0 |
||||||
|
2. FIO |
71,1 |
66,3 |
63,6 |
70,6 |
||||||
|
3. FTmacan |
4,4 |
10,2 |
9,32 |
17,7 |
** |
|||||
|
4. MeTm |
73,3 |
57,1 |
63,6 |
71,6 |
||||||
|
5. FeMs (-) |
4,4 |
3,1 |
8,5 |
0 |
||||||
|
6. FHg |
55,2 |
64,2 |
46,6 |
54,0 |
** |
|||||
|
7. FHgtr |
5,8 |
3,1 |
1,7 |
8,8 |
* |
|||||
|
8. FPT |
31,1 |
37,8 |
35,6 |
38,2 |
||||||
|
9. FLTIdu |
1,1 |
2,04 |
1,0 |
3,9 |
||||||
|
10. FPmvean |
52,2 |
68,1 |
56,8 |
78,4 |
* |
*** |
*** |
|||
|
11. FMtlgdu |
5,6 |
9,4 |
6,8 |
1,96 |
||||||
|
12. FPl |
21,4 |
24,5 |
20,3 |
27,5 |
||||||
|
13. FPCdu (M 2 ) |
30,1 |
22,9 |
28,0 |
23,5 |
||||||
|
14. FMbmst |
25,0 |
27,1 |
18,8 |
8,8 |
||||||
|
15. FBsla |
21,1 |
32,7 |
20,3 |
37,3 |
||||||
|
16. Fasac |
68,9 |
52,0 |
77,8 |
74,5 |
* |
*** |
*** |
|||
|
17. FMxzmla |
24,4 |
31,6 |
40,7 |
21,6 |
* |
** |
||||
|
18. FPmla |
0 |
1,0 |
0 |
2,9 |
||||||
|
19. FFsOc(mx) |
4,6 |
2,0 |
6 |
7,0 |
||||||
|
20. FCS |
3,4 |
0 |
1,7 |
1,0 |
* |
|||||
Примечание: 1 - г. Нальчик, 2 - с. Псынадаха, 3 - с. Бедык, 4 - п. Эльбрус; N-число изученных сторон черепа. Уровень значимости межгрупповых различий по отдельным признакам: * p<0,05; **p<0,01; *** p<0,001
первые два значимо чаще встречаются в псы-надахской, а последний в бедыкской выборке (табл. 3). По частоте встречаемости 4 признаков выявлены различия между выборками Псынадаха - Нальчик. Фенетические дистанции, полученные между всеми парами выборок домовой мыши ниже, чем у лесной, а в некоторых случаях даже статистически недостоверны. Недостоверной оказалась дистанция между бедык-ской и нальчикской выборками (0,004±0,006; d.f.=19; p>0,05) , статистически значимой, но небольшой между псынадахской и нальчикской (0,016±0,007; d.f.=19; p<0,05), а величина MMD между псынадахской и бедыкской выборками домовой мыши в 1,7 раз ниже, чем у малой лесной мыши (0,023±0,006; d.f.=19; p<0,01). Таким образом, уровень фенетического обособления предгорных антропогенных и контрольной выборок лесной мыши выше, чем у домовой.
Учитывая, что раннее проведенные на Центральном Кавказе исследования [10], показали, что высота над уровнем моря и связанные с ней условия среды (высокая ультрафиолетовая радиация, низкое атмосферное давление, разреженность воздуха, недостаток тепла, резкие суточные и сезонные перепады температур и влажности и т.д.) оказывают на организм малой лесной мыши более неблагоприятные воздействия, чем условия загрязнения отходами Тыр-ныаузского вольфрамо-молибденового комбината представляло интерес оценить влияние высоты местности на выборки исследуемых ви- дов. С этой целью в анализ включена выборка из среднегорий Центрального Кавказа – п. Эльбрус – 1800 м над ур. м.
Результаты сравнения выборок с учетом высоты местности демонстрируют следующую картину. При сравнении среднегорной выборки (п. Эльбрус) с предгорной (г. Нальчик) у малой лесной мыши различия выявлены по частоте встречаемости 7 признаков из 20, что составляет 35% от изученного числа признаков. Частота встречаемости отмеченных признаков в большинстве случаев выше в эльбрусской выборке, за исключением двух признаков – отверстие в основании крыловидного отростка (FPT) и одиночное отверстие на небной кости (FPl), отмечающихся с высокой частотой в нальчикской выборке. Фенетическая дистанция оказывается статистически высоко достоверной: MMD= 0,099±0,007 ( χ 2= 113,5; d.f.=19; p<0,001). Почти в 2 и более раза ниже фенетическая дистанция, полученная между среднегорной (п. Эльбрус) и предгорными (Псынадаха, Бедык) выборками из агроценозов: 0,050±0,006 ( χ 2=71,23; d.f.=19; p<0,001) и 0,046±0,006 ( χ 2=70,11; d.f.=19; p<0,001) соответственно. Аналогичное сравнение выборок домовой мыши, происходящих с разных высотных уровней (Эльбрус – Нальчик) выявило значимые различия по 5 признакам (25%). Также как и у лесной мыши, частота встречаемости большинства признаков (3 из 5) выше у зверьков среднегорья. Однако, у домовой мыши фенетическая дистанция, полученная между среднегорной и предгорной выборками уступает таковой малой лесной мыши в 2 и более раза: MMD=0,041±0,007 ( χ 2=58,9; d.f.=19;
p<0,001). У обоих видов в большей степени дифференцированной и наиболее уникальной оказывается эльбрусская выборка. Из полученных результатов можно предположить, что высота выступает в качестве существенного фактора, усиливающего уровень дифференциации между выборками, что наиболее ярко проявляется на выборках малой лесной мыши.
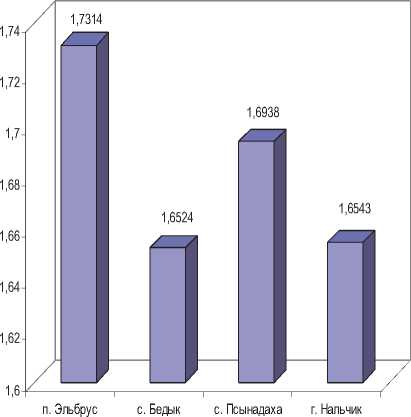
С целью оценки уровня внутрипопуляци-онного фенетического разнообразия антропогенных и контрольной выборок малой лесной и домовой мыши использовали показатель Л.А. Животовского (1980) – μ. Выявлено, что у малой лесной мыши фенетическое разнообразие достоверно выше в псынадахской выборке, тогда как бедыкская и нальчикская выборки характеризуются практически близкими значениями данного показателя и не отличаются. У домовой мыши величина фенетического разнообразия также выше в псынадахской выборке. Достоверные различия отмечены между бедык-ской и нальчикской, псынадахской и нальчикской выборками, между двумя антропогенными выборками по данному показателю различий не выявлено.
При включении в анализ среднегорной выборки (п. Эльбрус) отмечается, что у малой лесной мыши величина фенетического разнообразия достоверно выше в выборке с наибольшей высоты местности, чем во всех остальных. Интересно, что у домовой мыши среднегорная выборка напротив, отличается от всех остальных достоверно низкими значениями данного показателя.
Таким образом, выявлена видоспецифическая реакция на действие, как фактора вы-
Малая лесная мышь
Домовая мышь
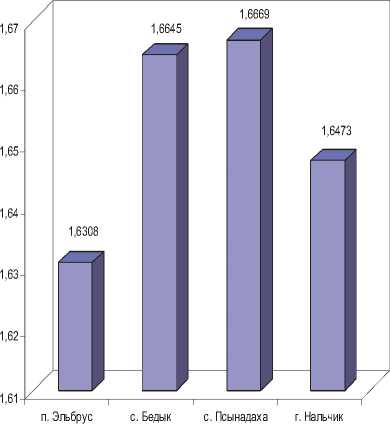
Рис. 2. Характеристика степени фенетического разнообразия выборок двух видов грызунов
соты, так и антропогенного загрязнения, что проявляется в несходной картине изменения частот фенов у исследуемых видов грызунов. У лесной мыши при сравнении всех выборок, включая и антропогенные, отмечены определенные закономерности в изменениях частот фенов. Так в направлении от равнин к горам увеличивается частота признаков – отсутствие окна на сосцевидной кости (FeMs (-)), дополнительное отверстие в области овального отверстия (Fasac) и множественные отверстия на затылочной кости в сочленовной ямке (FFsOc(mx)). Можно предположить, что данные признаки маркируют выборку с наибольшей высоты местности. У домовой мыши каких-либо закономерностей в изменениях частот фенов не выявлено.
В целом, параллельный анализ позволил выявить различия в частотах встречаемости признаков, выявленных у обоих видов. Так, все выборки малой лесной мыши характеризуются высокой частотой встречаемости особей, имеющих мыщелковые отверстия (55,6%, 59,6%, 54,7% и 64,6%), двойное отверстие на перегородке между овальным и круглым отверстием (18,3%, 28,7%, 27,8% и 42,7%), боковое предчелюстное отверстие (26,8, 33,3%, 26,9%, и 25,5%) тогда как в выборках домовой мыши данные признаки встречаются с очень низкой частотой (3,4%, 0%, 1,7%, 1%,), (1,1%, 2,0%, 1%, 3,9%), (0%, 1,0%, 0%, 2,9%) соответственно. У домовых мышей к числу таковых относятся признаки: наличие височного хода (73,3%, 57,1%, 63,6% и 71,6%), переднее вентральное предчелюстное отверстие (52,2%, 68,1%, 56,8% и 78,4%), боковое предчелюстное отверстие на скуловом отростке (24,4%, 31,6%, 40,7% и 21,6%) (табл. 1, 3).
ЗАКЛЮЧЕНИЕ
Результаты, проведенного фенетическо-го анализа двух видов показали, что различия между выборками из агроценозов и контрольными по величине MMD выше у малой лесной мыши по сравнению с домовой. Так, достоверные сдвиги частот при сравнении выборок малой лесной мыши из биотопов с разной антропогенной нагрузкой отмечаются по 3-7 признакам. Что касается домовой мыши, отмеченные выборки слабо отличаются по частотам фенов неметрических признаков, достоверные различия выявлены по одному, трем и четырем признакам. При этом у малой лесной мыши в большей степени дифференцированными оказываются бедыкская и нальчикская (табл. 2), тогда как у домовой псынадахская и бедыкская выборки, т.е. выборки из антропогенных биотопов. Фенети-ческая дистанция между бедыкской и наль- чикской выборками домовой мыши напротив, оказывается статистически недостоверной. В целом, полученные результаты указывают на разнонаправленное действие антропогенного фактора и высоты местности на изучаемые виды грызунов, что вероятно связано с особенностями исследуемых видов. Так, домовая мышь, обитая в постройках человека в меньшей степени подвержена влиянию средовых факторов, чем малая лесная мышь. Возможно, в качестве второй причины может быть и то, что популяции домовой мыши периодически пополняются за счет завоза зверьков из других районов, следствием чего может быть нивелировка эпигенетических различий.
Список литературы Сравнительный фенетический анализ популяций двух видов грызунов (Apodemus, Mus) в условиях Центрального Кавказа
- Васильев А.Г., Васильева И.А. Феногенетический мониторинг импактных популяций растений и животных в условиях антропогенного пресса//Научные ведомости Белгородского государственного университета. Естественные науки. 2009. Вып. 8. № 3(58). С. 5 -12.
- Васильев А.Г., Васильева И.А., Большаков В.Н. Сравнение мигрирующих и оседлых особей рыжей полевки по комплексу неметрических признаков//Экология. 1996. Т. 27. № 5. С. 371 -377.
- Васильев А.Г. Эпигенетические основы фенетики: на пути к популяционной мерономии. Екатеринбург: Академкнига, 2005. 640 с.
- Песков В.Н., Емельянов И.Г. Фенетика и феногеография водяной полевки (Arvicola terrestris L.)//Вест. зоологии. 2000. № 3. С. 39 -44.
- Феногенетический анализ популяций малой лесной мыши (Apodemus uralensis Pall.) в зоне влияния Восточно-Уральского радиоактивного следа // И.А. Васильева, А.Г. Васильев, Н.М. Любашевский, М.В. Чибиряк, Е.Ю. Захарова, О.В. Тарасов // Экология. 2003. № 6. С. 325 - 332.
- Васильев А.Г., Васильева И.А. Гомологическая изменчивость морфологических структур эпигенетическая дивергенция таксонов: Основы порпуляционной мерономии. М.: Тов-во науч. изд. КМК, 2009. 511с
- Hartman S.E. Geographic variation analysis of Dipodomys ordii using nonmetric cranial trains//J. Mammal. 1980. V.61. № 3. P. 436 -448.
- Васильев А.Г., Васильева И.А., Большаков В.Н. Эволюционно-экологический анализ устойчивости популяционной структуры вида (хроно-географический подход)//Екатеринбург: Изд-во «Екатеринбург». 2000 а. 132 с.
- Животовский Л.А. Показатели популяционной изменчивости по полиморфным признакам//Фенетика популяций. М.: Наука, 1982. С. 38 -44.
- Амшокова А.Х. Фенетический анализ краниологических признаков малойц лесной мыши (Sylvaemus uralensis Pall.) в условиях загрязнения тяжелыми металлами в среднегорьях Центрального Кавказа//Известия Самарского научного центра РАН. 2009. Т. 11. № 1. С.110 -117.
