Сравнительный лейкоцитарный профиль и размерные характеристики клеток крови сухопутных черепах из Самарского зоопарка

Автор: Романова Е.Б., Столярова И.А., Бакиев А.Г., Горелов Р.А.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Зоология - биологические науки
Статья в выпуске: 5 т.25, 2023 года.
Бесплатный доступ
Проведен сравнительный анализ лейкограмм и морфометрических особенностей клеток крови трех видов сухопутных черепах из Самарского зоопарка: Testudo horsfieldii Gray, 1844; Geochelone elegans (Schoepff, 1795); Chelonoidis carbonarius (Spix, 1824). Выявлены межвидовые статистически значимые различия по суммарному содержанию агранулоцитов и гранулоцитов. Лейкоцитарных профиль свидетельствовал о выраженных реакциях врожденного иммунного ответа сухопутных черепах и более активном клеточно-опосредованном адаптивном ответе C. carbonarius. По совокупности интегральных показателей лейкоцитарного профиля наиболее стабильный и сбалансированный иммунный ответ отмечен у T. horsfieldii. Выявлена вариабельность кариоцитометрических параметров клеток крови черепах. Доля вариабельности площади ядер клеток, зависящая от площади самой клетки, была наибольшей для разных клеток у разных видов: T. horsfieldii (базофилы), G. elegans (тромбоциты), C. carbonarius (эозинофилы). Площадь клеток крови (за исключением эозинофилов) и ядерно-цитоплазматические отношения однотипных клеток разных видов черепах варьировали. Установлена более высокая метаболическая активность клеток крови T. horsfieldii по сравнению с G. elegans и C. carbonarius.
Экологическая иммунология, клетки крови, лейкоцитарная формула, кариоцитометрия
Короткий адрес: https://sciup.org/148327988
IDR: 148327988 | УДК: 598.132.4 | DOI: 10.37313/1990-5378-2023-25-5-58-70
Comparative leukocyte blood profile and size of blood cells of tortoises from the Samara Zoo
A comparative analysis of leukograms and morphometric features of blood cells of three species of tortoises from the Samara Zoo was carried out: Testudo horsfieldii Gray, 1844; Geochelone elegans (Schoepff, 1795); Chelonoidis carbonarius (Spix, 1824). Interspecies statistically significant differences in the total content of agranulocytes and granulocytes were revealed. The leukocyte profile indicated a pronounced innate immune response in tortoises and a more active cell-mediated adaptive response in C. carbonarius. According to the totality of integral indicators of the leukocyte profile, the most stable and balanced immune response was noted in T. horsfieldii. The variability of karyocytometric parameters of turtle blood cells was revealed. The share of variability in the area of cell nuclei, depending on the area of the cell itself, was the highest for different cells in different species: T. horsfieldii (basophils), G. elegans (platelets), C. carbonarius (eosinophils). The area of blood cells (with the exception of eosinophils) and the nuclear-cytoplasmic ratios of cells of the same type in different turtle species varied. A higher metabolic activity of blood cells of T. horsfieldii was established in comparison with G. elegans and C. carbonarius.
Текст научной статьи Сравнительный лейкоцитарный профиль и размерные характеристики клеток крови сухопутных черепах из Самарского зоопарка
Иммунная защита играет важную роль для позвоночных животных в компромиссах их жизненного цикла [16]. Обнаружение таких компромиссов осложняется сложностью иммунной системы. Измерение иммунных показателей имеет большой потенциал в исследованиях экологической иммунологии для дальнейшего понимания экологических и эволюционных сил, вызывающих ответы иммунного статуса позвоночных.
Testudinidae включает, согласно распространенной в настоящее время систематике [23], более сорока современных видов, относящихся к 17 родам. Стратегии адаптации сухопутных черепах на уровне системы крови к естественным условиям обитания значительно варьируют в зависимости от систематической принадлежности этих животных, возраста, пола, времени года и опосредованы экологическими факторами [1, 2, 5, 18]. В связи с этим, исследования специфики адаптивных реакций черепах следует проводить не только в естественных, но и в унифицированных оптимальных условиях террариумного содержания. Необходимо учитывать, что таксоны позвоночных животных, подверженные из-за экологических ограничений более широкому разнообразию патогенов и высокому риску заболеваний, развивают более сильную иммунную защиту и отличаются значительными различиями в морфологии циркулирующих клеток и физиологических нормах гематологических показателей [16].
Наше исследование направлено на сравнительную оценку показателей лейкоцитарного профиля и размерных особенностей клеток крови представителей трех родов семейства
Testudinidae – Testudo Linnaeus, 1758, Geochelone Fitzinger, 1835 и Chelonoidis Fitzinger, 1835. Полученные результаты можно использовать как данные к установлению базового уровня адаптивных реакций организма черепах: не только для фундаментальных исследований, но также для прикладных задач (для лечения сухопутных черепах в случае выявления патологий).
МАТЕРИАЛ И МЕТОДЫ
Объектами исследования явились содержащиеся в Самарском зоопарке половозрелые особи трех видов сухопутных черепах: среднеазиатская черепаха Testudo horsfi eldii Gray, 1844 – два самца и две самки; звёздчатая черепаха Geochelone elegans (Schoepff, 1795) – один самец; красноногая (= угольная) черепаха Chelonoidis carbonarius (Spix, 1824) – один самец. Террариумные условия содержания этих особей были следующие. В качестве подстилочного материала для среднеазиатской черепахи использована супесь, для звёздчатой и красноногой черепах – кокосовая щепа и сфагнум. Террариумы, где содержались последние два вида, имели водоемы. Террариумы, где содержались все три вида, оборудованы лампами с ультрафиолетовым спектром, искусственный световой день составлял от 10 (зимой) до 12 (летом) часов. Рацион всех названных черепах включал: зелень злаковых, подорожниковых и астровых растений, фрукты (яблоки, апельсины, бананы), овощи (капуста, морковь, листовой салат), вареные куриные яйца; рацион красноногой черепахи дополнялся мышами и говяжьим мясом.
T. horsfi eldii и G. elegans сейчас фигурируют в Красном списке МСОП с категорией VU (уязвимый вид), категория для Chelonoidis carbonarius пока не установлена [19].
Все работы проводились в соответствии с «Международными руководящими принципами для биомедицинских исследований на животных» (International Guiding…, 2012). Кровь у черепах брали путем прокола шейной вены. Мазки крови (по два от каждой особи) готовили с помощью гематологического штапеля, высушивали на воздухе, затем фиксировали в спирто-эфирной смеси (1:1). Зафиксированные мазки хранили до окрашивания при комнатной температуре. Мазки окрашивали в течение 20 минут красителем Гимза азур-эозин по Романовскому (10–12 кратный раствор, фирма «Био-лот»), приготовленном на фосфатном буфере (рН 6,8–7,2). Готовые мазки просматривали с иммерсией, при увеличении ×1500, с дифференцированным подсчетом лейкоцитарной формулы крови [3, 5]. С учетом морфологических особенностей определяли пять типов лейкоцитарных клеток (в %): гранулоциты (гетерофилы, базофилы, эозинофилы) и агранулоциты (моноциты, лимфоциты).
На основании лейкоцитарной формулы крови рассчитали интегральные лейкоцитарные индексы в относительных единицах (обозначения: Л – лимфоциты; Э – эозинофилы; Г – гетерофилы; Б – базофилы).
-
1. Индекс сдвига лейкоцитов, ИСЛ = Σ гранулоцитов / Σ агранулоцитов.
-
2. Индекс соотношения лимфоцитов и эозинофилов, ИСЛЭ = Л / Э.
-
3. Индекс соотношения гетерофилов и эозинофилов, ИСГЭ = Г / Э.
-
4. Лимфоцитарно-гранулоцитарный индекс, ИЛГ = Л × 10 / Э + Г + Б.
-
5. Индекс соотношения гетерофилов и лимфоцитов, ИСГЛ = Г / Л.
Цифровые изображения клеток выполняли камерой Vision САМ для тринокулярного микроскопа Meiji Techno (Japan, серия МТ 4000) с иммерсией, увеличение ×1500.
При цитометрическом и кариометрическом анализах изучены 1475 клеток крови ( T. horsfi eldii – 700; G. elegans – 488; C. carbonarius – 287 клеток). Определение размеров клеток крови и их ядер проводили с помощью окуляр-микрометра cross-line (0.1 mm). Площадь поверхности клеток S к и ядер S я (в мкм2) рассчитывали по формуле эллипса: S = π × a × b , где π = 3,14; a – длина большой полуоси эллипса, мкм; b – длина меньшей полуоси эллипса, мкм. Индекс удлинения клеток (Е) и их ядер (е) рассчитывали, как частное от деления длинного диаметра на короткий. Ядерно-цитоплазматическое отношение (ЯЦО) определяли как отношение площади ядра к площади клетки.
Данные сравнивали методами непараметрической статистики с расчетом критериев: Кра-скела – Уоллиса ( H ) (при множественном сравнении независимых групп по одному признаку), Данна ( D ) (множественный критерий при попарном сравнении групп), Манна – Уитни ( u ) при сравнении двух групп в пакете прикладных программ «STATISTICA v.10» и в среде R (R Core Team, 2015). Для упорядочивания и классификации показателей применялся метод главных компонент ( Principal component analysis, PCA). Методом многофакторного дисперсионного анализа по совокупности показателей определяли различие между видами. Для изучения зависимости метрических показателей клеток крови использовали регрессионный анализ. За величину статистической значимости принимали α = 0.05.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Клетки периферической крови сухопутных черепах, как и всех рептилий, представлены эритроцитами, лейкоцитами и тромбоцитами.
Зрелые эритроциты морфологически сходны у разных видов черепах [20] и представляют собой эллипсоидные клетки с округлыми полюсами и однородной серо-голубой цитоплазмой (рис. 1а, 2а, 3а).
Ядро эритроцита удлиненное с округлыми или неправильными полюсами. Длинный диаметр ядра параллелен большому диаметру клетки. Плотный хроматин окрашивается в темнофиолетовый цвет.
У черепах присутствуют два типа тромбоцитов: овальные с хорошо видимой мембраной и прозрачной цитоплазмой, и почти прямоугольные с мелкими выступами и слабо базофильной цитоплазмой [14]. Ядра тромбоцитов округлоовальные, темные (рис. 1б, 2б, 3б).
Классификация лейкоцитов рептилий вызывает много проблем, поскольку эти клетки демонстрируют морфологическую изменчивость, тем не менее количественное определение профиля лейкоцитов является надежным показателем иммунной функции и большинство исследователей дифференцирует пять основных типов: гранулоциты (гетерофилы, эозинофилы, базофилы) и агранулоциты (моноциты и лимфоциты) [7,15, 21, 22, 24]. Гетерофилы трех видов сухопутных черепах - это крупные клетки округлой или овальной формы с веретеновидными гранулами в цитоплазме и эксцентрично расположенным ядром (рис. 1в, 2в, 3в).
Менее многочисленные эозинофилы представляли собой округлые клетки. Их гранулы были более темными и круглыми. Ядро располагалось эксцентрично, однородного цвета, с комковатым хроматином (рис. 1г, 2г, 3г).
Цитоплазма базофилов была заполнена округлыми крупными гранулами, выступающими по периферии клетки. Цвет варьировал от темно-фиолетового до темно-синего. Ядра со сгустками хроматина практически не выделялись (рис. 1д, 2д, 3д).
Моноциты представляли собой клетки с крупными бобовидными ядрами с бледным и тонким хроматином, расположенными в сине-серой цитоплазме с вакуолями или без них (рис. 1е, 2е, 3е). Известно, что у рептилий имеется уникальный тип лейкоцитов - азурофилы, округлые клетки моноцитарного роста с крупным слегка изрезанным или полушаровидным ядром с грубо глыбчатым темно-фиолетовым хроматином и азурофильными гранулами [4]. Поскольку азурофилы принадлежат к молодым клеткам моноцитарного ряда и крайне редко встречаются у черепах [14, 17], в отдельную категорию мы их не выделяли. Лимфоциты имели компактное темное ядро и тонкую кайму цитоплазмы синего или бледно-фиолетового цвета (рис. 1ж, 2ж, 3ж).
Половые различия по лейкоцитарной системе крови среднеазиатской черепахи не выявлены, поэтому данные были объединены в общую группу, проведен сравнительный анализ гематологических препаратов ( n = 12) трех видов сухопутных черепах (табл. 1). Установлены

Рис. 1. Клетки крови Testudo horsfieldii : а- эритроцит; б - тромбоцит; в - гетерофил; г - эозинофил; д - базофил; е - моноцит; ж - лимфоцит
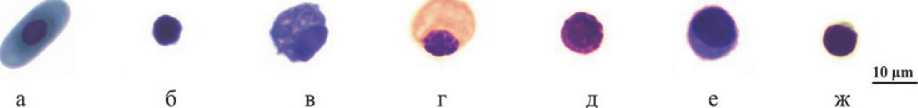
Рис. 2. Клетки крови Geochelone elegans : а- эритроцит; б - тромбоцит; в - гетерофил; г - эозинофил; д - базофил; е - моноцит; ж - лимфоцит
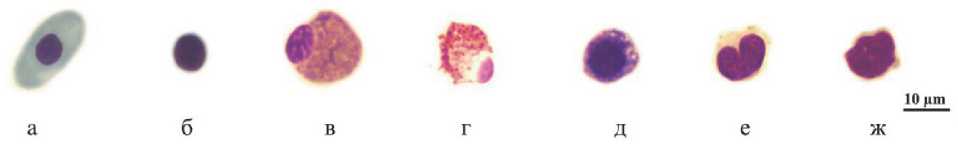
Рис. 3. Клетки крови Chelonoidis carbonarius : а - эритроцит; б - тромбоцит; в - гетерофил; г - эозинофил; д - базофил; е - моноцит; ж - лимфоцит
межвидовые статистически значимые различия по суммарному содержанию агранулоцитов и гранулоцитов. В порядке возрастания по содержанию гранулоцитов виды черепах расположились в ряду: красноногая ^ среднеазиатская ^ звёздчатая. В порядке возрастания по содержанию агранулоцитов - в ряду: звёздчатая ^ среднеазиатская ^ красноногая.
Лимфоциты и гетерофилы являются двумя наиболее распространенными типами лейкоцитов рептилий, которые вместе составляют до 95% всех лейкоцитов [8], но имеют различные иммунные функции. Доминирующими клетками в лейкограммах сухопутных черепах среди агранулоцитов были лимфоциты (40-55%), среди гранулоцитов - гетерофилы (27-33%). Лимфоциты играют главную роль в клеточно-опосредованном адаптивном иммунитете (Т-клетки) и гуморальном адаптивном имму- нитете (В-клетки), поскольку они продуцируют иммуноглобулины, которые специфически распознают антигены. За счет повышенного содержания лимфоцитов и моноцитов доля клеток агранулоцитарного ряда в крови красноногой черепахи была выше по сравнению с другими видами (H = 14,05, р = 0,001). Гетерофилы представляют собой фагоцитирующие клетки врожденной иммунной системы, формирующие первую линию иммунной защиты от бактериальных патогенов при воспалительных поражениях [12]. Содержание гетерофилов (Н = 3,45, р = 0,18) в лейкограммах сухопутных черепах трех видов не различалась, так же, как и базофилов (Н = 1,169, р = 0,558). В иммунном ответе, как известно, существуют сложные взаимосвязи [9, 10], поскольку врожденная и адаптивная системы не являются полностью независимыми друг от друга, поэтому гетерофилы и лимфоциты
Таблица 1. Лейкоцитарный состав периферической крови сухопутных черепах из Самарского зоопарка
|
Показатель лейкограммы |
Статистические показатели |
||||||
|
Me |
IQR |
Me |
IQR |
Me |
IQR |
H, P, D |
|
|
Виды черепах |
|||||||
|
1. Testudo horsfieldii |
2. Geochelone elegans |
3. Chelonoidis carbonarius |
|||||
|
Гетерофилы, % |
29,00 |
10,75 |
33,00 |
3,25 |
27,50 |
3,00 |
Н = 3,45, р = 0,18 |
|
Эозинофилы, % |
6,00 |
3,00 |
12,00 |
3,75 |
1,50 |
1,50 |
Н = 10,170, р = 0,006 D 1-2 = 1,360, p = 0,59; D 1-3 = 2,554, p = 0,015; D 2-3 = 3,095, p = 0,005 |
|
Базофилы, % |
8,50 |
4,00 |
12,50 |
5,25 |
11,00 |
3,00 |
Н = 1,169, р = 0,558 |
|
Моноциты, % |
5,50 |
6,25 |
1,50 |
1,00 |
8,00 |
1,75 |
Н = 9,287, р = 0,010 D 1-2 = 3,027, p = 0,001; D 1-3 = 0,262, p = 0,397; D 2-3 = 2,190, p = 0,014 |
|
Лимфоциты, % |
47,00 |
5,50 |
42,00 |
6,25 |
55,50 |
4,75 |
Н = 8,947, р = 0,011 D 1-2 = 1,720, p = 0,078; D 1-3 = 2,053, p = 0,055; D 2-3 = 2,983, p = 0,008 |
|
Гранулоциты, % |
45,00 |
4,25 |
56,50 |
5,75 |
38,00 |
3,50 |
Н = 14,221, р = 0,001 D 1-2 = 2,570, p = 0,014; D 1-3 = 2,190, p = 0,026; D 2-3 = 3,763, p = 0,005 |
|
Агранулоциты, О/ % |
55,00 |
4,25 |
43,50 |
5,75 |
61,00 |
3,50 |
Н = 14,056, р = 0,001 D 1-2 = 2,579, p = 0,014; D 1-3 = 2,150, p = 0,029; D 2-3 = 3,739, p = 0,001 |
Примечание: Me - медиана, IQR - интерквартильный размах, Н - Критерий Краскела - Уоллиса; D - критерий Данна, р - уровень значимости. Жирным шрифтом выделены статистически значимые различия классифицируют в качестве основных составляющих компонентов врожденной и адаптивной иммунной защиты, соответственно [16]. Доля эозинофилов в гранулоцитарной составляющей крови красноногой черепахи была минимальной по сравнению с другими видами.
Анализ полученных лейкоцитарных профилей свидетельствовал о сопоставимом уровне врожденного иммунного ответа трех видов сухопутных черепах и более выраженном клеточно-опосредованном адаптивном иммунитете у красноногой черепахи .
Оценка интегральных лейкоцитарных индексов в крови, как маркеров иммунного статуса организма черепах в условиях неволи (средового стресса) представляет наибольший интерес. Известно, что соотношение гетерофилов к лимфоцитам изменяются в ответ на внешние стрессоры, включая неблагоприятные климатические или погодные явления, загрязнение, усиленное размножение, антропогенный пресс или заражение паразитами и в экофизиологических исследованиях соотношение этих клеток используется как надежная мера физиологического стресса, указывая на взаимодействие между иммунитетом, физиологией и экологией вида; соотношение гетерофилов к лимфоцитам (ИСГЛ) не измеряет иммунный ответ как таковой [8], однако может отражать готовность организма справляться с инфекцией предпочтительнее за счет неспецифической резистентности организма (гетерофилов), чем с помощью специфической иммунной составляющей (через лимфоциты) [13]. Сравнительный анализ индекса ИСГЛ сухопутных черепах выявил повышенное значение этого показателя у самца звёздчатой черепахи, что иллюстрировало более выраженную активацию неспецифической защитной системы крови за счет суммарного вклада гранулоцитарных клеток по сравнению с красноногой черепахой, в крови которой прослеживалась тенденция повышения специфических реакций. Дополнительную информацию при оценки адаптивных возможностей организма можно получить при расчете интегральных лейкоцитарных индексов: ИСЛ - отражающего адекватность и своевременность иммунного ответа и ИЛГ - отражающего его сбалансированность. Низкий показатель ИЛГ звёздчатой черепахи свидетельствовал о недостаточном ресурсе адаптивных механизмов организма в условиях неволи по сравнению с другими видами.
Анализ лейкоцитарных индексов реактивности красноногой черепахи ( C. carbonarius ) выявил разнонаправленность изменений, а именно, повышение лимфоцитарно-гранулоцитарного индекса ИЛГ, снижение ИСЛ и ИСГЛ, что иллюстрировало активную и сбалансированную реакцию организма за счет вклада лимфоцитов.
По уменьшению значения индекса ИСЛ сухопутные черепахи расположились в ряду: звёздчатая ^ среднеазиатская ^ красноногая. По уменьшению значения индекса ИЛГ: красноногая ^ среднеазиатская ^ звёздчатая.
С целью изучения взаимосвязей, а также выявлении факторов, которые бы упростили процесс классификации изучаемых выборок по интегральным лейкоцитарным показателям, мы воспользовались методом главных компонент. Основным свойством этого метода является отсутствие корреляции, в отличие от исходных переменных, за счет этого и достигается концентрация информации. Вначале определили факторные оси (главные компоненты) в пространстве меньшей размерности, на которые спроецировали пространство переменных анализа (рис. 4). Первая главная компонента, соответствующая максимальному собственному значению 229,78 описывала приблизительно 89,23% общей дисперсии. Вторая компонента с собственным значением 26,20 описывала 10,17% оставшейся дисперсии. Третья и последующие компоненты были менее информативны.
Из построенных таблиц факторных координат переменных в пространстве главных компонент следует, что первая компонента (факторная ось) обладала сильной положительной корреляцией с переменными: ИСЛЭ (0,98), ИСГЭ (0,96), ИЛГ (0,92), суммой агранулоцитов (0,83) и сильной отрицательной корреляцией с переменными сумма гранулоцитов (-0,84) и ИСЛ (-0,78). Вторая главная компонента имела умеренную положительную корреляцию с суммой агранулоцитов (0,55) и ИЛГ (0,31), и умеренную отрицательную корреляцию с суммой гранулоцитов (-0,52) и ИСГЛ (-0,38).
График рассеяния выборок в пространстве двух первых компонент визуализировал отличия лейкоцитарного профиля сухопутных черепах. По совокупности интегральных показателей лейкоцитарного профиля среднеазиатская черепаха отличалась наиболее стабильным и сбалансированным иммунным ответом по отношению к другим видам сухопутных черепах Самарского зоопарка и группировалась во втором квадранте факторного пространства. Удаленно, в четвертом квадранте в области локализации переменных ИСЛЭ и ИСГЭ располагалась красноногая черепаха. Звёздчатая черепаха, имеющая в крови повышенную доля гранулоцитов всех форм, занимала третий квадрант (рис. 5).
Ранговым анализом вариаций по Краскела - Уоллису (Н) размерных характеристик клеток крови сухопутных черепах нулевая гипотеза об отсутствии различий была отклонена и принята альтернативная гипотеза о различии клеточных параметров. Для среднеазиатской черепахи са-
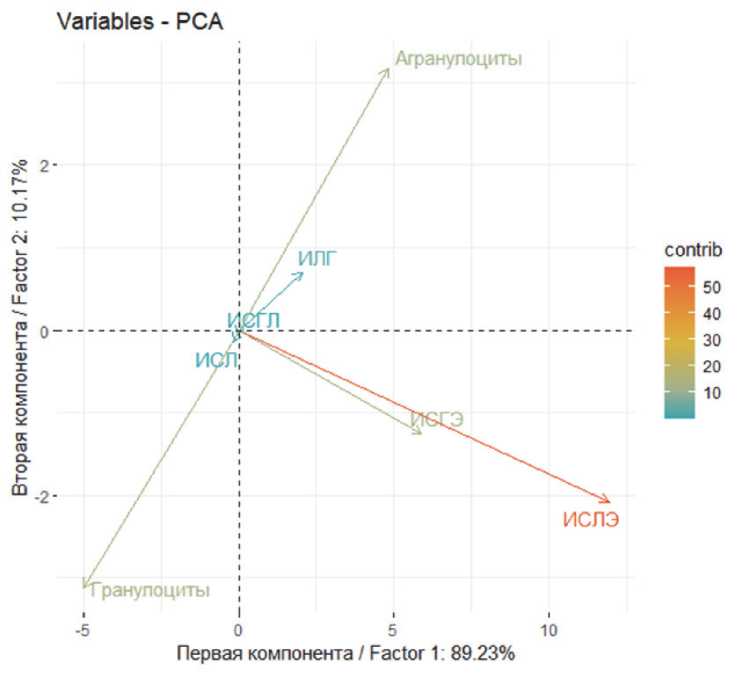
Рис. 4. График факторных координат, построенный на основе интегральных лейкоцитарных показателей крови черепах
Вторая компонента / Factor 2: 10.17%
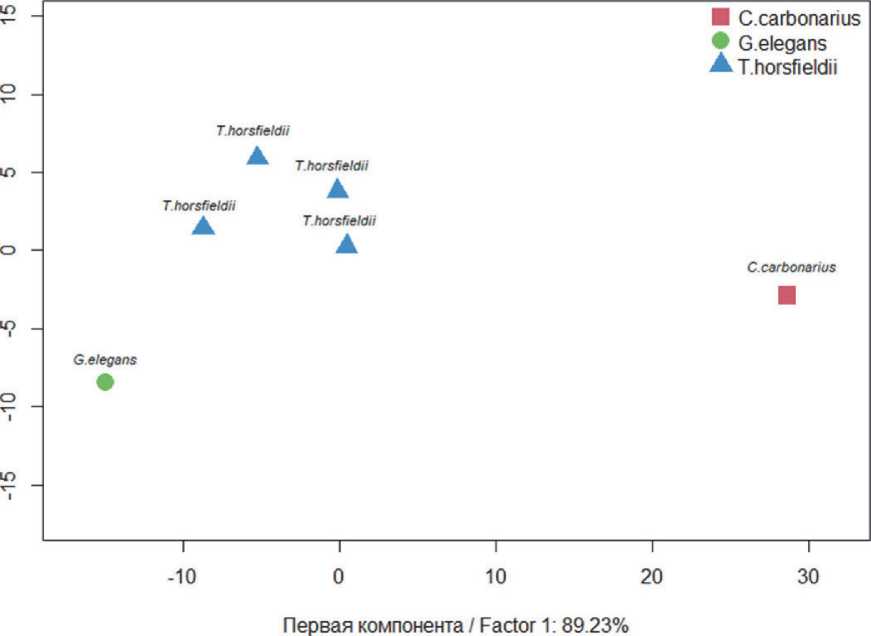
Рис. 5. График рассеяния особей сухопутных черепах Самарского зоопарка в пространстве главных компонент мыми крупными клетками оказались гетерофилы, которые по площади превосходили эритроциты (z = 5,27, p < 0,001), тромбоциты (z = 17,39, p < 0,001), эозинофилы (z = 6,68, p < 0,001), базофилы (z = 7,96, p < 0,001), моноциты (z = 3,15, p = 0,033), и лимфоциты (z = 14,07, p < 0,001). Ядра лейкоцитарных клеток были крупнее ядер эритроцитов и тромбоцитов. При этом у лейкоцитов наименьшую площадь имели ядра эозинофилов. Наиболее крупных ядра выявлены у моноцитов (65,47 мкм2), площадь которых в 3,37 раза превосходила площадь ядра эритроцитов (z = 16,41, p < 0,001) и в 2,64 раза – площадь ядра тромбоцитов (z = 13,13, p < 0,001). Площади ядер гетерофилов, базофилов и лимфоцитов среднеазиатской черепахи были сопоставимы и варьировали от 38 до 45 мкм2.
У звёздчатой черепахи самые мелкие клетки: лимфоциты и тромбоциты по линейным параметрам не различались (z = 2,34, p = 0,39). Эритроциты по индексу удлинения (Е) значимо отличались от всех клеток крови. Наиболее округлыми были базофилы, индекс удлинения которых приближался к единице. Наиболее крупные клетки (гетерофилы, эозинофилы и моноциты) были схожи по размерам.
Эритроциты красноногой черепахи не отличались по площади от эозинофилов и моноцитов. Наиболее крупными среди лейкоцитов оказались гетерофилы и моноциты. Эозинофилы и базофилы по площади были сопоставимы ( z = 0,46, p = 1,00).
Анализ зависимости площади ядер от размера клетки показывал, что для среднеазиатской черепахи только дисперсия площади ядер базофилов объяснялась дисперсией площади самой клетки на 98%. В случае других клеток эта зависимость выражена гораздо слабее и составляла от 0,8 до 21%. Напротив, для звёздчатой черепахи дисперсия площади ядра базофила объяснялась дисперсией площади самой клетки менее чем на 1%. При этом доля объяснимой дисперсии этих показателей, то есть доля вариабельности одного признака, зависящая от другого, для тромбоцита и моноцита составляла 88% и 66%, соответственно. Вариабельность площади ядра, зависящая от площади самой клетки, у красноногой черепахи составила 72% для эозинофилов и 48% для лимфоцитов (табл. 2).
Таблица 2. Анализ взаимозависимости площади ядер (y) и площади клеток (x) крови сухопутных черепах Самарского зоопарка
|
Клетка |
Уравнение регрессии |
r |
R2 |
p |
|
Testudo horsfieldii |
||||
|
Эритроцит |
у = 9,393 + 0,336 х |
0,40 |
0,15 |
0,00004 |
|
Тромбоцит |
у = 3,188 + 0,4271 х |
0,35 |
0,12 |
0,00021 |
|
Гетерофил |
у = 4,018 + 0,306 х |
0,39 |
0,15 |
0,00005 |
|
Эозинофил |
у = 2,208 + 0,364 х |
0,44 |
0,19 |
< 0,001 |
|
Базофил |
у = 0,022 + 0,994 х |
0,99 |
0,98 |
< 0,001 |
|
Моноцит |
у = 5,349 + 0,24 х |
0,29 |
0,08 |
0,0033 |
|
Лимфоцит |
у = 3,288 + 0,400 х |
0,45 |
0,21 |
< 0,001 |
|
Geochelone elegans |
||||
|
Эритроцит |
у = 11,444+0,061 х |
0,25 |
0,06 |
0,01 |
|
Тромбоцит |
у = - 0,808+0,811 х |
0,94 |
0,88 |
< 0,001 |
|
Гетерофил |
у = 9,439+0,108 х |
0,33 |
0,11 |
0,0007 |
|
Эозинофил |
у = 15,14+0,147 х |
0,43 |
0,18 |
0,0017 |
|
Базофил |
у = 18,47+0,207 х |
0,31 |
0,09 |
0,014 |
|
Моноцит |
у = 25,03+0,291 х |
0,81 |
0,66 |
< 0,001 |
|
Лимфоцит |
у = 25,85+0,219 х |
0,50 |
0,25 |
< 0,001 |
|
Chelonidis carbonarius |
||||
|
Эритроцит |
у = 1,64+0,146 х |
0,44 |
0,19 |
< 0,001 |
|
Тромбоцит |
у = 11,67+0,418 х |
0,53 |
0,28 |
0,015 |
|
Гетерофил |
у = 25,50+0,005 х |
0,02 |
0,0006 |
0,86 |
|
Эозинофил |
у = 25,99+0,088 х |
0,85 |
0,72 |
0,01 |
|
Базофил |
у = 24,82+0,226 х |
0,55 |
0,31 |
0,00002 |
|
Моноцит |
у = 44,47+0,186 х |
0,40 |
0,16 |
0,040 |
|
Лимфоцит |
у = 17,94+0,418 х |
0,69 |
0,48 |
< 0,001 |
По информативному индексу удлинения клетки различались эритроциты (Н = 55,60, р < 0,001) и тромбоциты (Н = 29,96, р < 0,001) всех видов черепах. Метрические параметры лейкоцитарных клеток были сходными и только индекс удлинения эозинофилов звёздчатой черепахи превышал аналогичный показатель среднеазиатской черепахи. По площади клетки крови разных видов варьировали, за исключением эозинофилов. Наиболее крупные эритроциты, тромбоциты, гетерофилы, базофилы и лимфоциты обнаружены у красноногой черепахи, площадь этих клеток значимо превышала аналогичные параметры двух других видов. Моноциты среднеазиатской и красноногой черепахи превышали площадь моноцитов звёздчатой черепахи (табл. 3).
Минимальные значения индекса удлинения ядра характерны для эритроцитов звёздчатой черепахи (1,29 ± 0,02). По индексу удлинения ядра гетерофилы звёздчатой черепахи (1,42 ± 0,05) занимали промежуточное положение между более округлыми ядрами среднеазиатской черепахи (1,23 ± 0,03) и более вытянутыми (1,64 ± 0,08) красноногой черепахи.
Ядра эритроцитов и базофилов красноногой черепахи превышали площадь ядер этих клеток среднеазиатской и звёздчатой черепах. Площади ядер моноцитов (Н = 4,49, р = 0,10) и эозинофилов (Н = 1,98, р = 0,36) трех видов черепах не различались.
Ядерно-цитоплазматическое отношение – объективный показатель оценки прижизненного состояния клетки и важная морфологическая характеристика, позволяющая получить определенное представление об уровне метаболизма и компенсаторных реакциях клеток эритроцитарного и лейкоцитарного рядов. Более высокие показатели ядерно-цитоплазматических соотношений свидетельствуют, что ядро в цитоплазме занимает больший объем. Сравнивая между собой морфометрические параметры, можно прийти к заключению, что в некрупных клетках крови (тромбоцитах и лимфоцитах) черепах ядерно-цитоплазматические соотношения были наибольшими. В крупных эритроцитах и гетерофилах ядерно-цитоплазматические отношения оказались наименьшими (табл. 4).
Следует обратить внимание на вариабельность ядерно-цитоплазматического отношения однотипных клеток разных видов черепах, что иллюстрировало разную степень функционирования и синтетической активности клеток. При этом, чем больше показатель, тем активнее идут метаболические синтетические процессы клетках конкретного вида. Методом многомерного дисперсионного анализа (λWilks = 0,651, р < 0,001) выявлены значимые различия по совокупности ядерно-цитоплазматических отношений всех изученных клеток крови трех видов черепах. В качестве примера на рис. 6 показано возрастание ЯЦО в ряду гетерофилов G. elegans → C. carbonarius → T. horsfi eldii, иллюстрирующее более высокую метаболическую активность клеток крови среднеазиатской черепахи выше по сравнению со звёздчатой и красноногой.
ЗАКЛЮЧЕНИЕ
Функциональные возможности организма сухопутных черепах и его реальные способности адаптироваться к разнообразным факторам среды (в том числе, к условиям неволи) интегрально отражало состояние им-муногематологической системы и уровень ее реактивности. Несмотря на небольшой размер выборки, удалось обнаружить вариации лейкоцитарного состава и морфометрических показателей крови, позволяющие предположить влияние условий содержания на состояние черепах, которые могут быть полезными при выявлении патологий и соответствующего лечения рептилий.
Показано, что среднеазиатская черепаха по совокупности интегральных показателей лейкоцитарного профиля характеризовалась более стабильным и сбалансированным иммунным ответом по сравнению с другими изученными видами сухопутных черепах Самарского зоопарка. Изучение кариоцито-метрических особенностей трех видов черепах позволило оценить и анализировать структурные изменения клеток крови, представляющие собой весьма сложные многокомпонентные системы, в которых устойчивость сочетается с большими возможностями адаптации к изменениям внутренней и внешней среды с множеством взаимосвязанных процессов. Выявлены значимые различия по большинству морфометрических характеристик клеток крови трех видов черепах. По площади клетки крови разных видов варьировали, за исключением эозинофилов. Наиболее крупные эритроциты, тромбоциты, гетерофилы, базофилы и лимфоциты обнаружены у красноногой черепахи, площадь этих клеток значимо превышала аналогичные параметры клеток двух других видов. Методом многомерного дисперсионного анализа выявлены значимые различия по совокупности ядерно-цитоплазматических отношений клеток крови трех видов черепах. Клетки крови среднеазиатской черепахи характеризовались более высокой метаболической активностью.
Настоящее исследование дополняет сведения о динамике формы клеток крови рептилий и их ядер, а также позволяет получить количественные характеристики лейкоци-
Таблица 3. Сравнительный анализ морфометрических параметров клеток крови Testudo horsfieldii (1), Geochelone elegans (2) и Chelonoidis carbonarius (3)
|
Клетки крови |
Н - критерий Краскела - Уоллиса |
Р - уровень значимости |
Z - критерий Данна |
р - уровень значимости |
|
E - индекс удлинения клетки |
||||
|
Эритроциты |
55,60 |
< 0,001 |
Z 1-2 =4,28 Z 1-3 = 7,40 Z 2-3 = 3,11 |
p 1-2 <0,001 p 1-3 < 0,001 p 2-3 = 0,005 |
|
Тромбоциты |
29,96 |
< 0,001 |
Z 1-2 = 3,29 Z 1-3 = 4,96 Z 2-3 = 2,43 |
p 1-2 =0,002 p 1-3 < 0,001 p 2-3 = 0,044 |
|
Гетерофилы |
0,66 |
0,71 |
- |
|
|
Эозинофилы |
10,35 |
0,005 |
Z 1-2 = 2,67 |
p 1-2 = 0,02 |
|
Базофилы |
4,38 |
0,11 |
- |
- |
|
Моноциты |
2,77 |
0,24 |
- |
- |
|
Лимфоциты |
3,62 |
0,19 |
- |
- |
|
е - индекс удлиненности ядра |
||||
|
Эритроциты |
9,76 |
0,007 |
Z 1-2 = 3,07 |
p 1-2 = 0,006 |
|
Тромбоциты |
8,62 |
0,01 |
Z 1-2 = 2,86 |
p 1-2 = 0,01 |
|
Гетерофилы |
17,33 |
0,0002 |
Z 1-2 = 2,53 Z 1-3 = 3,93 |
p 1-2 =0,03 p 1-3 = 0,0002 |
|
Эозинофилы |
1,45 |
0,48 |
- |
- |
|
Базофилы |
1,60 |
0,44 |
- |
- |
|
Моноциты |
1,38 |
0,50 |
- |
- |
|
Лимфоциты |
2,07 |
0,35 |
- |
- |
|
Площадь клетки, мкм2 |
||||
|
Эритроциты |
227,80 |
< 0,001 |
Z 1-2 = 14,57 Z 1-3 = 9,88 Z 2-3 = 4,69 |
p 1-2 <0,001 p 1-3 < 0,001 p 2-3 < 0,001 |
|
Тромбоциты |
11,40 |
0,003 |
Z 1-2 =2,51 Z 1-3 = 2,65 |
p 1-2 =0,03 p 1-3 = 0,02 |
|
Гетерофилы |
41,94 |
< 0,001 |
Z 1-2 = 6,34 Z 2-3 = 3,23 |
p 1-2 < 0,001 p 2-3 = 0,003 |
|
Эозинофилы |
3,89 |
0,14 |
- |
- |
|
Базофилы |
38,68 |
< 0,001 |
Z 1-2 =2,95 Z 1-3 =3,94 Z 2-3 =6,09 |
p 1-2 =0,009 p 1-3 = 0,0002 p 2-3 < 0,001 |
|
Моноциты |
8,37 |
0,01 |
Z 2-3 =2,79 |
p 2-3 =0,015 |
|
Лимфоциты |
14,56 |
0,0007 |
Z 1-3 = 2,83 Z 2-3 = 3,73 |
p 1-3 =0,013 p 2-3 = 0,0005 |
|
Площадь ядра, мкм2 |
||||
|
Эритроциты |
218,48 |
< 0,001 |
Z 1-2 = 13,57 Z 1-3 = 10,88 Z 2-3 = 2,68 |
p 1-2 < 0,001 p 1-3 < 0,001 p 2-3 = 0,021 |
|
Тромбоциты |
32,50 |
< 0,001 |
Z 1-2 = 5,15 Z 1-3 = 2,89 |
p 1-2 < 0,001 p 1-3 = 0,01 |
|
Гетерофилы |
82,32 |
< 0,001 |
Z 1-2 =8,19 Z 1-3 = 6,51 |
p 1-2 < 0,001 p 1-3 < 0,001 |
|
Эозинофилы |
1,98 |
0,36 |
- |
- |
|
Базофилы |
40,66 |
< 0,001 |
Z 1-2 = 3,03 Z 1-3 = 4,04 Z 2-3 = 6,24 |
p 1-2 =0,007 p 1-3 = 0,0001 p 2-3 <0,001 |
|
Моноциты |
4,49 |
0,10 |
- |
- |
|
Лимфоциты |
7,40 |
0,02 |
Z 1-3 =2,54 |
p 1-3 =0,032 |
Примечание: По критерию Z в таблице приведены только статистически значимые различия
Таблица 4. Сравнительный анализ ядерно-цитоплазматического отношения (ЯЦО) клеток крови сухопутных черепах из Самарского зоопарка:
Testudo horsfi eldii (1), Geochelone elegans (2) и Chelonoidis carbonarius (3)
Авторы выражают признательность сотрудникам Самарского зоопарка А.М. Балтушко, Н.А. Березину, Д.А. Казандаеву, А.Е. Кузовенко и А.А. Маряшину за организацию условий для забора крови у рептилий.
Список литературы Сравнительный лейкоцитарный профиль и размерные характеристики клеток крови сухопутных черепах из Самарского зоопарка
- Бондаренко, Д.А. Итоги изучения распространения, систематики и экологии среднеазиатской черепахи, Agrionemys horsfieldii (Gray, 1844) (Testudines, Testudinidae) / Д.А. Бондаренко // Труды Института зоологии Республики Казахстан». – Том 1, вып. 1. – Алматы: Институт зоологии Республики Казахстан, 2021. С. 37-70.
- Бондаренко, Д.А. Термобиология и суточная активность среднеазиатской черепахи (Agrionemys horsfi eldi) (Testudinidae, Reptilia) / Д.А. Бондаренко, Е.А. Перегонцев // Современная герпетология. – 2019. – Т. 19, вып. 1/2. – С. 7-30.
- Соколина, Ф.М. Гематология пресмыкающихся. Методическое пособие по курсу герпетология, большому практикуму и спецсеминарам / Ф.М. Соколина, А.В. Павлов, Р.Х. Юсупов. – Казань: Казанский государственный университет, 1997. – 31 с.
- Хайрутдинов, И.З. Характеристика крови рептилий и ее связь с условиями обитания: учебно-методическое пособие к курсу «Герпетология» / И.З. Хайрутдинов, Ф.М. Соколина. – Казань: Казанский университет, 2010. – 44 с.
- Alleman A.R., Jacobson E.R., Raskin R.E. Morphologic and cytochemical characteristics of blood cells from the desert tortoise (Gopherus agassizii) // American Journal of Veterinary Research. 1992. Vol. 53. P. 1645-1651.
- Bergamini B.C.S., Santos E.O., Wartchow B.S. et al. Hematologic Variation Values of Captive Red-footed Tortoise (Chelonoidis carbonaria) in South Brazil // Acta Scientiae Veterinariae. 2017. Vol. 45. P. 1426.
- Dash I. Microscopical analyses on characterization of peripheral blood cells of Western Pond turtle (Actinemys marmorata) (Baird and Girard, 1852) // Research Journal of Animal, Veterinary and Fishery Sciencesol. 2021. Vol. 9, no. 1. P. 25-30.
- Davis. A.K., Maney D.L., Maerz J.C. The use of leukocyte profi les to measure stress in vertebrates: A review for ecologists // Functional Ecology. 2008. Vol. 22. P. 760-772. (https://doi.org/10.1111/j.1365-2435.2008.01467.x).
- Fearon D.T., Locksley R.M. The instructive role of innate immunity in the acquired immune response // Science. 1996. Vol. 272. P. 50-54. (https://doi.org/10.1126/science.272.5258.50).
- Hoebe K., Janssen E., Beutler B. The interface between innate and adaptive immunity // Nature Immunology. 2004. Vol. 5. P. 971-974.
- International Guiding Principles for Biomedical Research Involving Animals. 2012. (https://grants.nih.gov/grants/olaw/guiding_principles_2012.pdf).
- Jacobson E.R., Garner M.M. (eds.). Infectious Diseases and Pathology of Reptiles. CRC Press, Tailor & Francis group, 2020. 1030 p. (https://doi.org/10.1201/9780429155567).
- Johnstone C. P., Reina R. D., Lill A. 2012. Interpreting indices of physiological stress in free-living vertebrates // Journal of Comparative Physiology B. Vol. 182. P. 861-879.
- Knotkova Z., Doubek J., Knotek Z., Hajkova P. Blood Cell Morphology and Plasma Biochemistry in Russian Tortoises (Agrionemys horsfi eldi) // Acta Veterinaria Brno. 2002. Vol. 71. P. 191-198. (DOI:10.2754/avb200271020191).
- Kophamel S., Rudd D., Ward L. C., Shum E. et al. Haematological and biochemical reference intervals for wild green turtles (Chelonia mydas): a Bayesian approach for small sample sizes // Conservation Physiology. 2022. Vol. 10, no 1. (coac043; doi:10.1093/conphys/coac043).
- Lee K.A. Linking immune defenses and life history at the levels of the individual and the species // Integrative and Comparative Biology. 2006. Vol. 46. P. 1000-1015.
- Martinez-Silvestre A. Morphology, cytochemical staining, and ultrastructural characteristics of the blood cells of the giant lizard of El Hierro (Gallotia simonyi) // Research in Veterinary Science. 2005. Vol. 78, no. 2. P. 127-134. (doi: 10.1016/j.rvsc.2004.07.009).
- Martins G.S., Alevi K.C., Azeredo-Oliveira M.T., Bonini-Domingos C.R. Cytochemical characteristics of blood cells from Brazilian tortoises (Testudines: Testudinidae) // Genetics and Molecular Research. 2016. Vol. 15, no. 1. (doi: 10.4238/gmr.15017549).
- Red List (Version 2022-1). 2022. (https://www.iucnredlist.org/assessment/red-list-index).
- Saint Girons M.-C. Morphology of the Circulating Blood Cell // Biology of the Reptilia. Vol. 3. Morphology C. London; New: York Academic Press, 1970. P. 73-91.
- Samour J.H., Howlett J.C., Silvanose C. et al. Normal haematology of free-living green sea turtles (Chelonia mydas) from the United Arab Emirates // Comparative Haematology International. 1998. Vol. 8, № 2. P. 102-107. (DOI:10.1007/BF02642499 102–107).
- Sposato P., Keating P., Lutz P. L., Milton S.L. Evaluation of immune function in two populations of green sea turtles (Chelonia mydas) in a degraded versus a nondegraded habitat // Journal of Wildlife Diseases. 2021. Vol. 57, no. 4. P. 761-772. (DOI: 10.7589/JWD-D-20-00204/).
- Uetz P., Freed P., Aguilar R., Hošek J. (eds.) The Reptile Database. 2023. (http://www.reptiledatabase.org).
- Wood F.E., Ebanks G.K. Blood cytology and hematology of the green sea turtle, Chelonia-mydas // Herpetologica. 1984. Vol. 40. P. 331-336.
