Сравнительный морфологический анализ лесостепной и степной популяций прыткой ящерицы Lacerta agilis в Новосибирской области

Автор: Симонов Е.П.
Журнал: Самарская Лука: проблемы региональной и глобальной экологии @ssc-sl
Рубрика: Научные сообщения
Статья в выпуске: 1 т.18, 2009 года.
Бесплатный доступ
Рассматриваются внутри- и межпопуляционные отличия окраски, рисунка, щиткования и флуктуирующей асимметрии лесостепной и степной популяций прытких ящериц из Новосибирской области. Показано, что окраска и рисунок более разнообразны у прытких ящериц из степной популяции, тогда как в лесостепной популяции преобладают отдельные вариации. Достоверные межпопуляционные отличия выявлены по 13 признакам, при этом существует две группы признаков демонстрирующих противоположную направленность изменений в отношении полимерности (расчлененности на элементы) фолидоза. Уровень флуктуирующей асимметрии достоверно выше в степной популяции, что подтверждает представление о том, что на территории Западной Сибири наиболее оптимальной для обитания вида является лесостепная зона.
Западная сибирь, морфологическая изменчивость, флуктуирующая асимметрия
Короткий адрес: https://sciup.org/148313170
IDR: 148313170
Текст краткого сообщения Сравнительный морфологический анализ лесостепной и степной популяций прыткой ящерицы Lacerta agilis в Новосибирской области
В настоящее время для Новосибирской области весьма актуальными в области герпетологии являются исследования эколого-морфологического характера, почти не осуществлявшиеся здесь ранее. Прыткая ящерица ( Lacerta agilis Linnaeus, 1758) – один из наиболее обычных и многочисленных видов рептилий на территории региона, особенно характерный для лесостепных и степных ландшафтов.
Данная работа выполнена в рамках исследования морфологических особенностей L. agilis из разных физико-географических районов Новосибирской области и изучения влияния климатических условий на особенности фолидо-за. На данном этапе главными задачами являлись: статистическая обработка полученных данных; внутрипопуляционный анализ; сравнительный морфологический анализ двух выборок (из лесостепной и степной зоны) прыткой ящерицы для выявления межпопуляционных отличий.
Материал для анализа был получен в результате полевых исследований в августе 2006 г. и в 2007 г. Данные по лесостепной популяции получены главным образом в мае-июне 2007 г. в окрестностях г. Новосибирска. Ящерицы, населяющие степь изучались 11 и 12 мая 2007 г. в окрестностях стационара Института систематики и экологии животных СО РАН, расположенного недалеко от пос. Троицкое Карасукского административного района Новоси-
∗ Евгений Петрович Симонов, студент.
бирской области, в 360 км к юго-западу от Новосибирска. Также были привлечены ранее не обрабатывавшиеся коллекционные материалы Зоологического Музея ИСЭЖ СО РАН (18 экз.).
Всего в анализе были использованы 26 особей (23 половозрелых и 3 ювенильных) прыткой ящерицы из степи и 40 особей (34 половозрелых и 6 ювенильных) из лесостепи. Изучение изменчивости окраски и рисунка проводилось только прижизненно и на половозрелых ящерицах (18 особей из степи и 21 из лесостепи). В соответствии с главными задачами исследования был сделан акцент на признаках характеризующих чешуйчатый покров животных. В качестве изучаемых признаков использовались традиционные показатели (Банников и др., 1977; Прыткая ящерица, 1976), и некоторые другие, обусловленные задачами исследования. Из них 5 метрических признаков: длина тела от кончика морды до переднего края клоакальной щели ( L ), хвоста ( L.cd. ), длина головы ( L.cap ), длина ( L.a. ) и ширина ( Lt.a. ) анального щитка; 17 меристических: число горловых чешуй ( G. ), число чешуй вокруг середины туловища ( Sq .), число чешуй вокруг 5 кольца хвоста ( Sq.c.cd .), число бедренных пор на одной ноге ( P.fm. ), количество преанальных щитков в первом и втором рядах ( Pre.an.-1 и Pre.an.-2 ), число поперечных рядов брюшных щитков ( Ventr. ), число задних носовых ( Na. ), скуловых щитков ( Lor. ) и их сумма ( Na.+Lor. ), количество нижнегубных ( S.lab .), нижнечелюстных ( НЧ ) и верхнегубных щитков до подглазничного ( Lab.1 ) и после него ( Lab.2 ), число увеличенных чешуй «воротника» ( Col .), количество верхнересничных щитков ( ВР ) и число щитков вокруг центральновисочного щитка ( ЩВЦВ ). А также рассчитывались 3 индекса: отношение ширины анального щитка к его длине ( Lt.a/L.a ), отношение длины головы к длине тела ( L.cap./L. ) и отношение длины тела к длине неповрежденного хвоста ( L./L.cd ). Особенности рисунка и окраски ящериц описаны по схеме, разработанной А.С. Барановым (1978), при этом доля каждой вариации (%) отражает частоту ее встречаемости во всей выборке. Оценка стабильности развития ящериц проведена путем анализа флуктуирующей асимметрии 10 меристических признаков ( P.fm., Na., Lor., Lab.1, Lab.2, S.lab., НЧ, ВР, ЩВЦ и дробление верхнегубных щитков). В качестве показателя стабильности развития использована средняя частота асимметричного проявления на признак ( ЧАПП ) (Методические рекомендации…, 2003).
Математическая обработка данных проведена общепринятыми методами (Лакин, 1990) с использованием электронных таблиц Excel 7.0 и пакета статистических программ Statistica for Windows 6.0. Для каждого признака определялось его среднее значение ( M ), ошибка среднего ( m ), среднее квадратичное отклонение ( σ ), ошибка среднего квадратичного отклонения ( S σ ), коэффициент вариации ( C.v. ) и его ошибка ( S cv ); различие средних оценено по критерию достоверности ( t ) Стьюдента. Для отдельных признаков рассчитывался коэффициент корреляции ( r ) с использованием поправки для малочисленных выборок, его достоверность ( t z ) определялась через z -преобразование Фишера (Лакин, 1990).
Внешняя окраска и рисунок тела прыткой ящерицы в Новосибирской об- ласти довольно консервативны и не демонстрируют всего диапазона измен- чивости. Как в лесостепи, так и в степи фон спинной поверхности у самок
Таблица 1
|
Встречаемость различных вариаций рисунка у прыткой ящерицы из лесостепной и степной популяций (%) ∗ |
|||
|
Вариация |
Частота в популяции |
P |
|
|
1 |
2 |
||
|
I |
4,8 |
22,2 |
> 0,05 |
|
Ll |
0,0 |
27,8 |
< 0,05 |
|
L |
95,2 |
50,0 |
< 0,01 |
|
L 1 |
4,8 |
22,2 |
> 0,05 |
|
L 2 |
0,0 |
5,6 |
> 0,05 |
|
L 3 |
81,0 |
44,4 |
< 0,05 |
|
D |
90,5 |
55,6 |
< 0,05 |
|
d |
0,0 |
11,1 |
> 0,05 |
|
Dd |
9,5 |
33,3 |
> 0,05 |
|
Dc |
4,8 |
5,6 |
> 0,05 |
|
Dn |
85,7 |
55,6 |
< 0,05 |
|
Dm |
14,3 |
33,3 |
> 0,05 |
|
Ds |
95,2 |
83,3 |
> 0,05 |
|
D 2 |
4,8 |
0,0 |
> 0,05 |
|
D 4 |
0,0 |
5,6 |
> 0,05 |
|
D 6 |
4,8 |
5,6 |
> 0,05 |
|
M |
100,0 |
100,0 |
> 0,05 |
|
M 1 |
23,8 |
44,4 |
> 0,05 |
|
M 2 |
76,2 |
55,6 |
> 0,05 |
|
M 3 |
76,2 |
44,4 |
< 0,05 |
|
M 4 |
23,8 |
55,6 |
< 0,05 |
∗ Примечание: 1 – лесостепная популяция; 2 – степная популяция. Вариации рисунка боковых спинных полос: l – отсутствие; Ll – прерывистость; L – наличие; L 1 – извилистость; L 2 – присутствие «веточек»; L 3 – захождение на головные щитки. Вариации рисунка центральной спинной полосы: D – наличие; d – отсутствие; Dd – прерывистость; Dc – темная; Dn – четкая; Dm – размытая; Ds – светлая; D 2 – образует утолщение в виде точки; D 4 – извилистая; D 6 – не доходит до головы. Характер пятнистости спины: M – наличие крупных пятен; М 1 – угловатые пятна; М 2 – округлые пятна; М 3 – пятна в светлой окантовке; М 4 – без окантовки.
всегда коричневый, у самцов основной цвет зеленый (50,0 и 66,7% соответственно; P > 0,05), реже оливковый (30,0 и 33,3%; P > 0,05), самцы с коричневой окраской были обнаруженный только в лесостепи (20,0%; P <0,05). В окраске горла самок лесостепной популяции встречаются белые (81,8%) и зеленые (18,2%; P < 0,05) тона, у степных – белые (66,7%) и коричнево-белые (33,3%; P < 0,01). У самцов – зеленая (90,0 и 66,7%; P > 0,05) либо оливковая (10,0 и 33,3%; P > 0,05). В степной популяции окраска брюха ящериц всегда соответствовала окраске горла; тогда как в лесостепной популяции 18,2 и 81,8% самок имеют коричневую и белую окраску брюха соответственно, а самцы в 10,0% случаев обладают белой окраской брюха, в 20,0% коричневой и в 70,0% – зеленой.
Частоты встречаемости различных вариаций рисунка у прыткой ящерицы из лесостепной и степной популяций представлены в табл. 1. Достоверные межпопуляционные отличия выявлены для 7 вариаций рисунка. Эти отличия носят скорее количественный характер (разные выборочные доли), нежели качественный (наличие или отсутствие отдельных вариаций). Так, вариация прерывистых боковых спинных полос ( Ll ) встречается только в степной популяции ( P < 0,05). По другим вариациям, обнаруженным лишь в одной из двух популяций ( L 2 , d, D 2 , D 4 ), достоверных отличий не обнаружено. В целом, рисунок
ящериц из степной популяции более разнообразен, тогда как в лесостепной популяции доминируют отдельные вариации: L 3 (81%), D (90,5%), Dn (85,7%),
M 3 (76,2%).
Все данные внутри- и межпопуляционного морфологического анализа представлены в табл. 2–4. По большинству признаков, для которых известен половой диморфизм у данного вида, были выявлены достоверные внутрипо- пуляционные отличия. За исключением количества чешуй вокруг середины туловища (Sq.) и вокруг 5-ого кольца хвоста (Sq.c.cd.), видимо, для выявления достоверных отличий по этим признакам необходим больший размер выборки. Кроме того, для ящериц из степной популяции обнаружено, что у самцов количество нижнечелюстных щитков (НЧ) в среднем достоверно больше чем у самок. При этом значение НЧ положительно коррелирует с длиной головы: r = 0,80 ± 0,164 (n = 23; P < 0,01). В лесостепной популяции такой корреляции не обнаружено (r = -0,02 ± 0,189; n = 29).
Таблица 2 Сравнительная морфологическая характеристика степной и
|
лесостепной популяций прыткой ящерицы из Новосибирской области |
||||||||
|
Признак |
Пол |
Степь |
Лесостепь |
P |
||||
|
n |
min – max M ± m |
P |
n |
min – max M ± m |
P |
|||
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
|
L. |
♂+♀ |
23 |
64–93 76,7 ± 1,47 |
– |
34 |
61–91 75,3 ± 1,44 |
– |
>0,05 |
|
L.cd |
♂+♀ |
14 |
91–142 115,4 ± 3,70 |
– |
22 |
103–156 120,8 ± 3,10 |
– |
>0,05 |
|
L.cap. |
♂ |
16 |
13,4–17,9 16,2 ± 0,27 |
<0,05 |
16 |
14,4–19,8 16,6 ± 0,41 |
<0,05 |
>0,05 |
|
♀ |
7 |
12,2–16,2 14,5 ± 0,56 |
18 |
13–17,9 15,4 ± 0,34 |
>0,05 |
|||
|
Ventr. |
♂ |
16 |
28–31 29,56 ± 0,258 |
<0,01 |
16 |
27–30 28,81 ± 0,228 |
<0,001 |
<0,05 |
|
♀ |
7 |
30–33 31,14 ± 0,404 |
18 |
29–33 30,78 ± 0,250 |
>0,05 |
|||
|
P.fm. |
♂+♀ |
26 |
9–16 13,27 ± 0,231 |
– |
39 |
12–16 13,92 ± 0,156 |
<0,05 |
|
|
Pre.an. 1 |
♂+♀ |
26 |
3–7 5,69±0,173 |
– |
33 |
4–7 5,82 ± 0,147 |
– |
>0,05 |
|
Pre.an. 2 |
♂+♀ |
26 |
9–15 12,08±0,308 |
– |
20 |
10–15 12,09 ± 0,201 |
– |
>0,05 |
|
Sq. |
♂ |
14 |
38–45 42,07±0,606 |
>0,05 |
13 |
41–50 44,31 ± 0,771 |
>0,05 |
<0,05 |
|
♀ |
7 |
39–47 42,29±1,322 |
15 |
40–49 43,13 ± 0,736 |
>0,05 |
|||
|
G. |
♂+♀ |
26 |
16–22 19,19±0,299 |
– |
36 |
16–22 19,22 ± 0,273 |
– |
>0,05 |
|
Col. |
♂+♀ |
26 |
7–11 9,31 ± 0,198 |
– |
40 |
9–13 10,60 ± 0,163 |
– |
<0,001 |
|
Sq.c.cd |
♂ |
14 |
28–34 31,38±0,473 |
>0,05 |
13 |
28–37 33,23 ± 0,652 |
>0,05 |
<0,05 |
|
♀ |
7 |
27–32 30,00±0,577 |
16 |
27–36 31,56 ± 0,645 |
>0,05 |
|||
|
Na. |
♂+♀ |
26 |
1–3 1,81 ± 0,100 |
– |
39 |
1–3 1,91 ± 0,045 |
– |
>0,05 |
|
Lor. |
♂+♀ |
26 |
1–4 2,08 ± 0,129 |
– |
39 |
0–3 1,38 ± 0,111 |
– |
<0,001 |
|
Na. + Lor. |
♂+♀ |
26 |
2–7 3,88 ± 0,193 |
– |
39 |
2–5 3,29 ± 0,102 |
– |
<0,01 |
|
Lab.1 |
♂+♀ |
26 |
4–5 4,10 ± 0,056 |
– |
39 |
4–5 4,04 ± 0,028 |
– |
>0,05 |
|
окончани е таблицы 2 |
||||||||
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
|
Lab.2 |
♂+♀ |
26 |
2–3 2,13±0,065 |
– |
34 |
1–3 2,09 ± 0,045 |
– |
>0,05 |
|
S.lab. |
♂+♀ |
26 |
6–8 6,35±0,117 |
– |
36 |
5–7 6,11 ± 0,057 |
– |
>0,05 |
|
НЧ |
♂ |
16 |
5–6 5,84±0,088 |
<0,001 |
14 |
5–6 5,32 ± 0,010 |
>0,05 |
<0,001 |
|
♀ |
7 |
5–7 5,07±0,071 |
15 |
5–6 5,43 ± 0,118 |
<0,05 |
|||
|
ВР |
♂+♀ |
26 |
4–6 4,96±0,062 |
– |
39 |
4–6 4,95 ± 0,055 |
– |
>0,05 |
|
ЩВЦВ |
♂+♀ |
26 |
6–10 7,77±0,195 |
– |
34 |
5–9 6,79 ± 0,140 |
– |
<0,001 |
|
L.cd. / L. |
♂ |
9 |
1,41–1,69 1,53 ± 0,036 |
>0,05 |
12 |
1,52–1,81 1,68 ± 0,024 |
<0,001 |
<0,01 |
|
♀ |
3 |
1,42–1,51 1,45 ± 0,030 |
10 |
1,24–1,59 1,49 ± 0,040 |
>0,05 |
|||
|
L.cap. / L. |
♂ |
16 |
0,20–0,22 0,21±0,002 |
<0,001 |
16 |
0,22–0,24 0,22 ± 0,002 |
<0,001 |
<0,001 |
|
♀ |
7 |
0,18–0,19 0,19±0,002 |
18 |
0,19–0,23 0,20 ± 0,003 |
<0,01 |
|||
|
L.ta. / L.a. |
♂ |
16 |
1,50–2,91 2,13±0,081 |
<0,05 |
14 |
1,72–2,32 1,90 ± 0,068 |
<0,001 |
<0,05 |
|
♀ |
7 |
1,29–2,5 1,71±0,161 |
16 |
1,39–2,10 1,53 ± 0,048 |
>0,05 |
|||
Примечание: данные по самцам и самкам представлены отдельно для признаков, по которым были обнаружены или известны по литературе половые отличия.
Достоверные межпопуляционные отличия обнаружены по 13 признакам (см. табл. 2): P.fm., Col., Lor., Na.+ Lor., ЩВЦВ, НЧ, L.cap./L. – как для самцов, так и для самок; Ventr., Sq., Sq.c.cd, L.cd./L., L.ta./L.a. – только для самцов, что обусловлено, вероятно, малым количеством самок в выборке из степи. Дробление верхнегубных щитков чаще происходит у ящериц из степной популяции (38,1% против 4,76%; P < 0,01).
Таблица 3 Частоты встречаемости различных комбинаций щитков в задненосовой области (Na./Lor.)
у прыткой ящерицы из лесостепной и степной популяций (%)
|
Популяция |
Задненосовая формула ( Na./Lor. ) |
||||||||||
|
1/1 |
1/2 |
1/3 |
2/0 |
2/1 |
2/2 |
2/3 |
2/4 |
3/2 |
3/3 |
3/4 |
|
|
Лесостепная |
0,00 |
6,41 |
3,85 |
10,26 |
44,87 |
33,33 |
0,00 |
0,00 |
1,28 |
0,00 |
0,00 |
|
Степная |
1,92 |
23,08 |
0,00 |
0,00 |
15,39 |
38,46 |
13,46 |
1,92 |
0,00 |
3,85 |
1,92 |
У двух особей из степной популяции были обнаружены следующие аберрации чешуйчатого покрова: мелкие щитки и зернышки между теменными, лобнотеменными, межтеменным и затылочным щитками; дробление лобнотеменных щитков; сросшиеся прианальные щитки в первом полукруге ( Pre.an. 1 ); срастание нескольких чешуй «воротника» ( Col. ) в его центральной части.
Анализ коэффициентов вариации ( C.v. ) демонстрирует сходную картину степени изменчивости меристических признаков в обеих популяциях (рисунок). Достоверные межпопуляционные отличия C.v. обнаружены только по 6 признакам из 17. При этом коэффициенты вариации выше в степной популяции по сравнению с лесостепной у P.fm., Pre.an. 2, Na., Lab., S.lab., и ниже у Lor .
Таблица 4
Частоты встречаемости числа щитков в задненосовой области ( Na.+Lor. ) у прыткой ящерицы из лесостепной и степной популяций (%)
|
Популяция |
Количество щитков в задненосовой области ( Na. + Lor. ) |
|||||
|
2 |
3 |
4 |
5 |
6 |
7 |
|
|
Лесостеп ная |
10,26 |
51,28 |
37,18 |
1,28 |
0,00 |
0,00 |
|
Степная |
1,92 |
38,47 |
38,46 |
13,46 |
5,77 |
1,92 |
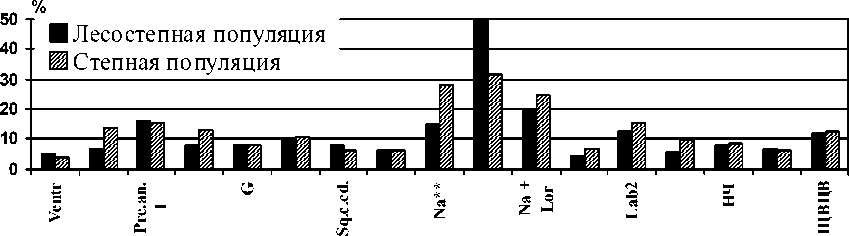
Рис. Коэффициенты вариации меристических признаков у прыткой ящерицы из лесостепной и степной популяций ( C.v., %)
Примечание: * – P < 0,05; ** – P < 0,01.
При изучении флуктуирующей асимметрии билатеральных структур были получены следующие значения ЧАПП : для степной популяции ЧАПП = 0,27 ± 0,029 ( n = 21), для лесостепной ЧАПП = 0,19 ± 0,025 ( n = 20). Значения ЧАПП достоверно отличаются ( P < 0,05). Полученные показатели ЧАПП подтверждают представление о том, что на территории Западной Сибири оптимальной для обитания вида является лесостепная зона (Булахова, 2004).
Таким образом, в результате проведенного исследования были выявлены признаки, для которых характерен половой диморфизм, и признаки, по которым существуют достоверные отличия между лесостепной и степной популяциями прыткой ящерицы. На данном этапе исследования можно говорить о двух группах признаков демонстрирующих противоположную направленность изменений в отношении полимерности (расчлененности на элементы) фолидоза: Ventr., Lor., Na. + Lor., ЩВЦВ, НЧ – увеличение в степенной популяции; Sq., Sq.c.cd., Col. – сокращение в степной популяции. При этом наиболее явную тенденцию к расчленению на отдельные элементы демонстрируют топографически близкие признаки щитко-вания головы, а в противоположном направлении – количество увеличен- ных чешуй «воротника», о чем так же свидетельствуют обнаруженные в степной популяции аберрации чешуйчатого покрова и более высокая частота расщепления верхнегубных щитков.
Увеличение полимерности чешуйчатого покрова в степной популяции для ряда признаков ( Ventr., Lor., Na.+ Lor., ЩВЦВ, НЧ ), по всей вероятности, обусловлено климатическими факторами (более высокие средние температуры, более низкая влажность и т.п.), эффект от которых формируется во время развития эмбрионов в яйцах (Хонякина, Ферхатова, 1977; Захаров, 1981; Ройтберг, 1981, 1989). Более подробно этот аспект и его роль в формировании клинальной изменчивости будут рассмотрены в последующих публикациях, когда для интерпретации полученных результатов будут использованы аналогичные данные по морфологии более северных популяций L. agilis.
Список литературы Сравнительный морфологический анализ лесостепной и степной популяций прыткой ящерицы Lacerta agilis в Новосибирской области
- Банников А.Г., Даревский И.С., Ищенко В.Г., Рустамов А.К., Щербак Н.Н. Определитель земноводных и пресмыкающихся фауны СССР. М.: Просвещение, 1977. 414 с.
- Баранов А.С. Выделение фенов по окраске у рептилий (на примере прыткой ящерицы) // Физиология и популяционная экология животных. Саратов, 1978. № 5. С. 68-72.
- Булахова Н.А. Ящерицы (Reptilia, Squamata, Lacertidae) юго-востока Западной Сибири (география, экология, морфология). Автореф. дис....канд. биол. наук. Томск, 2004. 27 с.
- Захаров В.М. Влияние температуры инкубации на фенотипическую изменчивость прыткой ящерицы (Lacerta agilis) // Вопросы герпетологии. Л.: Наука, 1981. С. 56-57.
- Лакин Г.Ф. Биометрия. М.: Высшая школа, 1990. 352 с.
- Методические рекомендации по выполнению оценки качества среды по состоянию живых существ (оценка стабильности развития живых организмов по уровню асимметрии морфологических структур). М.: Росэкология, 2003. 25 с.
- Прыткая ящерица. М.: Наука, 1976. 376 с.
- Ройтберг Е.С. Анализ внутри- и межпопуляционных различий по меристическим признакам фолидоза у прыткой и полосатой ящериц (Lacerta agilis и L. strigata) Дагестана // Вопросы герпетологии. Л.: Наука, 1981. С. 115-116.
- Ройтберг Е.С. О некоторых проявлениях изменчивости фолидоза ящериц рода Lacerta (К вопросу о роли механизмов морфогенеза в эволюции) // Вопросы герпетологии. Киев: Наукова думка, 1989. С. 212-213.
- Хонякина З.П., Ферхатова З.А. Особенности популяций полосатой ящерицы из низменного и предгорного районов Дагестана // Вопросы герпетологии. Л.: Наука, 1977. С. 216-217.
