Стабилизация соотношения между числом видов растений разного микоризного статуса – один из аттракторов прогрессивных сукцессий?

Автор: Веселкин Денис Васильевич
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Структура и динамика растительных сообществ
Статья в выпуске: 1-5 т.14, 2012 года.
Бесплатный доступ
Проанализированы закономерности участия растений разного микоризного статуса в сукцессиях растительности в степной и лесостепной зонах. В ходе прогрессивных сукцессий в составе фитоценозов возрастает доля облигатно микоризных видов и снижается доля немикоризных и факультативно микоризных видов. Соотношение видов разного микоризного статуса варьирует в сукцессионно зрелых сообществах в узких пределах: 5–10 % немикоризных видов; 15–20 % факультативно микоризных; 70–80 % облигатных микоризных. В раннесукцессионных сообществах изменчивость этого соотношения существенно выше.
Сукцессии, растительность, микориза, арбускулярная микориза, немикоризные растения, микоризные растения
Короткий адрес: https://sciup.org/148200787
IDR: 148200787 | УДК: 581.557.24
The proportion stabilization between plant species with different mycorrhizal status – one of the attractors of progressive successions?
Characterized by the participation of plants with different mycorrhizal status in several successions in the steppe and forest-steppe zones. During progressive successions the share of mycorrhizal plants species increases while the share of nonmycorrhizal and optional mycorrhizal plants species falls. Ratio of plants with different mycorrhizal status varies in the later successional stages within the narrow limits: nonmycorrhizal species – 5–10%; optional mycorrhizal species – 15–20 %; mycorrhizal species – 70–80 %. In the early successional stages the variability of this ratio is significantly higher.
Текст научной статьи Стабилизация соотношения между числом видов растений разного микоризного статуса – один из аттракторов прогрессивных сукцессий?
В разных концепциях сукцессий растительности с той или иной ясностью выражены представления об идеальных свойствах сукцессионно зрелых фитоценозов или экосистем. К таким свойствам относятся такие очевидные как: преобладание многолетних видов растений; максимизация биомассы особей и фитоценозов при выравнивании соотношения продукция/дыхание и максимизация массы органического вещества, депонированного в экосистеме; усиление межвидовой конкуренции или дифференциации экологических ниш. Подобные свойства и их конкретные численные характеристики можно считать аттракторами в соответствие с общим пониманием аттрактора как идеального состояния, к которому стремится система в своем развитии. Еще одно, возможно общее, свойство сукцессионно зрелых растительных сообществ – значительная стабильность соотношения между числом видов растений разного микоризного статуса.
Со спецификой микоризных взаимодействий связывают некоторые механизмы сукцессий [22, 24], чаще всего – различия в конкурентоспособности растений в отношении почвенного питания. Этим обстоятельством обусловлен интерес к изучению микоризных связей в ходе динамики фитоценозов. Работы на стыке ботаники, микологии и экологии, затрагивающие вопросы динамики и значения микориз при сукцессионных перестройках, преимущественно посвящены или анализу мико-ризности ограниченного числа модельных видов на разных этапах сукцессий [20, 25] или описаниям микориз у большего числа видов в ходе сукцессий, зачастую довольно специфических, таких как постледниковые [19, 23] или поствулканические [18, 21]. В методическом отношении последний способ можно обозначить как получение прямых оценок участия растений разной микоризности в сукцессиях. Возможен и иной – косвенный – способ изучения сукцессионной приуроченности растений раз-
ного микоризного статуса. В соответствие с ним оценки микоризного статуса базируются на компиляции литературных данных, например [3, 4]. В настоящем сообщении преследуется цель продемонстрировать, что заключение о стабильности соотношения между числом видов растений разного микоризного статуса в сукцессионно зрелых растительных сообществ не существенно зависят от методического подхода и подтверждаются как методами прямого наблюдения микориз в ходе сукцессий, так и косвенными.
Прямые оценки микоризного статуса видов в ходе сукцессий на разновозрастных отвалах угледобывающих предприятий степных и лесостепных районов взяты из работы Т.С. Чибрик с соавторами [14]. Из этой же публикации транслирована относительная шкала сукцессионной продвинутости растительных сообществ отвалов: простая или сложная растительные группировки или фитоценоз. Дополнительно привлечены сведения о соотношении немикоризных и микоризных видов в ненарушенных лесостепных и степных фитоценозах [12, 13]. Для характеристики микоризного статуса использована простая дихотомия оценок: особи того или иного вида в каком-либо местообитании могут быть либо немикоризными либо микоризными и один и тот же вид в разных условиях может учитываться в разных статусах.
В ходе прогрессивных сукцессий на техногенных субстратах доля микоризных видов последовательно возрастает (рис. 1), а немикоризных, соответственно, снижается. В сукцессионно молодых простых группировках на микоризные растения приходится примерно половина состава – 40–65 % видов. В фитоценозах на отвалах доля микотрофов увеличивается до четверти – 65–75 %, а в зональных фитоценозах – до 75–90 %. Эти оценки достаточно красноречивы, различия между ними значимы и утверждение об увеличении в ходе сукцессий доли микоризных видов можно, по-видимому, считать надежным и общим. Но важно отметить, что списки видов растений, проанализированных на микоризность, по крайней мере, в некоторых ме- стообитаниях неполны. Например, доступные нам оценки для зональных фитоценозов основываются на анализе лишь 20-45 видов в каждом из них, что, по всей вероятности, меньше общего числа видов в данных фитоценозах. Поэтому нельзя исключать смещения оценок соотношений между числом немикоризных и микоризных видов и необходимо сравнение намеченных закономерностей с закономерностями, наблюдаемыми при косвенном установлении микоризного статуса.
Косвенные оценки микоризного статуса видов использованы при анализе нескольких сукцессий: формирование «агростепи» в Башкирском степном Зауралье [1]; формирование пионерной растительности возле Медногорского медно-серного комбината в Оренбургской области [17]; зарастание отвалов вскрышных пород в сухих степях Казахстана
[8], степях Оренбургской области [15], отвалов металлургического производства на Центральном Кавказе в Кабардино-Балкарской Республике [5]. Этапы сукцессии разделили на ранние, средние и поздние, варьируя критерии их выделения в зависимости от методических схем, реализованных в разных исследованиях. Данные о микоризообразо-вании аккумулированы более чем из 30 публикаций, основные из которых [6, 10, 11, 16]. Виды разделили на: 1) немикоризные, не образующие микориз в соответствие со всеми источниками; 2) факультативно микоризные, образующие микоризы по одним оценкам и не образующие по другим; 3) микоризные (облигатно микоризные), постоянно формирующие микоризы, чаще всего, арбускуляр-ные.
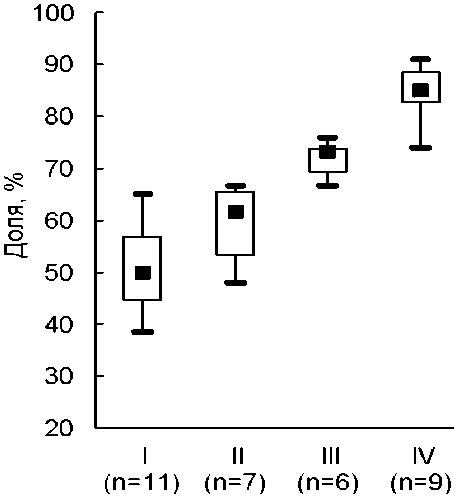
Рис. 1 . Доля микоризных видов в ходе зарастания отвалов (простая (I) и сложная (II) растительная группировка, III - фитоценоз) и в зональных фитоценозах степной и лесостепной зон (IV). Здесь и на рис. 2: числа в скобках - количество проанализированных сообществ; квадрат - медиана, прямоугольник - межквартильный, вертикальные линии - абсолютный размах.
Оценки с использованием метода косвенной идентификации микоризного статуса однозначно подтверждают закономерность направленного изменения соотношения видов разного микоризного статуса в ходе сукцессий (рис. 2). По мере приближения фитоценозов по составу и сложению к зональным в них возрастает участие облигатно микоризных видов при соответствующем снижении долей менее тесно связанных с арбускулярными грибами растений - факультативно микоризных и немикоризных.
Необходимо отметить большую вариабельность оценок участия видов разной микоризности на ранних этапах сукцессий и их меньшую вариабельность на поздних этапах. Высокая изменчивость в раннесукцессионных сообществах иллюстрируется большим размахом долей немикоризных и факультативно микоризных видов и объясняется специ- фикой условий среды и состава фитоценозов в разных районах. При рассмотрении сукцессионно продвинутых сообществ различия между разными районами в значительной степени нивелируются и соотношение видов разного микоризного статуса оказывается близким. Высокая изменчивость микоризной структуры сукцессионно молодых сообществ и относительная стабильность такой структуры в сукцессионно поздних сообществах очевидно отражает различие принципов их организации [7, 9]. На первых этапах сукцессий популяции разных видов не только слабо взаимодействуют между собой, но и могут следовать разным приспособительным стратегиям. Позднее в силу стабилизации и выравнивания условий выравнивается и стабилизируется спектр экоморф, что оказывается коррелятивно связанно со стабилизацией пропорций между видами разного микоризного статуса.
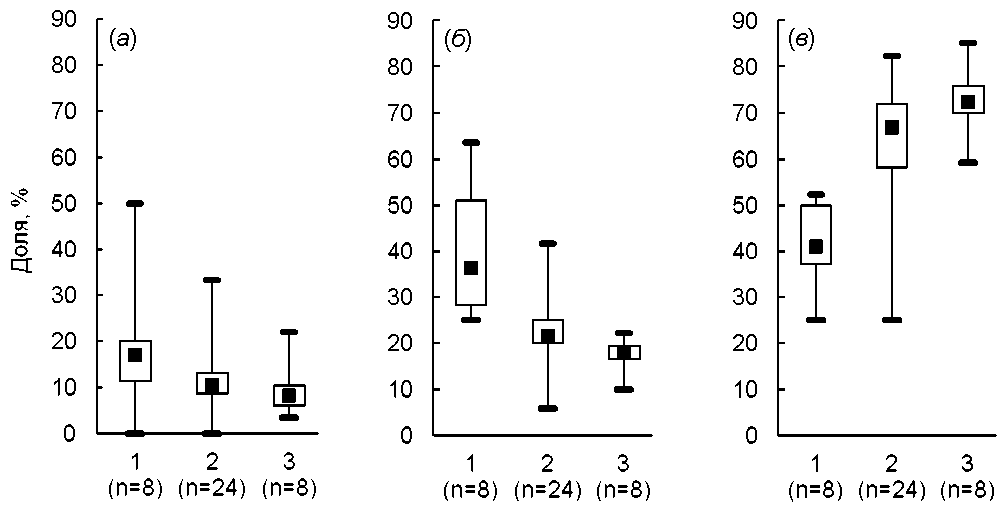
Рис. 2 . Доля немикоризных ( а ), факультативно ( б ) и облигатно ( в ) микоризных видов на ранних (1), промежуточных (2) и поздних (3) этапах прогрессивных сукцессий в степной и лесостепной зонах.
Увеличение доли микоризных видов – общее направление прогрессивных сукцессий в степной и лесостепной зонах. Этот феномен ясно воспроизводится как в исследованиях, основанных на прямых оценках микоризного статуса, так и при косвенной идентификации способности растений к микоризо-образованию. Это заключение соответствует гипотезе о сукцессионной специфике растений с разными стратегиями почвенного питания [24], частными случаями которой являются, например, высокая теснота микоризных связей у доминантов [12] и высокая встречаемость немикоризных видов среди растений рудеральной стратегии Грайма– Раменского [2]. Достаточная узость границ изменчивости соотношения числа видов разных микоризных стратегий в сукцессионно продвинутых сообществах позволяет рассматривать это соотношение (5–10 % немикоризных видов; 15–20 % факультативно микоризных; 70–80 % облигатных ми-коризообразователей) как устойчивое состояние признака – аттрактор развития «трофической» структуры фитоценоза. Такая интерпретация заставляет предполагать существование причин устойчивости пропорций между группами видов разной микоризности, т.е. существование функциональной зависимости между способностью образовывать микоризы и ценотическим положением видов растений.
Работа выполнена при поддержке Программы развития ведущих научных школ (НШ-5325.2012.4).
Список литературы Стабилизация соотношения между числом видов растений разного микоризного статуса – один из аттракторов прогрессивных сукцессий?
- Абдуллин М.Р., Миркин Б.М. Опыт создания «агростепей» в Башкирском степном Зауралье//Бюл. МОИП. Отд. биол. 1995. Т. 100. № 5. С. 77-84.
- Бетехтина А.А., Веселкин Д.В. Распространенность и интенсивность микоризообразования у травянистых растений Среднего Урала с разными типами экологических стратегий//Экология. 2011. № 3. С. 176-182.
- Веселкин Д.В. Закономерности участия растений разного микотрофного статуса в антропогенно индуцированных сукцессиях в степях//Вестник Курганского гос. ун-та. Сер.: Естественные науки. 2012. Т. 2. № 21. С. 75-80.
- Веселкин Д.В., Бетехтина А.А. Участие растений разного микотрофного статуса в техногенно обусловленных сукцессиях в степной зоне Урала//Вестник ОГУ. 2011. № 12. (131). С. 44-47.
- Высокогорная геоэкология в моделях/М.Ч. Залиханов и др. Отв. ред.: Г.С. Розенберг, Г.К. Сафаралиев. М.: Наука, 2010. 487 с.
- Елеусенова Н.Г., Селиванов И.А. Микотрофность растений во флоре северных пустынь Казахстана//Микориза растений. Уч. зап. Перм. гос. пед. ин-та. Т. 112. Пермь, 1973. С. 100-111.
- Крышень А.М. К вопросу о механизмах устойчивости и развития растительных сообществ//Актуальные проблемы геоботаники. III Всерос. школа-конф. Лекции. Петрозаводск, 2007. С. 157-175.
- Куприянов А.Н., Манаков Ю.А. Динамика зарастания отвала вскрыши бывшего Федоровского угольного месторождения за 30 лет//Рекультивация нарушенных земель в Сибири. Вып. 3. Кемерово: КРЭОО "Ирбис", 2008. С. 45-55.
- Миркин Б.М., Наумова Л.Г. Наука о растительности (история и современное состояние основных концепций). Уфа: Гилем, 1998. 413 с.
- Нозадзе Л.М. Микориза растений некоторых пустынных и степных фитоценозов Восточной Грузии//Микориза и другие формы консортивных связей в природе. Пермь: ПГПИ, 1989. С. 44-49.
- Селиванов И.А., Бейрах Э.А., Мельникова С.Л., Саламатова Н.Г. К инвентаризации микотрофных растений лесостепного Зауралья//Уч. зап. Перм. госуниверситета. Т. 114. Биология. Пермь, 1964. С. 63-78.
- Селиванов И. А. Микосимбиотрофизм как форма консортивных связей в растительном покрове Советского Союза. М.: Наука, 1981. 232 с.
- Утемова Л.Д. О микотрофности некоторых растений юга Красноярского края//Уч. зап. Перм. гос. пед. ин-та. Т. 112. Пермь, 1973. С. 94-99.
- Чибрик Т.С., Нагибина Т.И., Рябкова Т.Е. О микотрофности растений на отвалах угольных разработок Урала//Растения и промышленная среда. Свердловск, 1980. С. 33-79.
- Чибрик Т.С., Елькин Ю.А. Формирование фитоценозов на нарушенных промышленностью землях: (биологическая рекультивация). Свердловск: Изд-во Урал. ун-та, 1991. 220 с.
- Чибрик Т.С., Саламатова Н.А. Микосимбиотрофизм культурфитоценозов Коркинского угольного разреза//Растения и промышленная среда. Свердловск, 1985. С. 54-69.
- Шилова И.И., Лукьянец А.И. Сукцессии степной растительности на территориях, подверженных аэротехногенному воздействию предприятий цветной металлургии//Растительность в условиях техногенных ландшафтов Урала. Свердловск: УрО АН СССР, 1989. С. 56-79.
- Ahulu E.M., Nakata M., Nonaka M. Arumand Paris-type arbuscular mycorrhizas in a mixed pine forest on sand dune soil in Niigata Prefecture, central Honshu, Japan//Mycorrhiza. 2005. Vol. 15. № 2. P. 129-136.
- Chapin F.S., Walker L.R., Fastie C.L., Sharman L.C. Mechanisms of primary succession following deglaciation at Glacier Bay, Alaska//Ecol. Monogr. 1994. Vol. 64. № 2. P. 149-175.
- Daft J., Nicolson T.N. Arbuscular mycorrhizas in plants colonizing coal wastes in Scotland//New Phytol. 1974. Vol. 73. № 6. P. 1129-1138.
- Gemma J.N., Koske R.E. Mycorrhizae in recent volcanic substrates in Hawaii//Amer. J. Bot. 1990. Vol. 77. № 9. P. 1193-1200.
- Heijden van der M.G.A., Klironomos J.N., Ursic M. et al. Mycorrhizal fungal diversity determines plant biodiversity, ecosystem variability and productivity//Nature. 1998. Vol. 396. P. 69-72.
- Jumpponen A., Trappe J.M., Cazares E. Occurrence of ectomycorrhizal fungi on the forefront of retreating Lyman Glacier (Washington, USA) in relation to time since deglaciation//Mycorrhiza. 2002. Vol. 12. № 1. P. 43-49.
- Lambers H., Raven J.A., Shaver G.R., Smith S.E. Plant nutrient-acquisition strategies change with soil age//Trends Ecol. Evol. 2008. Vol. 23. № 2. P. 95-103.
- Miller R.M. Some occurrences of vesicular-arbuscular mycorrhiza in natural and disturbed ecosystems of the Red Desert//Can. J. Bot. 1979. Vol. 57. № 6. P. 619-623.
