Старение симбиотического клубенька у бобовых растений: молекулярно-генетические и клеточные аспекты (обзор)
")
Автор: Серова Т.А., Цыганов В.Е.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Обзоры, проблемы, итоги
Статья в выпуске: 5 т.49, 2014 года.
Бесплатный доступ
Старение - естественный этап в развитии симбиотического клубенька. В результате процесса старения происходит реутилизация различных питательных веществ из клубенька в другие органы растения. В большинстве случаев старение запускается после окончания цветения бобовых растений, хотя первые признаки старения могут наблюдаться очень рано в развитии клубенька. Задержка запуска программы старения позволила бы продлить период активной азотфиксации, а, следовательно, увеличить содержание симбиотрофного азота в растениях, и, в конечном счете, повысить урожайность бобовых растений. Поэтому неудивительно, что в последние годы старения азотфиксирующих клубеньков активно изучается. В обзоре рассматриваются основные этапы развития азотфиксирующего симбиотического клубенька бобовых растений, особенности развития симбиотических клубеньков детерминированного и недерминированного типов. У бобовых растений с недерминированными клубеньками время жизни симбиосом невелико, потому что центральные инфицированные ткани постоянно обновляются за счет апикальной меристемы. Выделяют две последовательные стадии в процессе старения недерминированного клубенька: сначала происходит деградация бактероидов и гибель нескольких растительных клеток, затем полностью разрушаются и симбиосомы, и инфицированные клетки. В детерминированных клубеньках старение инициируется в центральной зоне клубенька и распространяется к периферии. Анализируются морфологические признаки старения симбиотического клубенька на ультраструктурном уровне. Обсуждается роль в старении симбиотического клубенька цистеиновых и треониновых протеаз. Деградация белка, которая позволяет повторно утилизировать азот и другие вещества, - вероятно наиболее важный из процессов, происходящих на протяжении старения. Существуют доказательства, что в корневых клубеньках бобовых растений цистеиновые протеазы вовлекаются в функционирование клубенька, в адаптацию клеток хозяина к физиологическим стрессам и в контроль старения клубенька. При широкомасштабном анализе транскриптома клубеньков люцерны ( Medicago truncatula Gaertn.) выявлены несколько групп генов, активность которых характерна для последовательных стадий старения недерминированного клубенька. Рассматривается роль в старении таких фитогормонов как этилен, абсцизовая и жасмоновая кислоты, гиббереллины и монооксид азота. Следует отметить, что до настоящего времени представления о гормональной регуляции процесса старения остаются достаточно фрагментарными. Анализируется окислительный стресс, сопровождающий процесс старения клубенька. Показано, что с возрастом клубенька содержание пероксидов, карбонилов белков, модифицированных оснований ДНК и каталитического железа увеличивается. Железо, содержащееся в перибактероидном пространстве, усиливает перекисное окисление липидов, что может способствовать деградации перибактероидной мембраны в стареющих клубеньках. Количество окисленных форм глутатиона и гомоглутатиона значительно повышается по мере развития клубенька, восстановленных - снижается при старении, что свидетельствует о развитии окислительного стресса в стареющих клубеньках. В обзоре также рассматривается роль генов, кодирующих белки, вовлеченные в транспорт широкого круга молекул, и генов, продукты которых выполняют регуляторные и сигнальные функции в клетке; отличие стресс-индуцированного старения от естественного процесса. С использованием модельных бобовых Lotus japonicas (Regel) K. Larsen и M. truncatula были клонированы некоторые гены, мутации по которым приводят к инициации раннего старения. Важно отметить, что эти гены кодируют самые разные белки, участвующие в функционировании симбиотического клубенька. К настоящему времени у M. truncatula выявлены два транскрипционных фактора, задействованных в процессе старений симбиотического клубенька. При этом отмечено, что индуцированное старение происходит быстрее естественного и имеет признаки окислительного стресса и запрограммированной клеточной гибели.
Бобово-ризобиальный симбиоз, развитие клубенька, генетический контроль, окислительный стресс, протеазы, этилен, абсцизовая кислота, жасмоновая кислота, гиббереллины, монооксид азота, ризобии
Короткий адрес: https://sciup.org/142133539
IDR: 142133539 | УДК: 631.461.52:581.557.22:581.148.5:577.21
Symbiotic nodule senescence in legumes: molecular-genetic and cellular aspects (review)
Senescence is the natural stage in development of symbiotic nodule. As a result of senescence, reutilization of different nutrients from nodule to the other plant organs occurs. Generally senescence in legumes is triggered after flowering finishing, although the first traits of senescence can be observed very early during nodule development. A delay of the triggering of senescence program will allow to prolong the active nitrogen-fixation period and therefore to increase the amount of symbiotrophic nitrogen in plants and, finally, to elevate legume productivity. That is why no wonder that in the recent years the senescence of nitrogen-fixing nodules is actively studied. In this review the main developmental stages of nitrogen-fixing symbiotic nodule of legumes, particularities of symbiotic nodule development of determinate and indeterminate types are considered. In legumes with indeterminate nodules, the symbiosomes are not long leaving as the infected tissues are permanently renewing due to apical meristem. There are two subsequent stages identified in an indeterminate nodule senescent. First a bacteroid degradation and the death of some infected cells occur, and then both symbiosomes and all infected cells are destroyed. In determinate nodules, the senescence initiated in the central part of a nodule, then extends to the peripheral zone. In this review morphological characters of nodule senescence at ulatrstructural level are analysed. The role of cysteine and threonine proteases is discussed. Reutilization of nitrogen and other products of protein degradation are probably the most important during senescence. There are the evidences that in the root nodules of legumes the cysteine proteases are involved into nodular functions, adaptation of the host plant cells to physiological stresses, and the nodule senescence control. By a large-scale analysis of nodule transcriptome of Medicago truncatula Gaertn. several gene groups expressed at successive stages of the senescence of indeterminate nodule are revealed. In this review the role of phytohormones, such as ethylene, abscisic acid, jasmonic acid, gibberellins and nitrogen monooxide in senescence of symbiotic nodule is considered. Nevertheless, until recent days our knowledge about hormonal control of a nodule senescence is still incomplete. The oxidative stress, accompanying the process of nodule senescence is discussed. On the nodules aging, the concentrations of peroxides, protein carbonils, modified DNA nucleotides and catalytic Fe increase. Iron activates lipids peroxidation in a peribacteroid space, resulting in degradation of the peribacteroid membrane in senescent nodules. The concentrations of oxidized glutathione and homoglutathion rise significantly during the nodule development, and the reduced forms decrease under senescence, indicating an oxidative stress in the senescing nodules. In this review the role of genes, encoding proteins involved in transport of wide-range of molecules, and genes, whose products are involved in regulatory and signal functions in cell; differences between stress-induced senescence and natural senescence are considered. Using model legumes, Lotus japonicus (Regel) K. Larsen and M. truncatula, several genes were cloned the mutations of which caused early senescence. It is emphasized that these genes encode different proteins involved into functions of a symbiotic nodule. Until now, two transcription factors in M. truncatula are described, which are involved into nodule senescence. An induced senescence is more rapid, comparing to natural senescence, and manifests the signs of an oxidative stress and programmed cell death.
Текст обзорной статьи Старение симбиотического клубенька у бобовых растений: молекулярно-генетические и клеточные аспекты (обзор)
Работа финансово поддержана Министерством образования и науки (Государственный контракт № 16.552.11.7085, соглашение 8109), грантом Президента РФ (НШ-337.2012.4), РФФИ (14-04-00383).
Старение — естественный этап в развитии симбиотического клубенька. В результате процесса старения происходит реутилизация различных питательных веществ из клубенька в другие органы растения. В большинстве случаев этот механизм запускается после окончания цветения бобовых, хотя первые признаки старения могут наблюдаться на очень ранних стадиях развития клубенька.
Развитие симбиотического клубенька. Это развитие начинается в результате сигнального взаимодействия между бобовым растением и клубеньковыми бактериями, обобщенно называемыми ризобия-ми. Различные аспекты процесса развития симбиотического клубенька неоднократно рассмотрены в многочисленных обзорах (1-5). Кратко его можно описать следующим образом. Бобовые растения выделяют флавоноиды, которые индуцируют у ризобий синтез липохитоолигосахаридов, называемых Nod-факторами. Nod-факторы воспринимаются специфичными рецепторами растений, и активируются механизмы сигнальной трансдукции, в результате чего, с одной стороны, происходит инфицирование хозяина ризобиями, а с другой — индуцируется формирование примордия симбиотического клубенька (1, 5). Инфекционный процесс начинается с деформации и скручиваний корневых волосков с образованием ими инфекционного кармана, в котором оказываются заключенными ризобии (2, 5). Отсюда они продвигаются вглубь корневого волоска, а впоследствии и клеток корня благодаря специальному каналу, формируемому растением и называемому инфекционной нитью (3-5). Одновременно в коре корня и перицикле индуцируются клеточные деления, приводящие к образованию клубенькового примордия. Инфекционная нить достигает примордия, где появляются специальные выросты, окруженные лишь плазматической мембраной и лишенные стенки, ограничивающей в норме инфекционную нить (3, 4). Из этих выростов, называемых инфекционными каплями, посредством процесса, напоминающего эндоцитоз, происходит высвобождение ризобий в цитоплазму растительной клетки (4). При этом бактерии остаются окруженными симбиосомной (перибактероидной) мембраной растительного происхождения, но с включениями бактериальных белков (1, 4). В дальнейшем происходит дифференцировка бактерий в специализированную форму — в бактероиды. Бактероид, ограниченный симбиосомной мембраной, называется симбиосомой, которая представляет собой основную азотфиксирующую единицу (1, 4, 5). У бобовых растений из умеренных широт (горох Pisum sativum L., люцерна Medicago sativa L . ) формируется меристема, которая сохраняет активность на протяжении всей жизни клубенька (такие клубеньки называются недетерминированными). В результате постоянной активности меристемы в клубеньке можно выделить следующие зоны: I — собственно меристему; II, III и IV — соответственно зоны инфекции, азотфиксации и старения (рис. 1, А, В). Для недетерминированных клубеньков характерна вытянутая удлиненная форма. В клубеньках бобовых из южной зоны (соя Glycine max L., фасоль Phaseolus vulgaris L.) меристема функционирует непродолжительное время, поэтому зональность клубенька отсутствует, и они имеют сферическую форму (такие клубеньки называются детерминированными) (1, 4, 5).
Инфицированные растительные клетки, находясь в зоне азотфиксации, функционируют на протяжении ограниченного периода времени, после которого наступает старение, представляющее собой организованный процесс клеточной гибели.
У бобовых растений с недетерминированными клубеньками время жизни симбиосом невелико, потому что центральные инфицированные
Рис. 1. Симбиотические азотфиксирующие клубеньки у гороха Pisum sativum L. родительской линии SGE (А, Б, возраст соответственно 4 и 6 нед) и полученного на ее основе симбиотического мутанта SGEFix - -8 ( sym25) , характеризующегося признаками раннего старения симбиотических клубеньков (В, Г, возраст соответственно 4 и 6 нед) : I — зона меристемы, II — зона инфекции, III — зона азотфиксации, IV — зона старения (масштабная линейка — 200 мкм).
ткани постоянно обновляются за счет апикальной меристемы. Так, первые признаки старения симбиосом у гороха и люцерны наблюдаются уже на 14-е сут после инокуляции (6), хотя активный процесс старения запускается значительно позже — после окончания цветения. У видов бобовых растений с детерминированными клубеньками начало старения также совпадает с концом цветения—началом плодоношения, что было показано на G. max (7) и Vigna mungo (L.) Hepper (8). Часто старение клубенька ускоряется, если симбиоз неэффективен из-за генетических мутаций в растениях (9, 10) или в бактериях (11). Старение может также индуцироваться различными абиотическими стрессами (12).
Морфологические признаки старения. Визуально старение в клубеньках проявляется через изменение цвета азотфиксирующей зоны от розового, связанного с функционированием леггемоглобина, до зеленого, что обусловлено разрушением гемовой группы этого белка и образованием биливердина (13).
При старении клетки обоих симбиотических партнеров разрушаются (см. рис. 1, Б - Г, 2, А, Б). Однако в случае недетерминированных клубеньков зона азотфиксации постоянно обновляется за счет клеток, переходящих в нее из меристемы, хотя с возрастом клубенька зона старения увеличивается. На ультраструктурном уровне обнаруживаются изменения в симбиосомах и клеточных органеллах: у цитоплазмы уменьшается электронная плотность, наблюдаются везикулы и остатки мембран как результат разрушения клетки хозяина и симбиосом (14, 15).
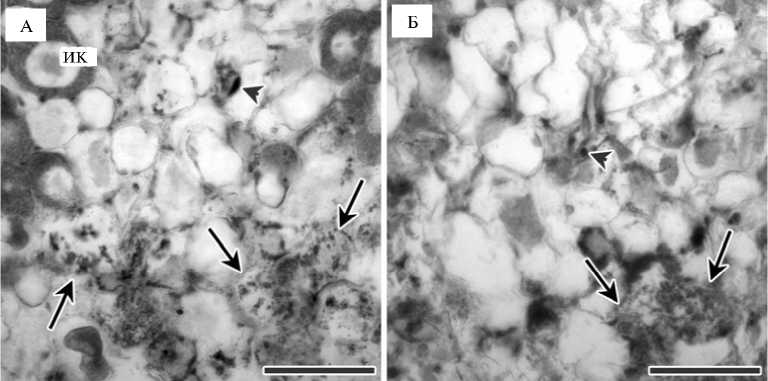
Рис. 2. Зона старения симбиотического азотфиксирующего клубенька у гороха Pisum sativum L. родительской линии SGE (А) и полученного на ее основе симбиотического мутанта SGEFix - -8 ( sym25) , характеризующегося признаками раннего старения симбиотических клубеньков ( Б) : ИК — инфицированная клетка; малая стрелка — ранняя деградация бактероидов и клеток хозяина (клетки II типа), большая стрелка — глубокая деградация клеток хозяина и бактероидов (клетки III типа) (масштабная линейка — 100 мкм).
В детерминированных клубеньках старение инициируется в центральной зоне клубенька и распространяется к периферии (16).
На молодых клубеньках у модельного бобового растения Medicago truncatula Gaertn. было показано (15), что в центре зоны азотфиксации имеется несколько стареющих инфицированных клеток так называемого I типа. У них отмечаются признаки деградации симбиосом, которая сопровождается увеличением везикулярной активности и возрастанием количества шероховатого эндоплазматического ретикулума. Растительные клетки при этом остаются интактными (без видимых признаков старения). В той же зоне наблюдаются клетки с более выраженными симптомами старения (клетки II типа), характеризующиеся нарушением целостности клеточной стенки. В более старых клубеньках наряду со стареющими клетками I и II типа у основания клубенька присутствуют многочислен- ные клетки III типа с полностью разрушенными симбиосомами и признаками гибели клетки хозяина (отслаивание плазматической мембраны от клеточной стенки).
Основываясь на ультраструктурных исследованиях, выделяют две последовательные стадии в процессе старения недетерминированного клубенька: первая, более ранняя, когда происходит деградация бактероидов и гибель нескольких растительных клеток, и более поздняя, на протяжении которой полностью разрушаются и симбиосомы, и инфицированные клетки (12, 15).
У гороха в клубеньках неэффективных мутантов с ранним старением в стареющих клетках наблюдались лизосомоподобные компартменты, содержащие деградирующие бактероиды (9).
Форма зоны старения в зрелых недетерминированных клубеньках M . truncatula характеризуется коническим фронтом, который направлен в сторону роста клубенька. Инфицированные клетки разрушаются раньше неинфицированных, которые важны для транспорта веществ к проводящим тканям. Коническая организация фронта старения, вероятно, способствует оптимальной ремобилизации продуктов распада, потому что в этом случае последними погибают клетки, локализованные на периферии около проводящих пучков. Таким образом, центральное расположение места инициации старения обусловлено наибольшей дистанцией от периферической проводящей системы и окружающей клубенек среды, что, возможно, связано с ролью градиента концентрации кислорода или молекул, поступающих от проводящих пучков (12).
В клубеньках люцерны, начиная с 7-й нед, помимо IV зоны, была выделена V зона, расположенная проксимально по отношению к зоне старения (14). В пределах V зоны наблюдается новый этап выхода ризобий из оставшихся инфекционных нитей, приводящий к возобновлению инвазии в отношении старых растительных клеток. Эта зона представляет собой экологическую нишу, где внутриклеточные ризобии используют взаимодействие исключительно для своей пользы как сапрофитные партнеры (14).
Протеазы. Функционально старение представляет собой процесс ремобилизации веществ, накопленных в старых тканях и транспортируемых в другие части растения, где они могут быть использованы для образования новых органов. Деградация белка, которая позволяет повторно утилизировать азот и другие вещества, — вероятно, наиболее важный из процессов, происходящих на протяжении старения. Поэтому неудивительно, что многие гены, для которых описана позитивная регуляция при старении, — это гены протеаз (17).
В процессе старения у растений задействованы несколько протеаз. В проведенных ранее исследованиях (12, 15, 18) было установлено, что в старении клубенька участвуют в основном цистеиновые и треониновые протеазы.
Существуют доказательства, что в корневых клубеньках бобовых растений цистеиновые протеазы вовлекаются в функционирование клубенька, в адаптацию клеток хозяина к физиологическим стрессам и в контроль старения клубенька. Протеазы и их ингибиторы были идентифицированы в цитоплазме инфицированных клеток клубенька (7, 19). Цистеиновые протеазы с кислым оптимумом рН были описаны в клубеньках фасоли ( P. vulgaris ) (19); их активность увеличивалась к началу старения. Подобные исследования проводились на сое ( G. max ) (7), черном нуте ( V. mungo ) (8) и люцерне ( M. sativa ) (10).
На растениях гороха (P. sativum) изучали характер экспрессии генов цистеиновых протеаз 1 (PsCypl) и 15а (PsCyp15a) (18). Кроме эффективно- го штамма ризобий (Rhizobium leguminosarum bv. viciae, 3841), в работе использовался дефектный по синтезу липополисахаридов штамм В661, при инокуляции которым формировались рано стареющие клубеньки. Экспрессия генов обеих протеаз (PsCypl и PsCyp15a) повышалась с возрастом клубенька (18).
Позже при широкомасштабном анализе транскриптома клубеньков люцерны ( M . truncatula ) выявили несколько групп генов, активность которых характерна для последовательных стадий старения недетерминированного клубенька. Усиление экспрессии генов, относящихся к семейству протеаз, наблюдалось на поздних этапах старения при значительной степени деградации обоих партнеров симбиоза (15).
Среди выявленных молекулярных маркеров поздних стадий старения был отмечен ген вакуолярного процессирующего энзима (VPE, семейство С13), тоже входящего в группу цистеиновых протеаз и ответственного за созревание вакуолярных белков, а также гены, кодирующие треониновые протеазы, которые участвуют в последовательных процессах, описанных как F-box-специфичный убиквитин/26S протеосомный путь (15).
Гopмo^ы . Развитие бобово-ризобиального симбиоза также подвергается гормональной регуляции со стороны растения-хозяина. Из фитогормонов в старении симбиотического азотфиксирующего клубенька участвуют этилен, абсцизовая и жасмоновая кислоты, гиббереллины и оксид азота NO. Тем не менее, следует отметить, что до настоящего времени представления о гормональной регуляции этого процесса остаются достаточно фрагментарными.
Этилен играет роль одного из активаторов старения симбиотического клубенька бобовых растений, что иллюстрируется позитивной регуляцией в стареющих клубеньках соответствующих факторов транскрипции (ERF) и генов биосинтеза этилена, таких как S-аденозилметионин (SAM)-синтетаза и 1-аминоциклопропан-1-карбоксилат-оксидаза (15).
Индукция липоксигеназных генов свидетельствует о вовлечении жасмоновой кислоты в различные стадии старения симбиотического клубенька. Окисление полиненасыщенных жирных кислот при участии липоксигеназных ферментов представляет собой первый этап биосинтеза оксилипинов, таких как жасмонаты (15, 20).
На протяжении естественного старения клубенька наблюдается уменьшение аскорбат-глутатион антиоксидантного пула в сочетании со снижением углерод-азотного соотношения в тканях клубенька. Эти изменения могут восприниматься и передаваться при посредстве сигнальных механизмов, ассоциированных с абсцизовой кислотой (АБК), мобилизующей протеолитическую активность в стареющем симбиотическом клубеньке (15, 21). АБК может участвовать в индукции фермента, повышающего акцепторную способность, и таким образом содействовать раннему развитию отдельных клубеньков (22). Однако повышение скорости синтеза АБК при старении может стать причиной гибели клубенька. Содержание АБК в клубеньках гороха ( P. sativum ) довольно высокое на протяжении первых 2 нед, но затем этот показатель выходит на плато, снова увеличиваясь на поздних стадиях развития (23).
Гибберелловая кислота может подавлять старение клубенька, что иллюстрируется активацией генов, кодирующих гиббереллин-2-оксидазу, превращающую активную форму фитогормона в неактивную (15, 24).
Недавно с использованием модельного бобового растения M. truncatula выявили регуляцию старения симбиотического клубенька монооксидом азота (25). Было показано, что в контроле за количеством NO в клубеньке большую роль играет бактериальный флавогемоглобин, поскольку мутантный штамм hmp, неспособный к его биосинтезу, проявлял признаки более раннего старения, а также характеризовался повышенным содержанием NO в тканях клубеньков, в то время как штамм со сверхэкспрессией гена hmp++, напротив, демонстрировал признаки замедленного старения и пониженное содержание NO в клубеньках. Эндогенное добавление монооксида азота также приводило к преждевременному старению клубенька (25).
Окислительный стресс при старении. Условия функционирования бобово-ризобиального симбиоза изначально способствуют формированию активных форм кислорода (АФК). Показано, что оксилег-гемоглобин подвержен медленному автоокислению с сопутствующим образованием супероксид-аниона О2 - , который диспропорционирует с образованием перекиси водорода Н2О2 (26). Кроме того, реакция леггемогло-бина с Н2О2 приводит к образованию окисленных форм (например, белков с феррильным железом и белковых радикалов) (27). Перекись водорода также может вызывать разрушение белков с высвобождением каталитического железа, то есть железа в молекулярной форме, которое способно активировать перекисное окисление липидов и образование гидроксильных радикалов (28). АФК выделяются и при восстановительных процессах в условиях, необходимых для азотфиксации, и при действии некоторых белков, включая ферредоксин, уриказу и гидрогеназу (29).
Однако в клубеньках содержится также большое количество антиоксидантов: аскорбата, глутатиона, супероксиддисмутаз, каталаз и ферментов аскорбат-глутатионового цикла, пероксиредоксина (30). Антиоксиданты поддерживают АФК в постоянных низких концентрациях: свободные радикалы были детектированы в клубеньках (31, 32), и их концентрация повышалась с возрастом клубенька. Важную роль играют различные антиоксидантные системы ризобий. Более того, экстракты из стареющих клубеньков вызывали перекисное окисление липидов из мембран растительных клеток и липидов перибактероидных мембран (26). Цитохимический анализ с использованием хлорида церия выявил присутствие электронно-плотных преципитатов пергидроксида церия в стенках стареющих инфицированных клеток и вокруг перибактероидных и бактероидных мембран, что доказывает участие перекиси водорода в процессе старения симбиотического клубенька (33, 34).
Исследование, проведенное на 2- и 10-недельных клубеньках растений сои ( G. max ), продемонстрировало, что окислительный стресс сопровождает процесс старения симбиотического клубенька бобовых растений. С возрастом клубенька содержание пероксидов, карбонилов белков и модифицированных оснований ДНК увеличивалось, каталитического железа — тоже значительно возрастало. Железо, содержащееся в перибактероидном пространстве, усиливало перекисное окисление липидов, что могло способствовать деградации перибактероидной мембраны в стареющих клубеньках. Количество окисленных форм глутатиона и гомоглутатиона значительно повышалось по мере развития клубенька, восстановленных — снижалось при старении, что свидетельствует о развитии окислительного стресса в стареющих клубеньках. В первые сутки развития клубенька уже наблюдались значительные разрушение ДНК и белков, демонстрируя тем самым, что оксислитель-ный стресс может происходить в период становления симбиоза (35).
Диффузионный барьер, который в функционирующих клубеньках создает градиент кислорода, в стареющих, вероятно, ниже, что может способствовать увеличению потока кислорода к центральным тканям и уско- рению продукции АФК (35).
Генетический контроль. В экспериментах по мутагенезу, проводившихся в течение многих лет с использованием различных бобовых растений, были выявлены мутанты, характеризующиеся неспособностью к фиксации атмосферного азота (Fix - фенотип). Изучение гистологической и ультраструктурной организации полученных Fix - мутантов показало, что их значительная часть имеет фенотип Nop - (от англ. nodule persistence), то есть они неспособны поддерживать структурно-функциональную стабильность клубеньков. Как результат, в клубеньках у таких мутантов запускается процесс преждевременного (раннего) старения (см. рис. 1, В, Г; 2, Б). Например, у гороха было показано, что раннее старение свойственно для мутантов по генам ^ym13 , sym25 , sym26 , sym27 и sym42 (36). По всей видимости, проявление раннего старения можно рассматривать как типичную реакцию со стороны растения-хозяина на формирование неэффективного симбиоза.
С использованием модельных бобовых Lotus japonicus (Regel) K. Larsen и M . truncatula были клонированы некоторые гены, мутации по которым приводят к инициации раннего старения. Важно отметить, что эти гены кодируют самые разные белки, участвующие в функционировании симбиотического клубенька.
Первым был клонирован ген LjSSTl , кодирующий расположенный на симбиосомной мембране клубенек-специфичный переносчик сульфатов из цитоплазмы растительной клетки к бактероидам (37). У мутантов по этому гену формируются рано стареющие клубеньки, а в инфицированных клетках появляются литические вакуоли (37).
Ген LJIGN1 кодирует белок с анкириновыми повторами и трансмембранными участками, который необходим для дифференциации и поддержания функционирования бактероидов, хотя его точная роль остается неизвестной (38). Мутант по указанному гену характеризуется агрегацией бактероидов внутри инфицированных клеток с дезинтегрированными внутриклеточными структурами, причем преждевременное старение у него наступает значительно раньше, чем у других Fix - мутантов L . japonicus (38).
Ген LjSENl кодирует интегральный мембранный белок, гомологичный nodulin-21 у G. max, Fe/Mn переносчику CCC1 у Saccharomyces cer-evisiae и переносчику железа VIT1 у Arabidopsis thaliana L. LjSEN1 экспрессируется только в инфицированных клетках клубенька (39). Мутанты по этому гену демонстрируют фенотип раннего старения (40).
У M . truncatula был клонирован ген MtDNF2 , который в результате альтернативного сплайсинга может транскрибироваться с образованием пяти транскриптов. Предоминантный транскрипт кодирует PI-PLC-like protein (белок, подобный фосфатидилинозитол-фосфолипазе C). Предполагается, что он может связываться с фосфатидилинозитолом или фосфорилированным фосфатидилинозитолом, предотвращая их расщепление на фосфат инозитола и диацилглицерин, которые служат вторичными мессенджерами или предшественниками вторичных мессенджеров, запускающих защитные реакции. Мутанты по описанному гену образуют клубеньки с признаками раннего старения (41).
Недавно у M. truncatula описан мутант esn1 с признаками раннего старения, при этом у него наблюдалась дифференциация бактероидов и экспрессия гена nifH , кодирующего один из компонентов нитрогеназы, и LHG , кодирующего леггемоглобин (42).
Следует отметить, что к настоящему времени не получены индуцированные мутанты с фенотипом замедленного старения. В то же время соз-10
даны трансгенные растения люцерны M . truncatula с заблокированной экспрессией гена цистеиновой протеазы 15а ( Сур15а ), которые проявляют фенотип отсроченного старения симбиотических клубеньков (43).
Анализ транскриптома стал еще одним подходом для выявления генов, участвующих в реализации программы старения симбиотического клубенька. Так, в 2006 году у люцерны ( M. truncatula ) разного возраста проанализировали транскриптом клубеньков с целью выявления ассоциированных с их старением генов (15). С помощью кластерного анализа обнаружили три группы генов, соответствующие последовательным стадиям старения клубенька. К первому кластеру были отнесены регуляторные гены, активные при инициации старения: гены, кодирующие факторы транскрипции семейства APETALA/Ethylene Response Factor (AP2/ERF), которые задействованы также в обеспечении устойчивости к болезням и в стрессовых ответах; гомолог гена, кодирующего транскрипционный фактор, нечувствительный к абсцизовой кислоте и обладающий ДНК-свя-зывающими доменами для AP2/ERF и В3; гены протеинкиназ и МАР-киназ, продукты которых участвуют в трансдукции сигналов, ассоциированных со стрессом и реакцией на условия окружающей среды; DEAD-box генов РНК-хеликазы, продукты которых участвуют в экспорте мРНК при стрессе и развитии растения (15). Гены, объединенные во второй и третий кластеры, вовлечены в процессы соответственно на I стадии старения, сопровождающейся разрушением бактероидов, и на более поздней II стадии, когда клетки хозяина подвергаются деградации. Оба обозначенных кластера включают гены, выполняющие потенциально регуляторные и сигнальные функции. Кроме того, механизм деградации белков, расщепления нуклеиновых кислот, мембранных липидов и сахаров регулируется на уровне транскрипции. Усиление экспрессии генов катаболизма проявляется в разрушении симбиосом на ранней стадии старения и клеток хозяина — на более поздней стадии. В то же время индукция генов, которые кодируют белки, вовлеченные в транспорт широкого круга молекул (АТФ-связывающие белки и специфические переносчики фосфатов, аминокислот, ионов металлов), показывает, что деградация макромолекул и функции мобилизации тесно связаны и при старении клубенька происходит рециклизация метаболитов. Эти катаболические события, связанные с транспортными процессами, превращают клубенек из реципиента углерода в донора питательных веществ (15).
К настоящему времени у M. truncatula выявлены два транскрипционных фактора, задействованных в процессе старения симбиотического клубенька. Так, наблюдалось усиление экспрессии гена MtATB2 , кодирующего фактор транскрипции bZIP-типа. Транскрипты обнаруживались в апикальной части и проводящих тканях клубенька. Транскрипция этого гена репрессируется сахарозой, а сам белок MtАТВ2 участвует в регуляции активности генов аминокислотного метаболизма (44). Другой транскрипционный фактор — MtNAC969 участвует в репрессии генов, которые активируются в корнях при солевом стрессе, причем оказалось, что РНК-интерференция транскриптов этого гена приводит к проявлению признаков преждевременного старения у симбиотического клубенька (45).
В дополнение к транскрипционному контролю в случае определенных ответов клубенька, ассоциированных со старением, осуществляется регуляция на уровне трансляции, так как некоторые из изученных генов (например, ген, кодирующий 40S ribosomal protein S8) кодируют рибосомальные белки, факторы элонгации и другие протеины, вовлеченные в регуляцию трансляции (15).
Стресс-индуцированное и естественное старение.
В результате стрессового воздействия может инициироваться преждевременное старение клубенька. При этом индуцированное старение происходит быстрее естественного и имеет признаки окислительного стресса и запрограммированной клеточной гибели (15).
При сравнительном анализе естественного и стресс-индуцирован-ного старения клубеньков у M . truncatula на ультраструктурном уровне выявили особенности, характерные для стресс-индуцированного процесса: содержимое бактероидов уплотненное, перибактероидное пространство увеличено, часто наблюдается слияние различных симбиосом. Однако пери-бактероидная мембрана остается интактной даже тогда, когда содержимое большинства бактероидов исчезает, что представляет собой явное отличие от естественного старения, когда симбиосомы разрушаются полностью. Транспорт везикул при этом тоже менее выражен. После вакуолярного разложения цитоплазма полностью деградирует, митохондрии исчезают и видимыми остаются лишь остатки перибактероидных мембран и бактероидов. Сапрофитные бактерии из инфекционных нитей заселяют старые клетки уже на ранней стадии деградации (12).
Старение, индуцированное темнотой, происходит быстрее и сопровождается разрушением содержимого симбиосом в отсутствие клеточных сигналов для ремобилизации веществ и быстрой колонизацией сапрофитными бактериями (12).
При сравнении естественного и индуцированного темнотой старения клубенька были отмечены различия не только в морфологии, но и в изменении экспрессии генов. На уровне транскрипции из генов, активируемых при естественном старении клубенька, 50 % не участвуют в стресс-индуцированном старении. К ним относятся гены, ответственные за регуляцию и транспорт, деградацию мембран и белков и стрессоустойчивость (кодирующие синтаксин и две фосфотидил-4-фосфат-5-киназы, обеспечивающие специфический транспорт везикул) (46). В отношении некоторых других генов отмечалась лишь временная положительная регуляция при индуцированном темнотой старении клубеньков. Гены, которые, скорее всего, не вовлечены в такое индуцированное старение, кодируют многие функции, связанные с деградацией белков при посредстве протеосом, и некоторые цистеиновые протеазы, что, видимо, связано с упрощенной организацией процесса деградации при стресс-индуцированном старении по сравнению с естественным старением. В недавнем исследовании показано, что индуцирование старения переносом растений в темноту сопровождается резким увеличением pH перибактероидного пространства (47).
Итак, в настоящее время молекулярно-генетические и клеточные механизмы, лежащие в основе реализации программы старения симбиотического клубенька, активно изучаются. Понимание этих механизмов позволит приступить к практическому созданию сортов бобовых растений с продолжительной фиксацией азота, что может положительно сказаться на насыщении почвы биологическим азотом и урожайности возделываемых культур на фоне снижения доз химических удобрений.
