Стерильность при межподвидовой гибридизации риса Oryza sativa L. в связи с поиском генов широкой совместимости и отнесением образцов к подвидам Indica и Japonica

Автор: Харитонов Е.М., Гончарова Ю.К.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Совместимость при гибридизации
Статья в выпуске: 5 т.48, 2013 года.
Бесплатный доступ
Между подвидами риса ( Oryza sativa L.) indica и japonica существует ряд переходных форм как по фенотипу, так и по физиологическим признакам, обусловливающими особенности развития и адаптации к стрессам. Гетерозисный эффект у межподвидовых гибридов ( japonica / indica ) может составлять 100 % и выше. Использовать сверхвысокий гетерозис таких гибридов можно, если вовлекать в гибридизацию образцы с генами широкой совместимости (WC), которые обеспечивают озерненность гибридов более 70 %. Мы провели гибридизацию между образцами отечественной и зарубежной селекции, относящимися к подвидам indica и japonica. Анализ озерненности полученных межподвидовых гибридов не позволил выделить среди образцов российской селекции сорта с генами широкой совместимости. Для корректного отнесения сортов риса к подвидам выполнено молекулярное маркирование с использованием 11 полиморфных SSR-маркеров, типичных для подвидов. Обсуждаются вопросы получения фертильных межподвидовых гибридов риса и перечень признаков, используемых для разделения образцов на подвиды. Установлена необходимость включения дополнительных характеристик в список признаков, на основании которых должно проводиться разделение отечественных образцов на подвиды indica и japonica.
Рис, гетерозис, гибриды, гены широкой совместимости, подвиды риса японский и индийский
Короткий адрес: https://sciup.org/142133444
IDR: 142133444 | УДК: 633.18:631.522/.524:575.222.7
Sterility of intersubspecific hybrids of Oryza sativa L. in connection with search of genes of wide compatibility and correct reference of samples to subspecies Indica and Japonica
In rice ( Oryza sativa L.) between indica and japonica subspecies a number of transitional forms both on phenotype and on physiological determinants present, which have the characteristic features in their development and adaptation to stress. The heterosis effect in intersubspecific hybrids ( japonica/indica ) can be of 100 % and more. The superhigh heterosis can be used during intersubspecific hybridization on conditions that the genes of wide compatibility (WC), which ensure a grain content more of 70 %, will be involve in breeding. The authors made the crossings between the samples of domestic and foreign breeding from japonica subspecies. The analysis of grain formation in the hybrid combinations did not allow to reveal WC genes in studied varieties of Russian selection defining high fertility of hybrids. The molecular scanning of hybrids and their parental forms was made with the use of 11 polymorphous SSR-markers, typical for subspecies. A possibility to obtain the fertile intersubspecific hybrids of rice and the list of trait used for differentiating samples to subspecies japonica and indica are discussed. The obtained results show that it is necessary to include additional characteristics into the list of trait for the taxonomic attributing samples to indica and japonica subspecies.
Текст научной статьи Стерильность при межподвидовой гибридизации риса Oryza sativa L. в связи с поиском генов широкой совместимости и отнесением образцов к подвидам Indica и Japonica
Существуют две гипотезы происхождения подвидов. Согласно первой, японский подвид произошел из образцов индийского подвида при их адаптации к условиям более северных регионов (4). Другая теория говорит о вероятной параллельной доместификации подвидов, происходящих от различных диких родственников (5). Значительные генетические различия между подвидами приводят к высокому гетерозису при их гибридизации, что позволяет надеяться на возможность увеличения потенциала продуктивности культуры за счет межподвидовой гибридизации (6, 7). Исследования показали, что к дифференциации подвидов привела комбинация таких генетических событий, как вставки, делеции, замены нуклеотидов. Наибольшее число различий между подвидами, связанных с изменением нуклеотидных последовательностей, зафиксированы на 11-й и 12-й хромосомах (8-10).
На территории Российской Федерации в основном используются сорта риса подвида japonica. Низкая генетическая гетерогенность сортов этого подвида приводит к невысокому гетерозису при их гибридизации (11, 12). У межподвидовых гибридов (japonica/indica) гетерозисный эффект может составлять 100 % и более. Однако вследствие высокой пусто-зерности межподвидовые гибриды риса до сих пор промышленно не производятся, гибриды подвида japonica также не получили широкого распространения (посевные площади не превышают 5 %) из-за незначительного гетерозисного эффекта (13). В то же время посевные площади под гибридами индийского подвида в Китае в 4 раза больше занимаемых сортами того же подвида (14). Использовать сверхвысокий гетерозис межподвидовых гибридов, однако, можно, если вовлекать в селекцию сорта с генами широкой совместимости (WC), которые обеспечивают озерненность гибридов более 70 % даже при межподвидовой гибридизации (15). Подвид japonica несет аллель S5J, indica — S5, образцы с геном широкой совместимости — аллель S5n. Гетерозиготные формы S5J/S5 стерильны, образцы, несущие гены широкой совместимости, фертильны (S5J/S5n, S5i/S5n) (16, 17). Внедрение генов широкой совместимости в формы с высокой комбинационной способностью даст возможность получить фертильные межподвидовые гибриды с величиной гетерозиса до 100 % (18). Кроме того, межподвидовая гибридизация позволит расширить норму реакции создаваемого образца за счет использования генетического потенциала обоих подвидов и, следовательно, обеспечит большую стабильность получаемых урожаев риса. Гены, обусловливающие широкую совместимость, картированы, определен список микросателлитных SSR-маркеров (simple sequence repeat), позволяющих осуществлять интрогрессию этих генов в перспективные сорта (19-22). Однако выявленные доноры генов широкой совместимости в большинстве случаев представлены образцами из тропического региона (например, Дулар, Нагина 22) и несут ряд признаков, характерных для диких видов (осыпаемость, низкая продуктивность, склонность к полеганию, высокорослость и т.д.) В то же время имеются сообщения о выявлении генов WC у современных сортов (23).
Цель настоящей работы заключалась в уточнении таксономической принадлежности изучаемых образцов риса к двум подвидам, выявлении у них генов широкой совместимости и сравнении свойств получаемых гибридов и их родителей в связи с перспективой создания фертильных межподвидовых гетерозисных гибридов.
Методика . Мы провели гибридизацию между образцами риса Ory-za sativa L. отечественной селекции, относящимися к подвиду japonica (Хазар, Регул, Лиман, Славянец, Дружный, ВНИИР 7679, ВНИИР 7653 и т.д.), а также образцами подвида indica отечественной и зарубежной селекции. Оценку гибридов проводили при выращивании в сосудах на вегетационной площадке (10 растений на сосуд) на оптимальном фоне минерального питания N120P 6 0K 6 0. В выборках (20 растений на сорт и гибридную комбинацию) в фазу созревания проводили биометрический анализ. Родительские формы, гибридные комбинации которых показывали стерильность более 30 %, относили к не обладающим генами широкой совместимости. Пус-тозерность определяли как процентное отношение числа стерильных колосков к их общему числу.
Молекулярный анализ выполняли на 20 растениях каждого образца с использованием SSR-маркеров, полиморфных для типичных образцов подвидов. При молекулярном маркировании использовали сорта Ханкай-ский, Садко, Приморский, Лиман, Гарант, Павловский, Рапан, Новатор,
Серпантин, Боярин, Регул, Янтарь, Жемчуг, Лидер, Хазар, Аметист, Нарцисс, Дружный, Спринт, Виола, Дальневосточный, Фонтан, Касун, Юпитер, Атлант, Курчанка, Факел, Снежинка, Шарм, Анаит, Флагман, Изумруд, IR 66, NSIC RC 158, IR 68897B (последние три — типичные образцы подвида indica селекции IRRI (IRRI — International Rice Research Institute, Филиппины). Среди сортов российской селекции подвид indica был представлен сортами Серпантин, Нарцисс Снежинка, Шарм, Изумруд.
ДНК выделяли из этиолированных проростков и листьев с помощью STAB-метода в различных модификациях, постановку полимеразной цепной реакции (ПЦР) и визуализацию продуктов амплификации проводили по методике IRRI (24).
Статистическую обработку данных осуществляли в программе Sta-tistica v. 6.0.
1. Озерненность гибридов первого поколения при скрещивании сортов риса ( Oryza sa-tiva L.) отечественной селекции, относимых к подвидам japonica и indica по индексу зерновки (вегетационная площадка, 2008-2009 годы)
|
Материнская форма х опылитель | |
Пустозерность, % |
|
japonica х indica : |
|
|
Хазар х Снежинка |
5,76 |
|
Хазар х Изумруд |
18,13 |
|
Диана х Майя |
14,04 |
|
Диана х Снежинка |
9,09 |
|
Дружный х Снежинка |
20,70 |
|
indica х japonica : |
|
|
Снежинка х Хазар |
12,57 |
|
Снежинка х Регул |
9,90 |
|
Майя х Хазар |
6,75 |
|
Изумруд х Хазар |
14,25 |
|
japonica х japonica : |
|
|
Хазар х Лиман |
3,60 |
|
Регул х Славянец |
30,40 |
|
Дружный х Лиман |
20,00 |
|
Диана х Лиман |
19,40 |
|
ВНИИР 7653 х Дружный |
17,20 |
Резуёьтаты . Мы провели поиск генов широкой совместимости (WC) среди коллекционных образцов риса. Родительские формы, гибридные комбинации которых при межподвидовой гибридизации показывали пустозерность более 30 %, относили к не обладающим генами широкой совместимости (табл. 1).
Сравнение озерненно-сти гибридов от скрещивания образцов российской селекции, которые ранее были отнесены к разным подвидам на основании различий по индексу зерновки (отношение ее длины к ширине), показало низкую пустозерность, не от личающуюся достоверно от таковой у гибридов одного подвида (см. табл. 1.). Удивило то, что при гибридизации российских образцов одного
подвида пустозерность во многих случаях превышала таковую у межподвидовых гибридов. Вывод из этого может быть только один: или многие российские сорта несут гены широкой совместимости, или принятая в России классификация не позволяет корректно отнести формы к подвидам. Анализ происхождения селекционного материала выявил достаточно близкое родство образцов отечественной селекции, в том числе номинируемых как разные подвиды. Ранее образцы риса можно было разделять на подвиды согласно происхождению и форме зерновки, однако широкая гибридизация подвидов в ряде исследовательских центров привела к появлению большого числа форм, которые за счет рекомбинации несут гены, характерные для разных подвидов (25-27).
По наблюдениям многих ученых, в настоящее время невозможно на основании различий по одному признаку или гену достоверно дискриминировать подвиды indica и japonica (28), их разделение только по индексу зерновки более чем в 39 % случаев приводит к неправильной классификации. Аттрибутирование по длине опушения цветковых чешуй также дает ошибку в 20-40 % вариантов. Для более точной дифференциации предложено учитывать несколько признаков: окраску листьев, фенольную ре- акцию, длину опушения цветковых чешуи (или длину волосков апикулю-са), чувствительность к низким температурам, хлорату калия (табл. 2). При использовании вышеперечисленной комбинации признаков погрешность классификации не превышает 0,5 % (29-31). Указанные в этой таблице признаки расположены в порядке их применимости для дискриминации образцов. Так, устойчивость к хлорату калия позволяет более точно дифференцировать образцы на подвиды. Для сортов индийского подвида в большинстве случаев характерен позитивный ответ на фенольную реакцию (32, 33). Хотя спектр вариации по каждому признаку для подвидов частично перекрывается, возможность дифференциации все же указывает на наличие у них значительных и разнонаправленных модификаций.
2. Признаки, дифференцирующие подвиды indica и japonica у риса ( Oryza saliva L.) (33)
Признак
Устойчивость к хлорату калия (log повреждающей концентрации)
Устойчивость к засухе (выживание, %)
Длина волосков апикулюса (мм)
Устойчивость к холоду (%)
Фенольная реакция
indica ( n = 42)
Низкая (3,34±0,60)
Высокая (92,4±8,75)
Короткие (0,26±0,06)
Чувствительный (0,58±0,18)
Позитивная
| japonica ( n = 47)
Высокая ( - 1,49±0,35)
Низкая (59,3±20,67)
Длинные (0,44±0,22)
Устойчивый (0,18±0,22)
Негативная
Еще один вариант аттрибутирования предложен китайскими учеными, которые считают целесообразным в перечень показателей, позволяющих достоверно разделить образцы риса на подвиды, включить (в дополнение к уже перечисленным) отношение числа больших сосудистых пучков к числу малых в ножке метелки. У подвида indica указанный индекс приблизительно равен 1, у japonica — 0,5 (18). Следует отметить, что даже тест на стерильность гибридов не всегда позволяет правильно классифицировать образцы. В частности, скрещивание образцов, несущих некоторые гены, характерные для диких и предковых форм, с современными сортами также приводит к частичной стерильности гибридов (2).
3. Озерненность у гибридов риса ( Oryza saliva L.) первого поколения при скрещивании родительских форм подвидов japonica (j) и indica (i) из различных эколого-географических групп (вегетационная площадка, 2008-2009 годы)
Материнская форма х опылитель | Пустозерность, % japonica х indica:
|
Хазар х Олимпиада |
50,90 |
|
Дружный х Олимпиада |
65,10 |
|
ВНИИР 7679 х Паритет |
59,64 |
|
Донг Нонг 418 (japonica тропическая) х Стрелец |
83,98 |
|
Vialone Nano х а/12 6205 |
58,27 |
|
Жемчуг х а/12 6206 |
54,50 |
|
indica / japonica : А/12 6205 х Приморский |
93,30 |
|
А/12 6206 х Аметист |
40,00 |
|
А/12 6205 х Baldo |
74,90 |
|
А/12 6205 х Атлант |
41,20 |
|
А/12 6205 х Славянец |
20,70 |
|
А/12 6205 х Дончак |
56,89 |
|
А/12 6205 х Лиман |
32,04 |
|
Jefferson х Arborio |
8,02 |
|
Jefferson х Аметист |
6,73 |
|
Jefferson х Виола |
7,71 |
|
Jefferson х Фонтан |
3,75 |
|
Внутриподвидовые гибриды: Донг Нонг 418 (japonica тропическая) х Боярин (j) |
46,82 |
|
Донг Нонг 418 (japonica тропическая) х Павловский (j) |
32,85 |
|
А/12 6205 (i) х Изумруд (i) |
89,70 |
|
Снежинка (i) х KCL (i) |
30,59 |
В проведенном нами исследовании гибридов в комбинациях, где родительские формы принадлежали к различным эколого-географическим группам, была обнаружена высокая
4. Распределение полиморфных SSR-маркеров, позволяющих типиро-вать подвиды japonica и indica , на хромосомах риса ( Oryza sa-tiva L.) (7)
|
Хромосома |
| SSR-маркер |
|
1 |
RM 226, RM 259, RM 23 |
|
2 |
RM 250, RM 29, RM 211, RM 240 |
|
3 |
RM 231, RM 232, RM 16, RM 251 |
|
4 |
RM 255, RM 335 |
|
5 |
RM 249, RM 267 |
|
6 |
RM 190 |
|
7 |
RM 248, RM 234, RM 214, RM 18 |
|
8 |
RM 25 |
|
9 |
RM 242 |
|
10 |
RM 258, RM 222, RM 228 |
|
11 |
RM 4 |
|
12 |
RM 247, RM 19 |
пустозерность растений F i (табл. 3).
Полученные данные показывают отсутствие генов широкой совместимости у образцов, вовлеченных в гибридизацию. Только образец из США Jefferson во всех комбинациях имел значительную озерненность гибридов (в вариантах с ним пустозерность варьировала от 3,75 до 8,00 %). Анализ генезиса сорта подтвердил, что причиной высокой озерненности гибридов в этом случае не может быть близкородственное происхождение исходных форм. О том же свидетельствовали значительные различия по морфофизиологическим характеристикам у
образцов, вовлеченных в гибридизацию с этим сортом.
Наиболее достоверно разделить образцы на подвиды можно с помощью молекулярного маркирования. В настоящее время для подобной дифференциации предлагается использовать ряд типичных для подвидов риса полиморфных SSR-маркеров, распределенных по всему геному (табл. 4). Образцы принято относить к тому подвиду, большее число характерных аллелей которого они несут (7). Для того чтобы исключить ошибочное отнесение сортов к подвиду indica, мы также провели их моле кулярное маркирование.
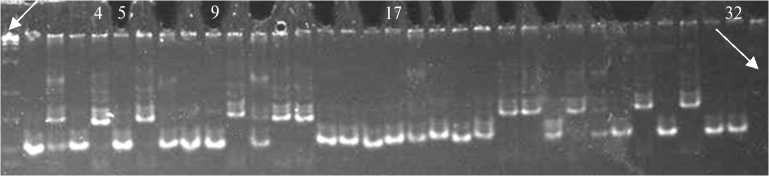
Рис. 1. Полиморфизм у 32 сортов риса ( Oryza sativa L.) отечественной селекции, выявляемый с использованием SSR-маркера RM 335: 4 — Шарм, 5 — Снежинка, 9 — Серпантин, 17 — Нарцисс, 32 — Изумруд (сорта российской селекции, подвид indica ); остальные дорожки — сорта российской селекции подвида japonica . Маркер молекулярной массы GenPak DNA Markers M 50 (ООО «Лаборатория Изоген», Россия) (отмечен стрелкой).
Изучение генома с использованием 11 SSR-маркеров показало, что российские сорта, относимые к подвиду indica , полиморфны по некоторым из этих маркеров, например по RM 335 (рис. 1), RM 240, RM 247, RM 25, RM 259, RM 242. Мономорфными у всех российских сортов оказались маркеры RM 255, RM 258, RM 4, RM 29, RM 19 (рис. 2, А и Б). По мономорфным маркерам все изученные отечественные сорта несли аллель, характерный для подвида japonica , по полиморфным — у большей части этих сортов, относимых к подвиду indica , также имелись аллели, типичные для подвида japonica (табл. 5). Так, в 11 изученных локусах у сортов Хан-кайский, Приморский, Изумруд были выявлены только три аллеля, специфичных для подвида indica , у сортов Нарцисс и Шарм — четыре, у сорта Снежинка — один аллель, типичный для этого подвида. Только сорт Серпантин может быть отнесен к подвиду indica на основании анализа с использованием изученных маркеров, которые сцеплены с локусами, полиморфными для образцов подвидов. С точки зрения разделения российских образцов на подвиды наиболее информативными оказались маркеры
RM 335, RM 240, RM 247, RM 25, RM 259 и RM 242.
A
Б
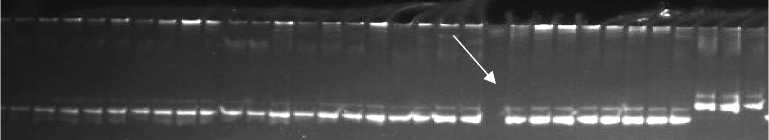
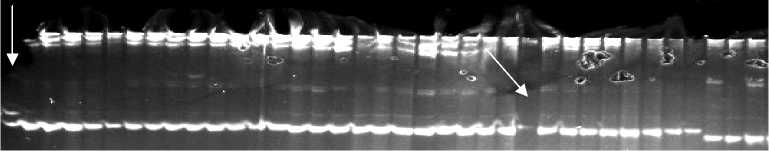
Рис. 2. Полиморфизм у сортов риса ( Oryza sativa L.) разного эколого-географического происхождения, выявляемый с использованием SSR-маркеров RM 29 (A) и RM 19 (Б) : три последние дорожки справа — типичные сорта подвида indica зарубежной селекции, остальные дорожки — сорта российской селекции. Маркер молекулярной массы GenPak DNA Markers M 50 (ООО «Лаборатория Изоген», Россия) (отмечен стрелкой).
5. Полиморфизм у сортов риса ( Oryza sativa L.) отечественной селекции, выявляемый с использованием SSR-маркеров, типичных для подвидов indica и japonica
|
Cорт |
i |
см i |
гм i |
см i |
см см i |
i |
04 i |
i |
(М i |
(М i |
ОО гл i |
Доля аллелей, не характерных для подвида japonica |
|
Ханкайский 1 2 1 2 2 1 2 1 2 2 1 3/11 Приморский 1111112 12 11 3/11 Серпантин 1112 2 12 1111 6/11 Нарцисс 12 12 112 1111 4/11 Снежинка 2 2 1 2 1 1 2 1 2 2 1 1/11 Шарм 12 12 2 12 112 1 4/11 Изумруд 1 2 1 2 1 1 2 1 2 1 1 3/11 japonica 22111121221 0 ________ p p m p p m m m р pm Примечание. p и m — соответственно полиморфный и мономорфный маркер. |
||||||||||||
Таким образом, наблюдаемую у межподвидовых гибридов высокую озерненность при скрещивании российских образцов можно объяснить их генетическим родством и вероятным неправильным отнесением к подвиду indica по индексу зерновки. По результатам анализа озернен-ности гибридов установлено, что вовлеченные в скрещивания сорта российской селекции не несут гены широкой совместимости, определяющие высокие значения этого показателя у гибридов. Продемонстрирована необходимость включения дополнительных характеристик в перечень признаков, на основании которых проводится разделение отечественных образцов на подвиды indica и japonica , для сокращения числа некорректных идентификаций.
