Стрессовая память растений: ключевые аспекты (обзор)
")
Автор: Щенникова А.В., Филюшин М.А., Кочиева Е.З.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Обзоры, проблемы
Статья в выпуске: 3 т.60, 2025 года.
Бесплатный доступ
Растения постоянно подвергаются разрушительному воздействию неблагоприятных экологических условий - как абиотических, так и биотических. В случае сельскохозяйственных культур это негативно сказывается на урожайности. Многочисленные лабораторные исследования сфокусированы на влиянии различных стрессовых факторов на рост и развитие растений. Результаты попыток переноса получаемых данных на полевые условия культивирования свидетельствуют о присутствии критических аспектов взаимодействия растение-стресс. В представляемом обзоре мы рассматриваем некоторые из этих аспектов. Так, большинство работ сфокусировано на изучении молекулярных реакций растений на однофакторные стрессы (Y. Saijo с соавт., 2020). Для естественной среды характерны многофакторные воздействия в различных сочетаниях. Поэтому в последнее время растет число публикаций с анализом реакции растений на двух- и трехфакторные стрессовые комбинации абиотических факторов с абиотическими (T. Obata с соавт., 2015; N. Suzuki с соавт., 2016; S.I. Zandalinas с соавт., 2022) и биотическими (S. Rasmussen с соавт., 2013; N. Suzuki с соавт., 2014; A.R. Devireddy с соавт., 2020) стрессорами. Показано, что механизм ответа растения при комбинаторном воздействии возникает как результат интеграции и модулирования ответов на индивидуальные составляющие стресса (N. Suzuki с соавт., 2014), которые по отношению друг к другу могут быть нейтральными, антагонистическими или несвязанными, что часто непредсказуемо и нуждается в подтверждении опытным путем (S. Rasmussen с соавт., 2013). Ответы на абиотические и биотические факторы контролируются разными, иногда конфликтующими сигнальными путями, которые при комбинаторном стрессе могут взаимодействовать и/или ингибировать друг друга (S. Rasmussen с соавт., 2013; N. Suzuki с соавт., 2014). Качество стрессового ответа растения зависит не только от генотипа, но и от эпигенетической регуляции экспрессии соответствующих генов и накопления необходимых метаболитов (Z.H. Chen с соавт., 2020), известной как стрессовая память (A. Weinhold, 2018). В естественной среде произрастания растений нередки возвратные события однотипных и комбинаторных стрессов. Формирование обратимой наследуемой стрессовой памяти происходит как результат переживания растением первичного стрессового воздействия (прайминг). Растение запоминает свою защитную реакцию посредством внесения в геном эпигенетических изменений и при повторе стресса запускает запомнившиеся реакции на предыдущий опыт (M. Hilker с соавт., 2016; J. Lämke с соавт., 2017). Различают цис-прайминг, когда вид первичного и повторного воздействия совпадает, и транс-прайминг, когда память о прайминге срабатывает в ответ на стимул другого типа (M. Hilker с соавт., 2016; A.U. Nair с соавт., 2022). Набор эпигенетических меток, возникший после прайминга, составляет память растения о стрессовом событии и направлен на регуляцию экспрессии конкретных генов (M. Hilker с соавт., 2016; L. Virlouvet с соавт., 2018; A. Nishad с соавт., 2021; C. Jacques с соавт., 2021). Стрессовая память может быть кратковременной (митотически наследуемой) и долговременной (мейотически наследуемой) (M. Hilker с соавт., 2016; J. Lämke с соавт., 2017). В отсутствие повторных стрессовых событий происходит забывание, когда растение активирует эпигенетические регуляторные механизмы, сбрасывающие метки стрессовой памяти (A. Wibowo с соавт., 2016). Защитный ответ растения на отдельные стрессоры и их сочетание при прайминге и повторных событиях выражается в существенном модулировании экспрессии генов и содержания различных метаболитов (A. Aharoni с соавт., 2011; S.I. Zandalinas с соавт., 2022). Транскрипты и метаболиты с дифференциальной реакцией на стресс могут служить биомаркерами стрессовой памяти (O. Aina с соавт., 2024). Характерной особенностью биомаркеров (генов, метаболитов) считаются существенные различия в колебаниях экспрессии (гены) и содержания (метаболиты) при прайминге, повторных стрессах и возвратах в нормальные условия (Y. Ding с соавт., 2013, 2014; C. Jacques с соавт., 2021). Предполагаемые гены-биомаркеры могут быть ассоциированы с осуществлением метаболизма различных соединений, включая компоненты антиоксидантной защиты растения, фитогормоны, патогенез-связанные PR-белки и др. (O.K. Anisimova с соавт., 2021; S.I. Zandalinas с соавт., 2022; O. Aina с соавт., 2024), а также с регуляцией генной экспрессии (транскрипционные факторы, некодирующие РНК, циркадные осцилляторы, др.) (H. Wei с соавт., 2022; W. Cao с соавт., 2024). На основе изложенного материала предполагается, что исследования защитных/адаптивных реакций растений, особенно культивируемых, направленные на повышение стрессоустойчивости, должны проводиться с учетом обсуждаемых в обзоре аспектов. Результатом таких исследований может стать определение ключевых биомаркеров памяти растения о воздействиях различных стрессовых факторов и их комбинаций и, как следствие, выявление доноров эпиаллелей/эпимутаций, ассоциированных с наследуемой стрессовой памятью.
Стрессовые факторы, стрессовая память растения, биомаркеры стрессовой памяти
Короткий адрес: https://sciup.org/142246194
IDR: 142246194 | УДК: 631.522/.524 | DOI: 10.15389/agrobiology.2025.3.397rus
Stress memory in plants: key aspects (review)
Plants are constantly exposed to the destructive effects of unfavorable environmental conditions, both abiotic and biotic. In the case of crops, this negatively affects yields. Numerous laboratory studies have focused on the effects of various stress factors on plant growth and development. The results of attempts to transfer the obtained data to field cultivation conditions indicate the presence of critical aspects of the plant-stress interaction. In this review, we consider some of these aspects. Thus, most studies are focused on the analysis of molecular responses of plants to single-factor stresses (Y. Saijo, et al., 2020). The natural environment is characterized by multifactorial effects in various combinations. Therefore, recently there has been an increasing number of publications analyzing the response of plants to two- and three-factor stress combinations of abiotic factors with abiotic (T. Obata et al., 2015; N. Suzuki et al., 2016; S.I. Zandalinas et al., 2022) and biotic (S. Rasmussen et al., 2013; N. Suzuki et al., 2014; A.R. Devireddy et al., 2020) stressors. It has been shown that the mechanism of plant response under combinatorial influence arises as a result of integration and modulation of responses to individual stress components (N. Suzuki et al., 2014), which can be neutral, antagonistic or unrelated to each other, and is often unpredictable and requires experimental confirmation (S. Rasmussen et al., 2013). Responses to abiotic and biotic factors are controlled by different, sometimes conflicting signaling pathways, which under combinatorial stress can interact and/or inhibit each other (S. Rasmussen et al., 2013; N. Suzuki et al., 2014). The quality of the plant stress response depends not only on the genotype, but also on the epigenetic regulation of the expression of the corresponding genes and the accumulation of the necessary metabolites (Z.H. Chen et al., 2020), known as “stress memory” (A. Weinhold, 2018). In the natural environment of plant growth, recurrent events of the same type and combinatorial stresses are not uncommon. The formation of reversible heritable stress memory occurs as a result of the plant experiencing a primary stress effect (priming). The plant remembers its defense reaction by introducing epigenetic changes into the genome and, when the stress is repeated, triggers the remembered reactions to the previous experience (M. Hilker et al., 2016; J. Lämke et al, 2017). A distinction is made between cis-priming, when the type of primary and repeated stresses coincides, and trans-priming, when the priming memory is triggered in response to a stimulus of a different type (M. Hilker et al., 2016; A.U. Nair et al., 2022). The set of epigenetic marks that arise after priming constitutes the plant's memory of the stressful event and is aimed at regulating the expression of specific genes (M. Hilker et al., 2016; L. Virlouvet et al., 2018; A. Nishad, A.K. Nandi, 2021; C. Jacques et al., 2021). Stress memory can be short-term (mitotically inherited) and long-term (meiotically inherited) (M. Hilker et al., 2016; J. Lämke et al., 2017). In the absence of repeated stressful events, forgetting occurs when the plant activates epigenetic regulatory mechanisms that reset stress memory marks (A. Wibowo et al., 2016). The plant's defense response to individual stressors and their combination during priming and repeated events is expressed in significant modulation of gene expression and the content of various metabolites (A. Aharoni et al., 2011; S.I. Zandalinas et al., 2022). Transcripts and metabolites with a differential response to stress can serve as biomarkers of stress memory (Aina et al., 2024). A characteristic feature of biomarkers (genes, metabolites) is considered to be significant differences in fluctuations in expression (genes) and content (metabolites) during priming, repeated stresses and returns to normal conditions (Y. Ding et al., 2013, 2014; C. Jacques et al., 2021). Putative biomarker genes may be associated with the metabolism of various compounds, including components of plant antioxidant defense, phytohormones, pathogenesis-associated PR proteins, etc. (O.K. Anisimova et al., 2021; S.I. Zandalinas et al., 2022; O. Aina et al., 2024), as well as with the regulation of gene expression (transcription factors, non-coding RNAs, circadian oscillators, etc.) (H. Wei et al., 2022; W. Cao et al., 2024). Based on the presented material, it is assumed that studies of plant defense/adaptive responses, especially cultivated plants, aimed at increasing stress resistance, should be carried out taking into account the aspects discussed in the review. The result of such studies may be the identification of key biomarkers of plant memory about the effects of various stress factors and their combinations, and, as a consequence, the identification of donors of epialleles/epimutations associated with inherited stress memory.
Текст обзорной статьи Стрессовая память растений: ключевые аспекты (обзор)
Весь жизненный цикл большинства растений проходит в месте укоренения, что обусловливает неизбежность и необходимость адаптации к неблагоприятным воздействиям окружающей среды. При этом стрессовыми факторами можно назвать любые неожиданные изменения условий, выбивающиеся из привычных рамок. В первую очередь это абиотические факторы, например изменения уровня освещенности, экстремальные температуры, засуха, избыток воды, засоленность, нехватка/избыток микроэлементов в почве. К биотическим факторам относят инфицирование фитопато-генными микроорганизмами (вирусы, бактерии, грибы, нематоды) и атаки насекомых-вредителей и травоядных животных (1).
Стрессовые эффекты, с одной стороны, вредоносны, но с другой стороны представляют собой движущую силу адаптационной эволюции и морфологической диверсификации растений (2). Более того, благодаря им существуют современные сельскохозяйственные культуры как результат доместикации и селекции соответствующих видов растений под влиянием различных стрессовых факторов; причем в сравнении с дикими сородичами одомашненные культуры сочетают повышенную адаптивность к контрастным средам с низкой устойчивостью к абиотическим стрессам (2, 3). Адаптационную эволюцию связывают в числе прочего с частым возникновением событий дупликации геномов (полиплоидии) и генов, что сопровождается возникновением генных мутаций, неофункционализацией генов и их аллельных вариантов, а также увеличением размеров генных семейств, связанных с сигнальными путями стрессовых ответов (2).
Однако качество стрессового ответа растения зависит не только от геномных мутаций, но и от механизмов регуляции экспрессии соответствующих генов и накопления необходимых метаболитов (2). Исследования эпигенетических регуляторных механизмов у растений привели к появлению понятия «стрессовая память» (4).
В последние годы наблюдается взрывной рост числа публикаций по механизмам эпигенетических модификаций и регуляций генной экспрессии, а также работ по ответу растений на различные стрессовые факторы и по стрессовой памяти. Поэтому целью представляемого обзора стало не повторение описания молекулярных путей стрессовых реакций (включая модификации эпигенома), а определение и сбор воедино особенностей и аспектов воздействия стрессовых факторов на растения на основе опубликованных данных. Учет таких аспектов, с нашей точки зрения, может способствовать более точной и четкой постановке тематических экспериментов и корректной интерпретации получаемых данных.
Стрессовое воздействие: однофакторное или комбинированное. Большинство исследований реакций растений на стрессовые условия описывают эффекты одиночных факторов. Однако для естественной среды, как правило, характерны стрессовые воздействия сразу нескольких факторов, различные сочетания которых могут влиять на развитие и продуктивность растения как негативно, так и позитивно. При этом молекулярно-генетическая реакция растения на комбинаторное воздействие значительно более сложная, чем на каждый из факторов по отдельности.
Усиление негативного воздействия при комбинации индивидуальных стрессовых факторов предполагает определенную степень независимости между механизмами ответа растения на факторы по отдельности или их конфликт друг с другом (антагонистичность). Это хорошо иллюстрируется состоянием устьиц при сочетании стрессов. К примеру, при тепловом стрессе растение увеличивает устьичную проводимость для охлаждения листьев, тогда как сочетание жары и засухи не дает раскрывать устьица, предотвращая потерю влаги и приводя к перегреву (5). При чрезмерной освещенности и инфицировании патогенами устьица закрываются, что также не согласуется с эффектом повышенной температуры (5). При совпадении воздействия на устьица отдельных факторов их комбинация может смягчать стрессовый эффект одного фактора за счет другого. Например, засуха снижает устьичную проводимость и при сочетании с озонированием (О3) повышает устойчивость растения к О3 (6).
Таким образом, при комбинации стрессовых условий возникает уникальный механизм защитного ответа, который основан на интеграции и модулировании индивидуальных ответов (7) и теоретически может представлять собой сочетание однофакторных нейтральных, антагонистических или несвязанных реакций транскриптов генов (8).
Однако другие работы по исследованию влияния различных парных сочетаний стрессовых факторов показали, что все не так просто и предсказуемо. Основополагающие для прогнозирования данные были получены при сравнении транскриптомов 10 экотипов модельного растения Arabidopsis thaliana после симуляции воздействия индивидуальных стрессовых факторов (холод, жара, яркий свет, избыток соли, элиситор флагеллин) и их двойных комбинаций (8).
Авторы провели сравнительный анализ взвешенных сетей коэкс-прессии генов (WGCNA) и предложили классифицировать ответные реакции с учетом динамики дифференциально экспрессирующихся между вариантами транскриптов (8). Первые три модели включают транскрипты, реакцию которых на двухфакторный стресс невозможно предсказать на основе однофакторных экспериментов. Сюда вошли транскрипты со сходным ответом на два стресса по отдельности, но с разным — на их сочетание (комбинаторная модель), а также транскрипты с неодинаковой реакцией на каждый из двух факторов, остающиеся на одном из достигнутых уровней (приоритетная модель) или возвращающиеся к дострессовым значениям (модель отмены) в ответ на комбинированный стресс. Другие две модели учитывают предсказуемые транскрипты, чья реакция на один или оба одиночных фактора не меняется при комбинаторном стрессе (независимая модель) или одинакова как для отдельных факторов, так и для их сочетания (аналогичная модель) (8). Было показано, что при ответе растения на парные стрессы большинство (около 61 %) дифференциально экспрессирующихся транскриптов действует по первым трем моделям (комбинаторная, приоритетная, отмена), соответственно около 28 и 11 % — по независимой и аналогичной моделям (8).
Эти данные наглядно продемонстрировали существенные различия механизма защитного ответа на сочетания стрессовых факторов в сравнении с реакциями на стрессоры по отдельности и, более того, непредсказуемость и связанную с этим необходимость определения такого механизма опытным путем.
В той же работе на основе полученных транскриптомных данных были определены пары стрессовых условий, вызывающие сходные (холод и яркий свет, соль и повышенная освещенность) или антагонистические с непредсказуемостью эффекта двухфакторного влияния (соль и холод, соль и жара, абиотические и биотические факторы) ответы растения. Также были предложены регуляторные модули, состоящие из транскрипционных факторов (ТФ), которые ассоциированы с ответом растения на индивидуальные и парные абиотические стрессоры (8). Такие обобщения, представленные в этой и подобных работах, могут успешно использоваться для создания моделей по предсказанию реакций растений на многофакторные стрессы.
Защитный ответ растения на индивидуальные стрессоры и их сочетание выражается и в модулировании содержания различных метаболитов, синтез которых координируется генными сетями, связывающими первичный и вторичный метаболизм (9, 10). Прежде всего это первичные метаболиты, такие как сахара и аминокислоты. Регулирование количества сахаров имеет решающее значение для выживания и адаптации растения в неблагоприятных условиях и зависит от типа стрессового фактора и от вида растения. Как и в случае экспрессии генов, реакции углеводного обмена при многофакторном воздействии могут быть предсказуемыми (по реакции на одиночные стрессы) и непредсказуемыми (11). В целом при большинстве абиотических стрессов и их сочетаниях характерно повышение содержания растворимых сахаров, действующих как осмопротекторы (12).
При стрессе растение вынужденно тратит энергию на адаптацию и для компенсации дефицита энергии модифицирует метаболизм аминокислот, используемых дыхательными путями в качестве субстратов (13). Отмечено, что индуцированные изменения количества различных аминокислот специфичны для типа стрессового фактора/факторов и могут способствовать осмопротекции и нейтрализации активных форм кислорода (АФК) (11, 13). Деградация цистеина приводит к закрытию устьиц (13). Более того, некоторые аминокислоты служат триггерами усиленного синтеза важных для защитного ответа растений вторичных метаболитов, к примеру абсцизовой кислоты (АБК) (13) или полиаминов (9, 14).
Вторичные метаболиты растения можно разделить на три основные группы: фенольные соединения, терпены и азот- или серосодержащие соединения (10). Все они играют важные роли в защите растений от окислительного стресса как результата воздействия неблагоприятных факторов и их сочетаний, что подтверждено экспериментально (9, 15). Среди метаболитов отдельно можно выделить фитогормоны, специфически накапливающиеся при стрессах. Ключевую роль отводят АБК, которая координирует свое действие с другими гормонами и/или сигнальными путями в зависимости от типа стрессовых факторов и их сочетаний (16-18).
Суммируя имеющиеся на данный момент сведения, можно отметить, что в большинстве современных исследований оценивается воздействие на растение однофакторных стрессов, включая определение генетических механизмов реакции на индивидуальные стрессоры, как, например, в работе Y. Saijo с соавт. (19). Одновременно растет число публикаций по изучению совместного влияния двух и иногда трех неблагоприятных условий, включая сочетания абиотических факторов (9, 11, 17), а также абиотических и биотических стрессоров (7, 8, 18). Однако пока ни в одном исследовании не выполнена оценка влияния многофакторного (более трех составляющих) абиотического стресса на генные и метаболические пути. А это особенно важно, так как в естественной среде растение подвергается именно многофакторному стрессу с часто меняющимися комбинаторными вариантами. С увеличением числа и сложности факторов, действующих одновременно, рост и выживаемость растений снижаются, даже если индивидуальный эффект каждого из факторов незначителен (9).
Сочетания абиотических и биотических стрессовых факторов. Таким образом, реакция растений на каждую комбинацию стрессовых 400
условий уникальна и не может быть экстраполирована, исходя из реакций на каждый из участвующих факторов. Все становится еще менее понятным, если к абиотическим воздействиям добавить биотические (фитопатогены и травоядные вредители), с которыми в естественных условиях растения сталкиваются постоянно (19, 20). Ответы на абиотические и биотические стрессоры контролируются разными, иногда конфликтующими сигнальными путями, которые при комбинаторном стрессе могут ингибировать друг друга (7, 8). При этом неблагоприятные абиотические воздействия ослабляют растение, что часто повышает его восприимчивость к инфекции (7). К примеру, экстремальные температуры нарушают защиту растений от патогенов (21). Однако известны случаи, когда абиотический стресс снижает восприимчивость к инфицированию (19, 20). Например, солевой стресс подавляет рост возбудителя мучнистой росы ( Oidium lycopersici ) у томата (22), а засуха и O 3 стимулируют накопление связанных со стрессовым ответом белков (PR, pathogenesis-related proteins) (6). В целом абиотические стрессы снижают устойчивость к гемибиотрофным и некротрофным патогенам и повышают сопротивляемость к биотрофам (23).
Транскриптомный анализ растений A. thaliana при воздействии засухи, жары, вирусной инфекции, а также их двойных и тройных комбинаций показал невозможность предсказания изменения генной экспрессии при сочетании стрессоров на основе данных по ответам на отдельные факторы (24). Тем не менее авторам удалось определить транскрипты генов 11 ТФ с дифференциальной экспрессией которых связаны все анализируемые в работе стрессовые реакции, а также 23 транскрипта, чья экспрессия не чувствительна к отдельным факторам и их парным комбинациям, однако специфически активируется при сочетании всех трех условий (24).
Известны случаи, когда при комбинации абиотических и биотических стрессоров растение реагирует приоритетным для одной из составляющих комбинации образом. Так, одновременное воздействие на A. thaliana патогенного элиситора флагеллина flg22 и ультрафиолетового излучения (УФ-В) стимулирует биосинтез соединений, противодействующих патогенезу, и при этом снижает накопление флавонолов, защищающих от УФ-В (25). Еще один пример: сочетание засухи и заражения нематодами у растений A. thaliana сопровождается изменениями экспрессии генов, большинство из которых ассоциированы с эффектом засухи (26).
Взаимодействие растения и биотической среды далеко не всегда носит характер патогенеза (19). При адаптации к неблагоприятным условиям растения могут прибегать к помощи микробных сообществ, ассоциированных с корневой системой, и в процессе стрессового ответа микробиом способен изменять активность и состав (27-29). При этом у растения формируется биотический иммунитет как ответ на изменение микробиома при вторжении патогена в условиях абиотического стресса (27-29).
В последнее время механизмы реакции растения на сочетания абиотических и биотических стрессоров активно исследуются, становясь предметом обзорных публикаций (19). Получаемые данные каждый раз подчеркивают сложную природу реакций на множественный стресс и, несомненно, важны для будущего прогностического моделирования ответов растения на многофакторные воздействия при сочетании абиотических и биотических условий.
Таким образом, при одновременном действии нескольких стрессоров (комбинации абиотических факторов друг с другом или с биотическими факторами) растение инициирует в значительной степени перекрывающийся набор сигнальных путей, после чего оптимизирует ответ, определяя его приоритетные направления и модулируя их активность и продолжительность. В каждый конкретный момент, по всей вероятности, происходит оценка эффективности реакции на каждую составляющую стрессовой комбинации, на основании чего могут меняться приоритетные направления. Такая многоуровневая регуляция осуществляется в условиях постоянного изменения характеристик стрессового воздействия (силы, продолжительности и комбинаторного состава) и динамики растительно-микробных ассоциаций. С учетом этого следует признать, что наше понимание модулирования иммунитета растений окружающей средой все еще весьма фрагментарно.
Стрессовая память растения. Немаловажным обстоятельством в сложной схеме взаимодействий растения со стрессовыми факторами служит то, что сходные типы воздействий (как отдельных, так и комбинаторных) могут повторяться несколько раз в течение жизненного цикла растения. Переживание первого прецедента «учит» растение защищаться от его повторов с помощью приобретаемой стрессовой памяти (30-32). При этом подчеркнем несколько важных, с нашей точки зрения, аспектов.
Запоминание . Первичное стрессовое событие, в процессе которого растение запоминает свою защитную реакцию и готовит себя к более эффективному ответу на повтор, получило название прайминга (или прайми-рования). При повторном воздействии происходит активация запомнившихся реакций на предыдущий опыт. Прайминг не меняет нуклеотидную последовательность генома, но инициирует в растении эпигенетические и затем клеточные, гормональные и другие изменения, что не мешает вернуться к исходному, то есть дострессовому состоянию (33). Синонимы для обозначения прайминга — системная приобретенная устойчивость (SAR, systemic acquired resistance, или повышенная устойчивость к патогенам после воздействия их сублетальных доз), акклиматизация/закаливание (улучшение холодостойкости после предобработки холодом), индуцированная системная устойчивость (ISR, induced systemic resistance, обеспечиваемая за счет колонизации корневой системы полезными ризобактериями или симбиотическими грибами) (33).
Исходно праймированием называли предпосевную гидратацию семян (34). В ряде обзоров подробно изложены история этого вопроса и современные представления об этом процессе, включая описание наиболее часто используемых химических и биологических агентов, а также наночастиц и неионизирующих физических воздействий для стимуляции роста, развития и устойчивости будущих растений к различным стрессовым факторам (34-36). Как результат прайминга, в семенах происходят различные положительные морфологические, физиологические и метаболические изменения, повышается синтез и накопление АТФ, ферментных и неферментных компонентов антиоксидантной защиты и стрессовых белков, активируются процессы репарации ДНК (34-36). Все это приводит как к ускорению прорастания семян, так и к более эффективному и усиленному росту и развитию растения, включая повышенную урожайность (34-36).
Следствием множества исследований эффекта стимулирующих воздействий стало распространение применения праймирования на сами растения. К примеру, обработка проростков морской травы Posidonia oceanica повышенными температурами и различными концентрациями NaCl способствовала повышению их устойчивости к повторным стрессовым событиям, что сопровождалось индукцией экспрессии генов, связанных с фотосинтезом, эпигенетическими модификациями и реакцией на стресс (37, 38). В обзоре F. Zulfiqar с соавт. (39) подробно рассмотрены подходы к повышению 402
солеустойчивости сельскохозяйственных культур с использованием в качестве рабочего материала и семян, и рассады. Изучение влияния повторяющегося холодового стресса на проростки A. thaliana разного возраста продемонстрировало, что праймирование возможно на разных стадиях жизненного цикла растения (40).
Главными преобразованиями, происходящими в ответ на праймиро-вание, считаются эпигенетические модификации как совокупность наследуемых (митотически и/или мейотически) изменений в активности генов при сохранении их нуклеотидной последовательности. Модификации осуществляются с помощью эпигенетических механизмов, среди которых основными служат метилирование ДНК, посттрансляционные модификации гистонов и активность некодирующих РНК (нкРНК, ncRNA) (41).
Метилирование ДНК у растений происходит в содержащих цитозин консенсусах CG, CHG и CHH (где H — это A, T или C) по РНК-зависи-мому пути (RdDM); метилирование регуляторных областей гена приводит к остановке его транскрипции (41). Доступ и отсутствие доступа транскрипционного аппарата к ДНК-мишеням определяется также плотностью упаковки хроматина, которая связана с посттрансляционными модификациями (ацетилирование, метилирование, фосфорилирование, убиквитинирова-ние, сумоилирование) коровых и линкерного гистоновых белков с участием комплексов ремоделирования хроматина. К наиболее известным маркерам гистонового кода гетерохроматина относятся H3K9me и H3K27me, тран-скрипционно активного эухроматина — H3K9ac, H3K4me и H3K36me2/3 (41). С направленной регуляцией метилирования ДНК и плотности упаковки хроматина связывают некодирующие РНК (нкРНК, ncRNAs), включая малые (микроРНК miRNAs; малые интерферирующие РНК, миРНК, siRNAs) и длинные (днкРНК, lncRNAs) РНК, которые участвуют в сайленсинге генов (в том числе по пути RdDM) (41). Углубленное описание эпигенетических механизмов в растении дано в обзоре A. Hannan Parker с соавт. (41).
Эпигенетические модификации как основа стрессовой памяти растения (33) направлены на подавление или активацию экспрессии конкретных генов, что приводит к необходимым метаболическим изменениям. Целевые гены и метаболиты могут служить биомаркерами памяти о стрессе, что будет рассмотрено ниже. Модификации обратимы, что делает их особенно эффективными при быстрой адаптации растения к меняющейся среде (4, 42). Набор эпигенетических меток, появившихся после прайминга, составляет память растения о стрессовом событии.
Стрессовая память может сохраняться в течение жизненного цикла растения (кратковременная, соматическая, митотически наследуемая) и даже передаваться по наследству (долговременная, мейотически наследуемая) (33, 43). Во втором случае эпимутации (мутации, приобретенные во время вегетативного роста) наследуются благодаря тому, что зародышевые клетки растения берут начало от меристематических клеток (4). При этом условное мейотическое наследование происходит в первую очередь по женской линии (44).
Длительность соматической памяти зависит от продолжительности прайминга и от того, на какой стадии развития растения он произошел (14). Чем короче прайминг, тем эффективнее может быть запоминание, поскольку при быстрой отмене стресса быстрее и корректнее восстанавливается профиль экспрессии генов ответа. Это показано на примере растений A. thaliana при сравнении воздействия 5- и 45-минутного теплового стресса на реакцию и послестрессовое восстановление состояния клетки (45), а также при сопоставлении эффектов 24-часовой и 2-недельной обработки холодом на экспрессию генов пластидной антиоксидантной системы при повторном воздействии (46).
Долговременную память может наследовать первое поколение потомков или более чем два поколения (трансгенерационная память) (4, 33, 43). Ее возникновению и передаче могут способствовать, в частности, постепенно нарастающие неблагоприятные процессы (например, почвенные накопления тяжелых металлов или солей). При этом потомки не обязательно окажутся устойчивы к ним генетически, но будут реагировать на них с повышенной эффективностью (4).
Эпигенетическая природа трансгенерационной памяти подтверждается обнаружением в растениях эпиаллелей, стабильных в течение сотен лет (например, радиальная симметрия мутантного цветка Linaria vulgaris опосредована метилированием промотора гена морфогенеза цветка) (43). Немногие примеры подтвержденных случаев трансгенерационной памяти о стрессе также связаны в основном с паттерном метилирования ДНК (43). Основным критерием трансгенерационной памяти принято считать ее мей-отическую стабильность, распространяющуюся как минимум на одно поколение, не подвергшееся стрессу, и не зависящую от изменений в последовательности ДНК. За передачу памяти о стрессе потомкам ответственны эпигенетические модификации в зародышевых клетках. При этом механизм наследования считается преимущественно связанным с материнским организмом, в котором сигналы стрессовой реакции могут передаваться от сомы к зародышевой линии с помощью нкРНК, митохондриальных белков и других неизвестных факторов (47). Отмечается, что механистическое понимание формирования и проявления трансгенерационной стрессовой памяти остается фрагментарным и требует дальнейшего изучения (43, 47).
Забывание . Сохранение и применение стрессовой памяти требует энергетических затрат, которые берутся из ресурсов, в норме направленных на рост и развитие растения. В связи с этим становится актуальной обратимость эпигенетических меток памяти. Стиранием меток, или «стрессовым забыванием», объясняется краткосрочность соматической стрессовой памяти растения (48, 49). Долговременная память может стираться в ходе гаметогенеза и раннего эмбриогенеза, однако при бесполом размножении растений эпимутации могут сохраняться в клоновых поколениях (50).
Забывание происходит в отсутствие повторных стрессовых событий (в том числе при переносе семян на дальние расстояния) Для этого растение активирует эпигенетические регуляторные механизмы, удаляющие метки, связанные с наследуемыми адаптивными реакциями, как показано на примере ответа A. thaliana на гиперосмотический стресс (44). Скрининг мутантов A. thaliana с нарушенной способностью приобретать стрессовую память выявил два регулятора хроматина — DDM1 и MOM1, чье совместное действие восстанавливает предстрессовое состояние хроматина, стирая эпигенетические метки (51).
Цис- и транс-прайминг . В зависимости от типа триггерного стрессового события (повторного стимула) различают цис-прайминг, когда стимул связан с таким же стрессовым фактором, что и при прайминге, и транс-прайминг, когда память о прайминге срабатывает в ответ на стимул другого типа (14, 33).
Примерами транс-прайминга может служить повышение солеустой-чивости растения после предварительного воздействия УФ-В-облучения (52), холода (53) или перекиси водорода (54). Устойчивость к обводнению проявляется после прайминга семян салициловой кислотой или селеном (55). Обработка саженцев мышьяком дает защиту от теплового стресса (56), 404
а тиомочевиной или засухой — от низких температур (57, 58). Эффект транс-прайминга объясняется тем, что ассоциированные стрессовые факторы по отдельности вызывают у растения сходную реакцию транскриптома (например, холод и повышенная освещенность (59)), даже если сигнальные пути ответа различаются.
Биомаркеры стрессовой памяти. Метаболиты и гены . Воздействие стрессоров при каждом повторе вызывает существенные изменения в транскриптоме и метаболоме растения, и часть транскриптов и метаболитов с изменившимся уровнем экспрессии и содержанием, соответственно, может служить биомаркерами, то есть индикаторами появления стрессовой памяти (60). Их может быть немало, так как механизмы стрессового ответа затрагивают накопление многих метаболитов, включая сахара, аминокислоты, флавоноиды, терпеноиды (61, 62), фитогормоны (49, 63, 64) и другие метаболиты. Примером последнего могут служить фосфорная, яблочная и янтарная кислоты, четырехатомный и пятиатомный спирты, а также метиловый эфир линолевой кислоты как маркеры устойчивости проростков пшеницы Triticum aestivum к ионам алюминия (65).
Характерной особенностью генов-биомаркеров (или генов памяти) является различная динамика профиля экспрессии при прайминге, повторном стимуле и возвратах в нормальные условия (фаза восстановления) (32, 66, 67). Гены, связанные с ответом на стресс, но не с памятью о нем, реагируют при прайминге и повторах воздействия одинаково, и в межстрессовый период их экспрессия быстро восстанавливается до предстрессового уровня (68). Экспрессия генов памяти меняется при прайминге и сохраняет изменение в фазу восстановления (I тип) или по-разному изменяется в каждом цикле стресса, а в фазы восстановления возвращается в норму (II тип) (69, 70). Согласно уточненной классификации, две группы генов памяти характеризуются индукцией (вариант реакции 1, +/+) или подавлением (2, - / - ) экспрессии и при прайминге, и при повторном стрессе, но при повторе степень модуляции меняется. Для других двух групп генов свойственна регуляция экспрессии, противоположная при прайминге и повторе: количество транскриптов снижается (прайминг) и повышается (повтор) (3, - /+) или наоборот (4, +/ - ) (32).
Таким образом, сравнительный анализ транскриптомного ответа растения на прайминг, повторы стрессового события и промежуточные возвраты в нормальные условия роста может выявить гены-биомаркеры. Например, анализ эффекта, оказываемого возвратной засухой на растения A. tha-liana , позволил отобрать более 2000 возможных генов памяти (гены-кандидаты), которые были ассоциированы с повышенным синтезом защитных соединений, восстановлением повреждений и детоксикацией, координацией роста и фотосинтеза, поддержанием гомеостаза и взаимодействием сигнальных путей, чувствительных к стрессу (66). Сравнение ответа на засуху у A. thaliana , кукурузы ( Zea mays ) и других видов растений сократило перечень до 556 генов как вероятных межвидовых биомаркеров; из них 18 % оказались ассоциированы с фотосинтезом (30, 32, 67, 71).
Кроме типа стрессового фактора, число и перечень биомаркеров зависит от генотипа, стадии развития, ткани и органа растения, а также от циркадных ритмов и может быть использовано для мониторинга реакции растений на стресс и селекции условно стрессоустойчивых сортов и линий сельскохозяйственных культур (60). Термин «условно» означает присутствие в геноме наследуемых эпигенетических меток, ассоциированных со стрессовой памятью. К примеру, анализ влияния засухи на такие биомаркеры, как содержание малонового диальдегида и активность антиоксидантных ферментов, позволил выявить генотипы кукурузы, наиболее успешно справляющиеся с осмотическим стрессом и, следовательно, перспективные в качестве доноров признака засухоустойчивости (72).
Примерами использования обнаруженных генов памяти могут послужить многочисленные работы по получению стрессоустойчивых растений методами генной инженерии. Так, многие гены, кодирующие ТФ семейства WRKY, при сверхэкспрессии или выключении повышают устойчивость растений томата ( Solanum lycopersicum ) к биотическим факторам (73). Сверхэкспрессия гена дегидрина ячменя ( HVA1 ) в трансгенных растениях пшеницы ( T. aestivum ) стимулирует их защиту от осмотических стрессоров (74). Сверхэкспрессия гена GhMYB36 (ТФ семейства MYB) увеличивает резистентность трансгенных растений A. thaliana и хлопчатника ( Gossypium hir-sutum ) к засухе и вертициллезному увяданию (75). Cверхэкспрессия генов VuNAC1 и VuNAC2 (ТФ семейства NAC) делает растения вигны ( Vigna unguic-ulata ) устойчивыми к засухе, засолению и экстремальным температурам (76).
Селекции стрессоустойчивых возделываемых растений может способствовать поиск источников признака резистентности посредством сравнения динамики содержания метаболитов-биомаркеров и/или уровня экспрессии генов памяти у образцов при стрессовом воздействии. В качестве примера можно привести упомянутый выше отбор доноров устойчивости к ионам алюминия среди образцов пшеницы (65) или выявление источников признака резистентности к засухе среди генотипов андского картофеля ( Solanum tuberosum ssp. tuberosum и ssp. andigena ) (77).
Нами также проводятся сравнительные исследования ответа на стрессовые факторы на примере контрастных по устойчивости сортов и линий чеснока ( Allium sativum ), кукурузы ( Z. mays ) и других культивируемых растений. Основываясь на полученных данных, можно выделить ряд возможных генов памяти среди генов PR-белков (78-81), унипортеров сахаров семейства SWEET (82), генов метаболизма аскорбата (83) и ТФ семейства DREB (84, 85).
Кроме того, использовав гидролизаты низкомолекулярного хитозана, мы праймировали растения двух сортов чеснока, различающихся устойчивостью к фузариозной гнили, и затем заражали их грибом Fusarium prolifer-atum (78). После прайминга с последующей инокуляцией F. proliferatum симптомы фузариоза не детектировались (в отличие от контрольных необрабатываемых растений), что указывало на повышение устойчивости к патогену у праймированных растений. Анализ экспрессии генов PR-белков (хитиназ и β -1,3-глюканаз) и соответствующей ферментной активности до и после инфицирования выявил PR -гены, которые предположительно могут служить в качестве биомаркеров стрессовой памяти (78).
Áèîìàðêåðû è öèðêàäíûå ðèòìû . После переноса растений из лаборатории в полевые условия стрессовый ответ часто меняется, что связано с увеличением числа стрессовых факторов. Особенно ярко проявляется зависимость защитной реакции от циркадных часов, которые синхронизируют суточные и сезонные колебания биологических процессов с циклическими изменениями сигналов окружающей среды (86, 87).
Циркадные ритмы тесно взаимосвязаны с окислительно-восстановительным статусом клеток, изменение которого происходит в ответ на любой стрессовый фактор (87). С одной стороны, на экспрессию генов, регулируемых циркадными часами, оказывает существенное влияние передача сигналов, опосредованная сахарами, особенно сахарозой, которая также связана с иммунитетом растения и его реакциями на абиотический стресс. С другой стороны, часы играют важную роль в контроле экспрессии генов углеводного метаболизма. В настоящее время достаточно активно исследуются участники сложной циркадной сети растений и их возможный вклад в новую концепцию «сладкого иммунитета» (88). Показано, что циркадные часы координируют сигнальные пути ответа растений на холодовой (89) и тепловой (90) стрессы. Компоненты циркадных часов действуют как важные интеграторы реакций растений на засуху, засоление и биотические факторы (91).
Таким образом, биомаркерами стрессовой памяти могут быть и гены циркадных осцилляторов, и регулируемые ими гены-мишени. К примеру, мишенями одного из ключевых осцилляторов — ТФ CCA1 служат гены, участвующие в передаче сигналов АБК (92). На примере растений риса показано, что активность OsCCA1 положительно связана с устойчивостью к различным абиотическим стрессовым факторам, что может осуществляться за счет регуляции АБК-сигнальных путей (92).
Если рассматривать гены-мишени циркадных часов, то эффективными биомаркерами могут также быть гены, ассоциированные с устьичной проводимостью и ассимиляцией CO 2 . В соответствии с суточными ритмами устьица днем открываются и ночью закрываются (93). Одновременно модуляция устьичной проводимости играет важную роль в ответе растений на абиотические и биотические стрессоры (5, 6, 94). Защита от осмотических стрессов (засуха, засоление), холода, чрезмерной освещенности и патогенной инфекции включает подавление дневного открытия устьиц (5, 6, 86, 95). Напротив, тепловой стресс и озонирование способствуют увеличению устьичной проводимости (5).
Биомаркеры и некодирующие РНК. В группу нкРНК входят, как уже отмечалось выше, малые (микроРНК, миРНК) и длинные (днкРНК) РНК, а также промежуточные (im-ncRNA) и кольцевые (circRNA) РНК. Все они считаются важными участниками регуляции экспрессии генов в ответе растений на абиотические и биотические стрессовые факторы (41, 96). Малые РНК участвуют в замалчивании генов посредством комплементарного связывания с их транскриптами и последующего расщепления этих транскриптов (микроРНК) или с помощью метилирования ДНК (RdDM-путь) и РНК-интерференции (миРНК). Участие днкРНК в регуляции экспрессии генов охватывает претранскрипционный (изменение топологии хроматина), транскрипционный (взаимодействие с ТФ) и посттранскрипционный (определение изоформ транскриптов при альтернативном сплайсинге) уровни влияния. Кольцевые нкРНК могут изменять уровень экспрессии генов, уменьшая взаимодействие миРНК с мРНК за счет конкурентного связывания или образования R-петель (96).
Обзор W. Cao с соавт. (96) углубленно рассматривает виды нкРНК растений и механизмы их влияния на экспрессию генов. В том числе в обзоре приведен список (данные на 2024 год) нкРНК, участвующих в реакциях растения на абиотические (засуха, жара, холод, засоление) и биотические (патогены и насекомые) стрессовые факторы (96). Перечисленные в обзоре нкРНК и, в перспективе, новые выявленные нкРНК могут выступать как биомаркеры стрессовой памяти растения.
Таким образом, биомаркеры стрессовой памяти растений могут включать: гены с определенным паттерном изменения экспрессии в ответ на повторяющиеся стрессы; первичные и вторичные метаболиты, включая компоненты ферментативной и неферментативной антиоксидантной защиты растения; фитогормоны; стрессовые белки; транскрипционные факторы; все типы нкРНК (рис.).
Экспрессия генов (структурные и регулярные гены метаболических путей, фотосинтеза, циркадных ритмов, патогенез-связанных белков и др.)
Компоненты антиоксидантной защиты: ферментативные (SOD, CAT, GR, АРХ, DHAR, MDHAR) и неферментативные (вторичные метаболиты: аскорбат, глутатион, каротиноиды, антоцианы, полифенолы, витамин Е)
Фитогормоны (АБК, СЛ, СК, ГК, ЭТ, ЖК)
Биомаркеры стрессовой
Первичные метаболиты, включая углеводы ► (крахмал, сахара) и аминокислоты
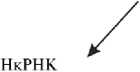
(микроРНК, миРНК, днкРНК, пиркРНК)
памяти растений

PR-белки, другие стрессовые белки
Транскрипционные факторы (семейства MYB, DREB, WRKY, MADS-box, Zink-flnger, другие)
Возможные биомаркеры стрессовой памяти растения: SOD — супероксиддисмутаза, CAT — каталаза, GR — глутатионредуктаза, APX — аскорбатпероксидаза, MDHAR — монодегидро-аскорбатредуктаза, DHAR — дегидроаскорбатредуктаза, АБК — абсцизовая кислота, СЛ — стриголактоны, СК — салициловая кислота, ГК — гиббереллины, ЭТ — этилен, ЖК — жасмоновая кислота, PR-белки — связанные с патогенезом белки.
В заключение отметим, что нашей целью было определение наиболее важных аспектов при изучении стрессового ответа растений. Поэтому в обзоре мы не приводим подробных сведений о молекулярно-генетических сигнальных путях, лежащих в основе реакций растения на неблагоприятные условия, особенно учитывая большое разнообразие известных стрессоров и их сочетаний, а также то, что ответные пути детализированы в недавних обзорах. Приведем лишь несколько примеров: реакция на холод (89, 97), засоление и засуху (98-100), различные абиотические и биотические факторы, включая некоторые их сочетания (101, 102), в том числе упомянутые выше (19, 20, 26); роль в стрессовом ответе циркадных часов (91). Получаемые молекулярно-генетические данные все чаще применяются наряду с традиционными подходами в селекции стрессоустойчивых высокопродуктивных сортов и линий агрокультур. Все более популярным становится не генно-инженерное получение сельскохозяйственных растений с нужными свойствами, а праймирование имеющихся культур эустрессорами для стимуляции появления эпигеномов с наследуемой устойчивостью к различным факторам и их сочетаниям (1). При этом важно помнить, что резистентность растения достигается не только посредством эпигенетической регуляции, элементы которой привносятся в геном с помощью прайминга стрессом, но и генетически. Вариабельность связанных с ответом на стрессы генов и регуляторных последовательностей между образцами одной культуры может коррелировать с устойчивостью к различным стрессорам, на что указывают проводимые пангеномные исследованиями разнообразия популяций (103).
Итак, многократное и/или длительное воздействие абиотических и биотических стрессовых факторов (как по отдельности, так и в различных комбинациях) оказывает деструктивное влияние на рост, развитие и продуктивность растений. К настоящему времени достаточно детально охарактеризованы реакции транскриптомов и метаболомов растений на основные типы неблагоприятных воздействий, на основании чего построены модели защитных/адаптивных механизмов. Перспективным представляется выявление биомаркеров стрессовой памяти, специфичных или общих для разных типов стресса, так как эту информацию можно использовать для поиска доноров хозяйственно ценных эпигеномов, ассоциированных с наследуемой стрессовой памятью.
ЛИТЕРАТУРЫ
Поступила в редакцию 22 июля 2024 года Принята к публикации 27 сентября 2024 года
Sel’skokhozyaistvennaya biologiya [ Agricultural Biology ], 2025, V. 60, № 3, pp. 397-414
STRESS MEMORY IN PLANTS: KEY ASPECTS (review)
A.V. Shchennikova ■ ■, M.A. Filyushin, E.Z. Kochieva
The authors declare no conflict of interests
Acknowledgements:
Supported financially by Russian Science Foundation (grant No. 24-16-00043, stress memory of plants) and the
Ministry of Science and Higher Education of the Russian Federation (effects of a combination of stressors)
