Строение верхнелохковской (нижний девон) органогенной постройки Лопъю-Кырта (р. Илыч, Северный Урал)
 органогенной постройки Лопъю-Кырта (р. Илыч, Северный Урал)")
Автор: Пономаренко Е.С.
Журнал: Вестник геонаук @vestnik-geo
Рубрика: Научные статьи
Статья в выпуске: 6 (282), 2018 года.
Бесплатный доступ
Изученные породы разреза скалы Лопъю-Кырта на р. Илыч (Северный Урал) представлены в основном биогермными известняками, а также известняковыми песчаниками, гравелито-песчаниками и брекчиями. Среди биогермных известняков преобладают строматолитовые разности, более редки коралловые, фистулеллово-микробиальные или строматопорово-микробиальные. Составление профилей и их взаимная корреляция позволили установить ландшафтно-морфологическую зональность постройки Лопъю-Кырта, выделить внешний склон и рифовое плато. В строении последнего элемента участвуют гребень рифа, песчано-гравийные отмели, центральная и тыловая зоны рифового плато. В целом строматолитовый состав каркаса и характерная рифовая зональность позволяют отнести Лопъю-Кырту к специфическому типу построек - строматолитовому рифу, своей зональностью резко отличающемуся от микробиальных холмов. Установлено, что Лопъю-Кырта имеет много общего с современным микробным рифом Хайбёрн Кэй (на Большой Багамской банке). Стратиграфически и географически более близкие к Лопъю-Кыртинской постройке верхнесилурийские строматолитовые рифы Северного Урала и Аляски значительно отличаются по архитектуре каркасов.
Нижний девон, лохковский ярус, строматолитовые рифы, палеоландшафтно-морфологический профиль, архитектура каркасов
Короткий адрес: https://sciup.org/149129320
IDR: 149129320 | УДК: 551.734.1 | DOI: 10.19110/2221-1381-2018-6-35-42
Structure of upper Lochkovian (Lower Devonian) Lop'yu Kyrta organic buildup (Ilych river, Northern Urals)
Studied rocks from Lop'yu Kyrta rock at the Ilych River (Northern Urals) are represented mainly by biohermal limestones, lime sandstones, gravelite-sandstones and breccias. Among Biohermal limestones stromatolitic ones are predominate, coral, Fistulella -microbial, and stromatoporoid-microbial ones are rarer. The comparison and correlation of transect series allow us to establish the landscape-morphological zonation of Lop'yu Kyrta buildup, an outer slope and a reef-flat are distinguished. The reef-flat was differentiated into the reef-crest, gravel-sand shoals, a reef core and a back-reef. In general, the stromatolite structure of framework and the characteristic reef zonation can attribute Lop'yu Kyrta to a specific type of buildups - a stromatolite reef. It was established that the Lop'yu Kyrta stromatolite reef had much in common with the modern microbial Highburne Cay reef at the Great Baham Bank. Stratigraphically and geographically similar Upper Silurian stromatolite reefs from the Northern Urals and Alaska are considerably different by their frameworks architecture from Lop'yu Kyrta buildup.
Текст научной статьи Строение верхнелохковской (нижний девон) органогенной постройки Лопъю-Кырта (р. Илыч, Северный Урал)
В палеозойской истории карбонатонакопления на Печорском Урале выделены несколько этапов развития органогенных сооружений, среди которых сандбий-ско-раннеэмский имел наиболее сложную историю [1]. Одним из удивительнейших эпизодов на данной территории является раннедевонский, характеризующийся пиком рифообразования с большим разнообразием ископаемых сообществ [1, 8,10] в прагиене-раннем эмсе, и который происходил на фоне позднесилурийско-раннедевонского так называемого « протяженного рифового кризиса » [13], регистрируемого по всему миру. Однако, первые девонские рифогенные образования появились ещё в позднелохковское время. Наиболее известным примером является Усинский рифовый массив на р. Б. Уса в районе устья р. Нияю [1, 8], но сильная доломитизация пород дает мало данных о его строении.
Другим примером верхнелохковских органогенных построек является разрез Лопью-Кырта на р. Илыч (рис. 1, а). Краткие сведения о строении этой постройки имеются в работах А. И. Антошкиной [1], фаунистическая характеристика дана В. С. Цыганко [8], интересные наблюдения над морфологией колоний кораллов сделаны И. И. Пархетой с соавторами [5]. Автором она изучалась во время полевых работ 2015 г. Изучение разреза сопровождалось прохождением ряда профилей (рис. 1, b). Определения условий залегания (по ватерпасам, залеганию кораллов и ориентировке биокласто-вого материала) показали, что на большей части разреза породы падают на юго-запад под углом 30°. Хорошая обнаженность и слабая степень эпигенетических изменений пород позволяют достаточно детально изучить строение и структуру органогенной постройки Лопъю-Кырта, провести палеогеографический анализ. Целью работы является анализ полученных данных и обоснование генезиса этого органогенного тела.
1. Строение разреза
Разрез Лопъю-Кырта (обн. 144; по [4]) вскрывается на правом берегу р. Илыч, ниже устья р. Лопъю (рис. 1, а). Обнажение представляет собой длинные выходы невысоких скал, преимущественно находящихся в лесу, на протяжении 0.3 км.
1.1. Основные типы пород
Органогенная постройка сложена биогермными известняками, но не редки здесь известняковые песчаники, гравелито-песчаники и брекчии.
Биогермные породы в основном представлены строматолитоподобными и строматолитовыми разностями (рис. 2, а). Из-за отсутствия хорошо отмытых по- 35
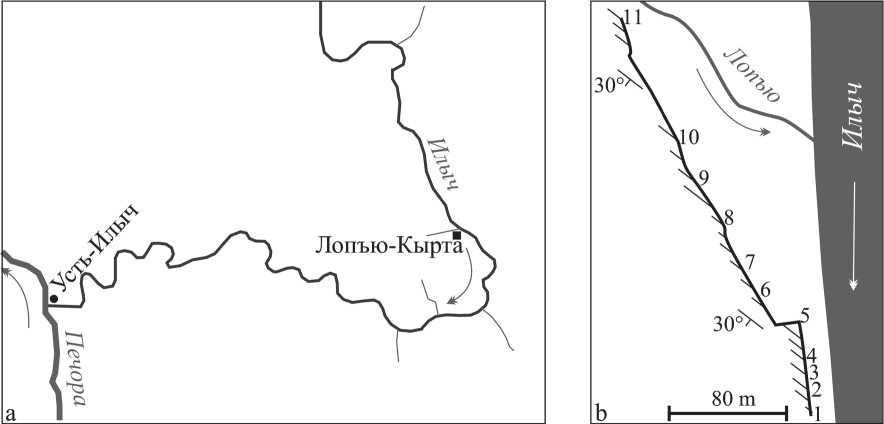
Рис. 1. а. Расположение разреза Лопъю-Кырта. b. Карта-схема обнажения Лопъю-Кырта с расположением пройденных профилей
Fig. 1. а. Location of the Lop’yu Kyrta section. b. A schematic map of the Lop’yu Kyrta outcrop with transects location
верхностей сложно определить размеры и морфологию строматолитов; только в шлифах отмечаются мелкие куполовидные (до 1.0 см в диаметре) и столбчатые (0.4—10 мм шириной и 0.8—20 мм высотой) морфоти-пы. Не исключается, что эти сложности здесь обусловлены большими размерами строматолитов. Так, например, в строении лудловского рифа Дрейк (Аляска, террейн Александра) описаны строматолитовые маты, достигающие в длину более 30 м [1]. Микроскопически строматолиты спонгиостроматовые, в них наблюдается чередование тонких более светлых и более темных слой-ков (0.2—1.0 мм), сложенных тонкозернистым кальцитом (0.007—0.036 мм), в которых иногда отмечаются теневые нитчатые образования 0.02—0.06 мм диаметром и 0.4—0.6 мм длиной (рис. 2, b). Поростроматовые структуры (рис. 2, c) отмечаются участками и представлены скоплениями (0.2—4.0 мм) цианобактерий, напоминающих Renalcis (?). Пространство между строматолитами (1.0—2.0 мм шириной и 0.4—1.0 см длиной) в основном залечено инкрустационным радиально-фибровым (корки 0.1—0.2 мм) и друзовым спаритовым кальцитом. Незначительными компонентами (до 5 %) этих пород является биокластовый и литокластовый материал разного размера (от 0.1 мм до 0.1 м).
Известняки биогермные фистулеллово-микроби-альные и микробиально-строматопоровые распространены участками, слагают пятна от 0.1 до 0.5 м в диаметре и выделяются по обилию цементирующей массы (рис. 2, d). Каркас здесь, как правило, сложен колониями метазой, инкрустированных цианобактериями (до 2.0 мм). Пространство между ними частично заполнено литокластовым и биокластовым материалом, но в основном — радиально-фибровым инкрустационным и друзовым спаритовым кальцитовыми цементами (рис. 2, e).
Интерпретация . Строматолитовое строение пород является результатом жизнедеятельности микробиальных матов со сложной биогеохимической и бактериальной зональностью [3]. Микро-тонкозернистый кальцит в них появился, по всей видимости, при тафономических преобразованиях цианобактериальных пленок и матов, [16], свидетельством чему служат тени их нитей.
Редкие биокласты среди тонкослоистого известняка микробиального происхождения являются результа- 36
том улавливания цианобактериальными матами мелких частиц карбонатного материала песчаной размерности. Последующая минерализация экстраполимерной субстанции (гликокаликса), слагающего большую часть мата, приводила к появлению твердых участков, на которых после гибели цианобактериального сообщества селились многоклеточные организмы. Об этом свидетельствует пятнистый характер распространения биогермных пород с остатками скелетов многоклеточных организмов. Их небольшие поселения мешали развиваться в ближайших окрестностях цианобактериальному сообществу, вероятно, из-за затенения поверхности субстрата, приводившего к локальному уменьшению жизнедеятельности фототрофных бактерий.
Известняковые песчаники и гравелито-песчаники (рис. 3, а) слагают слои среди биогермных разностей и имеют мощность от первых миллиметров до 2.0 м. Слоистая текстура обусловлена ориентировкой зерен, реже — вариациями их размеров. Литокласты (0.2— 0.5 мм), как правило, представлены обломками пелитоморфных известняков. В редких случаях, когда размеры дебриса достигают 2.0—5.0 мм, в их составе отмечают -ся фрагменты среднезернистых литокластовых известняковых песчаников, сгустковых и строматолитовых микробиальных известняков и фистулелловых известняков. Органогенный материал представлен обломками члеников криноидей, раковин брахиопод и остракод. Реже отмечаются фрагменты щитков трилобитов, раковин гастропод, веточек фистулелл, колоний красных водорослей, цианобактерий, табулят (аулопориды, фаво-зитиды), ругоз и строматопороидей. Цемент — друзовый спаритовый кальцит; крайне редко отмечаются радиально-фибровые оторочки или пелитоморфный кальцит.
Интерпретация . Эти известняки являются результатом осадконакопления в активной среде, о чем свидетельствуют кристаллические цементы и преимущественная сортировка зерен. Иногда проявляющаяся градационная слоистость также указывает на влияние периодических течений (вероятно, приливно-отливных). Широкое развитие обломков пелитоморфных известняков свидетельствует либо о размыве этих пород, либо о сильной микритизации обломков с образованием пе-лоидов-багамитов [12], что указывает на крайне мелко-
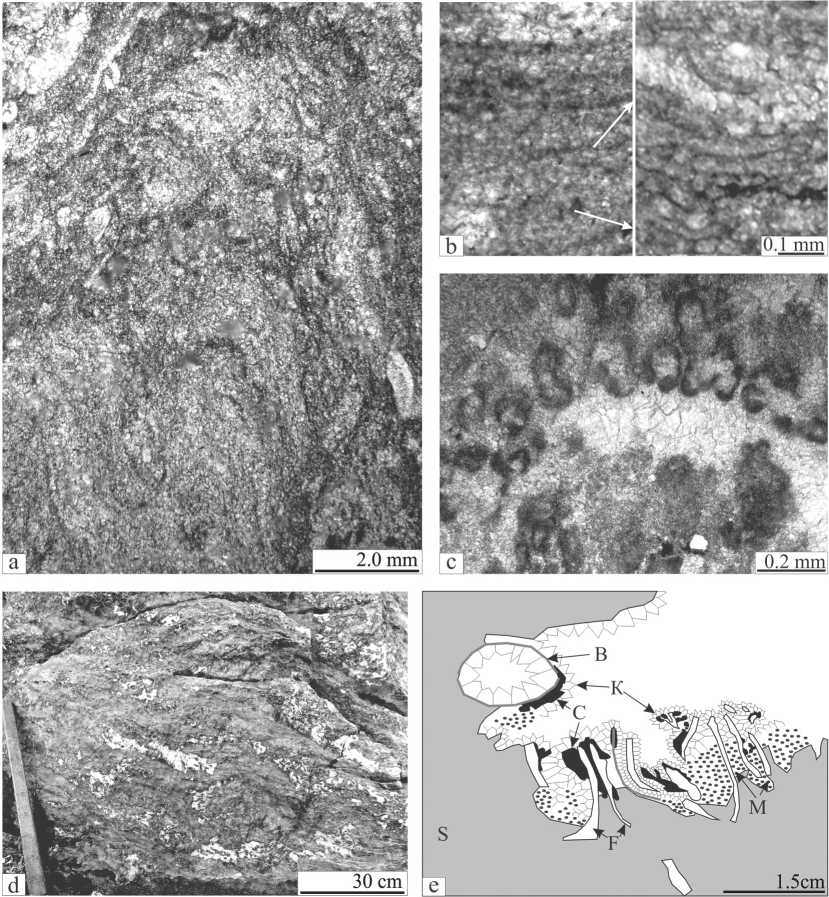
Рис. 2. Биогермные породы. а. Известняк строматолитовый перекристаллизованный. Отмечается теневая неясно-слоистая текстура с мелкими «прилипшими» биокластами. Шлиф П-Ил144/29. b. Теневые нитчатые микроструктуры в строматолите (слева; шлиф П-Ил144/6) и нити цианобактерий в обломке строматолита в склоновых турбидитах валганской свиты (справа; шлиф П-Ил113/1). Стрелки указывают на близкую толщину нитей. с. Колонии цианобактерий. напоминающие Renalcis (?). Шлиф П-Ил144/13. d. Участки с обилием цемента (светлое) во вмещающих микробиальных строматолитовых перекристаллизованных известняках. Фотография хорошо отмытой глыбы у уреза воды напротив профиля 7. е. Зарисовка взаимоотношения строматолитовых и фистулелловых разностей. S — биогермные строматолитовые известняки; F — Fistulella ; М — межкаркасное заполнение мелким био- и литокластовым материалом; C — инкрустации цианобактерий; К — цементные корки; B — фрагменты и раковины брахиопод. Шлиф П-Ил144/46
Fig. 2. Biohermal rocks. a. A recrystallized stromatolitic limestone. Note a relict of unclear-laminated structure with small «adherent» bioclasts. Thin section П-Ил144/29. b. Relicts of filamentous microtextures into stromatolite (at the left; thin-section П-Ил144/6) and cyanobacterial filaments in a stromatolite clast from the Valganskaya Fm turbidites (at the right; П-Ил113/1). Arrows show an approximately thickness of filaments. c. Cyanobacterial colonies which are similar to Renalcis (?). Thin section П-Ил144/13. d. Areas with abundant cements (light) in hosting recristallized microbial stromatolite limestones. The photo shows a well-washed large boulder near the water-line of the Ilych River opposite transect 7. e. The sketch of a stromatolite and Fistulella relationship. S — stromatolite boundstones; F — Fistulella , M — an inter-framework filling with small bio- and lithoclasts; C — cyanobacterial encrustations; K — crusts of cements; B — fragments and shells of brachiopods. Thin section П-Ил144/46
водные и прогреваемые морские воды. Биокластовый материал имеет местное происхождение, так как не несет следов окатанности.
Известняковые брекчии слагают линзовидные прослои и участки до 0.8 м в диаметре (рис. 3, b). Это серые пятнистые породы, состоящие из обломков пород (70— 80 %) и доломитового матрикса (20—30 %). Обломки (от 1.0 мм до 0.1 м) не окатаны, имеют угловатые очертания со слабоокругленными и вогнутыми краями. Часто в них встречаются впадинки и более мелкие карманы (рис. 3, c). Состав обломков, хоть и гетерогенный, но включает те типы пород, которые окружают брекчии. В основном это фрагменты известняков биогер-мных строматолитоподобных (17—76 %) и/или био-ли- токластовых песчаников (12—44 %). Реже отмечаются обломки известняков сгустковых (0—14 %), цианобактериальных (7—20 %), фистулелловых (0—13 %) и перекристаллизованных или цементных корок (7—13 %). Матрикс микро-тонкозернистый (0.007—0.08 мм) глинистый доломит (рис 3, d) с незначительным содержанием биокластов (0.2—1.1 мм), представленных обломками члеников криноидей, а также целыми раковинками мелких брахиопод. Важно отметить, что во вмещающих породах на границах с брекчиями часты структуры, похожие на Palaeomicrocodium.
Интерпретация . Эти брекчии представляют собой продукт разрушения органогенного сооружения и накопления обломков в понижениях рельефа. Линзовидные 37
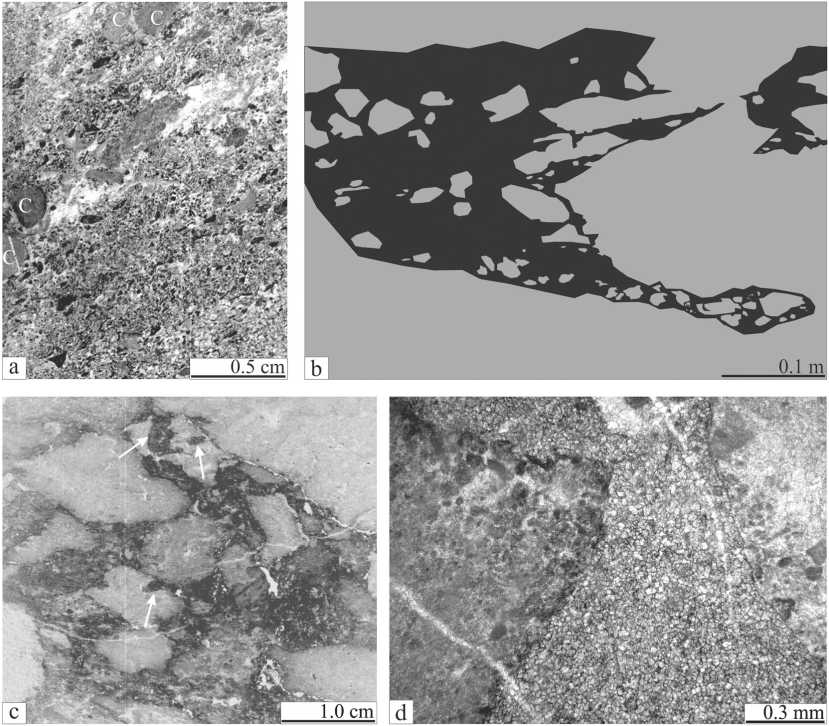
Рис. 3. Обломочные породы. а. Известняковый песчаник с тонким прослоем известнякового гравелито-песчаника. С — членики криноидей. Шлиф П-Ил144/22. b. Положение известняковых брекчий в разрезе Лопъю-Кырта. Полевая зарисовка с указанием наиболее крупных обломков. Профиль 7, верхняя часть. с. Известняковая брекчия с глинисто-доломитовым цементом. Стрелки указывают на карманы, интерпретируемые как следы растворения. Шлиф П-Ил144/11. d. Глинисто-доломитовый матрикс между обломками. Шлиф П-Ил144/11
Fig 3. Clastic rocks. a. The calcareous sandstone with thin layer of the calcareous gravelite-sandstone. C — crinoids ossicles. Thin section П-Ил144/22. b. Locations of calcareous breccias into the Lop’yu Kyrta section. A field sketch which marked largest clasts. The upper part of the transect 7. c. Calcareous breccias with clayey-dolomite matrix. Arrows point to small cavities that interpreted as traces of dissolution. Thin section П-Ил144/11. d. A clayey-dolomite matrix between clasts. Thin section П-Ил144/11
прослои могут являть собой поперечные и косые сечения системы каналов. Однако, некоторые участки брекчий могут иметь карстовое происхождение. Об этом свидетельствуют структуры Palaeomicrocodium во вмещающих породах (результат жизнедеятельности грибов и актиномицетов, развивавшихся в условиях субаэрального воздействия [2]), а также впадинки и карманы с рваными краями на обломках, интерпретируемые как следы растворения.
1.2. Особенности строения постройки
При описании разрезов профилей (рис. 4, а) внимание уделялось строению пород, распространению основных групп органических остатков и раннедиагенетических цементов.
Самые нижние «слои» органогенной постройки Лопъю-Кырта описаны на северной оконечности скалы в профиле 11 . Это светло-серые массивные биогер-мные известняки преимущественно строматолитовые, со скоплениями строматопороидей и фистулелл. В нижней части породы брекчированы. Видимая мощность пород в профиле — 7.5 м.
На расстоянии около 120 м на Ю-ЮВ отмечаются крупноглыбовые развалы биогермных известняков. Между профилями 10 и 11 не обнажено около 12.5 м по мощности. Профиль 10 вскрывает светло-серые биогер-мные пятнистые известняки, строматопороидно-стро-матолитовые в нижней части и фистулеллово-коралло-во-микробиальные — в верхних 1.5 м. Видимая мощ- 38
ность вскрытых пород (включая не обнаженные интервалы) — 9.5 м.
Профиль 9 расположен в 25 м к ЮЮВ от профиля 10, при этом нижние 2.1 м профиля 9 соответствуют верхней части профиля 10. Биогермные стромато-пороидно-строматолитовые известняки сменяются по латерали известняками темно-серыми биогермными микробиальными и известняковыми гравелито-песча-никами мощностью 2.0 м в нижней части профиля 9. В остальном здесь вскрыты массивные пятнистые темно-серые биогермные строматолитовые известняки с участками строматопорово-строматолитовых (более редки) и фистулеллово-строматолитовых (более часты) разностей. Видимая мощность — 11.8 м.
Профиль 8 в 20 м к ЮЮВ в небольшом выходе био-гермных известняков, видимой мощностью 2.5 м, соответствует средней части профиля 9. Здесь вскрыты в основном строматолитовые известняки. Участками развиты скопления колоний кораллов табулят и фисту-леллы. К гидроидным разностям приурочено развитие раннедиагенетических цементов.
Профиль 7 находится в 20 м к ЮЮВ от предыдущего. Здесь вскрыты слои, наращивающие биогермные отложения профиля 9. Нижний 1.0 м профиля 7 слагают известняковые био-литокластовые песчаники, а верхнюю часть — биогермные известняки, характеризующиеся, кроме развития строматолитов, обилием участков с табулятами и фистулеллами с большим вкла- дом раннедиагенетической цементации. В верхней части отмечаются небольшие (до 0.2 м) участки известняковых брекчий. Видимая мощность — 5.5 м.
Профиль 6 расположен в 20 м к ЮЮВ, и его нижняя часть соответствует верхнему 1.0 м интервалу профиля 7. Здесь также отмечается фациальное замещение биогермных разностей известняковыми гравелито-песчаниками. Выше залегают биогермные известняки с большим количеством фистулеллово-микробиаль-ных, коралловых разностей и раннедиагенетических цементов. В известняковых песчаниках широко развиты участки известняковых брекчий. Видимая мощность пород — 9.0 м.
Профиль 5 (в 30 м к ЮВ) своей верхней частью соответствует профилю 6. Здесь отмечается сложное че редование известняков биогермных, известняковых песчаников гравелито-песчаников и брекчий. В био-гермных известняках, по сравнению с предыдущим профилем, уменьшается роль метазойных организмов; они хоть и разнообразны, но встречаются преимущественно как обломки. Из-за широкого развития меха-ногенных осадков уменьшена роль и строматолитов. Преобладают обломки члеников криноидей и раковин брахиопод. Роль раннедиагенетических цементов незначительна. Видимая мощность пород — 9.0 м.
Профиль 4 описан в 20 м к Ю. Нижние 2.0 м его соответствуют верхней части профиля 5 и показывают латеральное замещение сложного чередования пород 1.5-метровым слоем известняковых гравелито-пес-чаников. Верхние 11 м профиля 4 сложены в основном
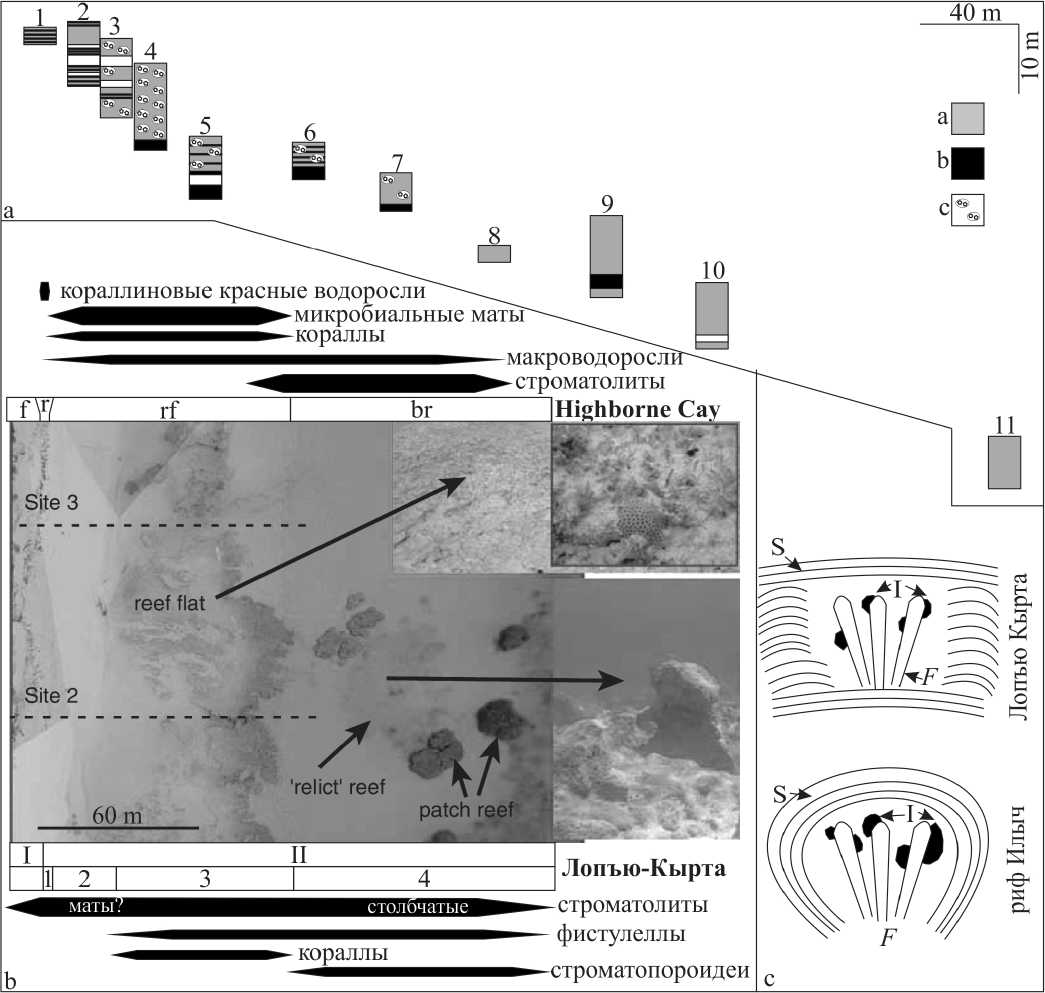
Рис. 4. Строение разреза Лопъю-Кырта. a. Вертикальные и латеральные взаимоотношения профилей в скале Лопъю-Кырта. а — биогермные известняки; b — известняковые гравелито-песчаники; с — известняковые брекчии. b. Сравнение ландшафтно-морфологической зональности и распространения организмов верхнелохковского строматолитового рифа Лопъю-Кырта и вероятного современного аналога. Highborne Cay : f — фронт рифа; с — гребень рифа; rf — рифовое плато; b — зарифовая область. Аэрофотоснимок рифа Highborne Cay [по: 17]. Строматолитовый риф Лопъю-Кырта : I — предриф-флет; II — рифовое плато (1 — гребень рифа; 2 — песчано-гравийные отмели; 3 — центральная зона; 4 — тыловая зона). с. Схема фистулел-лово-микробиальных взаимоотношений в верхнелудловском рифе Илыч и верхнелохковском строматолитовом рифе Лопъю-Кырта. F — Fistulella , I — инкрустаторы, S — строматолитовые корки.
Fig. 4. A composition of the Lop’yu Kyrta section. a. Vertical and lateral relationships of transects. a — biohermal boundstounes; b — calcareous gravelite-sandstones; c — calcareous breccias. b. The comparison of the landscape-morphological zonation and organisms distribution of Upper Lochkovian Lop’yu Kyrta stromatoreef and the modern Highburne Cay microbial reef. Highburne Cay : f — reef front; r — reef crest; rf — reef-flat; br — back-reef area. The aerial photograph of the Highborn Cay [After: 17]. The Lop’yu Kyrta : I — forereef-flat; II — reef-flat (1 — algal ridge; 2 — gravel-sand shoals; 3 — central part; 4 — back-reef). c. A scheme of the Fistulella -microbial relationships in the Upper Ludlovian reef on the Ilych River and in Upper Lochkovian Lop’yu Kyrta stromatoreef. F — Fistulella , I — encrusters; S — stromatolite crusts
биогермными строматолитовыми породами с незначительным количеством фистулелл. Табуляты в основном представлены обломками колоний. Широко развитие участков известняковых брекчий. Видимая мощность — 12.5 м.
Породы в профиле 3, в 12 метрах южнее, большей частью соответствуют профилю 4 за исключением верхних 3.5 м и показывают в целом похожее строение. Изменения заключаются в уменьшении количества известняковых брекчий. Известняковые песчаники развиты в виде незначительных прослоев (от первых мм до 3—4 см). Видимая мощность — 11.4 м.
Профили 1 и 2 расположены в 25 и 11 м к Ю соответственно. Они сложены, в целом, известняками строматолитовыми с прослоями (до первых сантиметров мощностью) известняковых песчаников. Известняковые брекчии не отмечены. Видимая мощность разреза в профиле 2 — 9.3 м, а в профиле 1 — 2.5 м.
2. Обсуждение материала
В строении рифогенных образований выделяются вертикальные (экологическая сукцессия) и латеральные (ландшафтно-морфологические) изменения. Вариации пород, изученных по профилям в разрезе Лопъю-Кырта, могут отражать как сукцессионные, так и морфологические изменения. Однако, в некоторых соседних профилях, частично вскрывающих стратиграфически одни и те же уровни, отмечается четкая латеральная смена фаций. Поэтому, вероятно, в данном случае будет правильно интерпретировать их с палео-ландшафтной позиции.
2.1. Палеоландшафтный профиль
Согласно [7], поперечный профиль рифа представлен внешним склоном (I), рифовым плато (II) и внутренним (лагунным) склоном рифа. В изученном разрезе можно выделить первые два элемента (I и II).
I. Внешний склон органогенной постройки Лопъю-Кырта выражен слабо. Здесь можно условно выделить зону предриф-флета [7]. К этой зоне можно предположительно отнести отложения, изученные в профилях 1 и 2. Хотя для этой зоны обычно нехарактерно накопление обломочного материала, в разрезе Лопъю-Кырта отмечаются тонкие прослои с обильными карбонатными песчинками, которые, видимо, были уловлены цианобактериальными матами и в дальнейшем вошли в состав минерализованной экстраполимерной субстанции.
II. Риф-флет или рифовое плато представляет собой субгоризонтальную поверхность. Нами в его строении выделяются: (1) гребень, (2) песчано-гравийные отмели, (3) центральная зона рифового плато, и (4) тыловая зона рифового плато.
1. Гребень рифа — часть рифа, находящаяся в полосе бурунов [7]. К этой зоне, вероятно, можно отнести отложения, изученные в профилях 3 и 4, характеризующиеся развитием биогермных строматолитовых известняков. Обилие участков с известняковыми брекчиями, по всей видимости, свидетельствует о частом выводе его выше уровня моря, хотя часть этих участков вполне могут быть и отложениями прибойных каналов.
2. Песчано-гравийные отмели рифового плато широко развиты как в современных, так и в ископаемых рифах [7]. Они диагностированы в нижнепалеозойских рифах Урала [1]. В разрезе Лопъю-Кырта эта зона изучена в профилях 5 и 6, в которых выделяется по широ-40
3. Центральная зона рифового плато изучена в профилях 7-9 и характеризуется, как и в ископаемых рифах Урала [1], широким развитием биогермных разностей, наибольшим разнообразием организмов и обилием раннедиагенетических цементов. Однако, несмотря на разнообразие здесь каркасостроителей, в количественном отношении преобладают биогермные строма-толитовые разности.
4. Тыловая зона рифового плато изучена в профилях 10 и 11. Здесь снова начинают преобладать биогер-мные строматолитовые разности с небольшими участками биогермных разностей другого состава (пэтч-рифы). Эта зона, по всей видимости, уже является переходной к зарифовым лагунным отложениям.
2.2. Тип постройки Лопъю-Кырта
2.3. Сравнительный анализ
кому развитию известняковых песчаников и гравелито-песчаников.
Одним из удобных принципов классификации органогенных сооружений является их разделение на мелководные рифы и холмы, развивавшиеся преимущественно в спокойных водах [14]. Для последних, как правило, характерны бактериальные образования. С этих позиций, разрез Лопъю-Кырта представляет уникальную возможность увидеть органогенное сооружение с палеоландшафтно-морфологическими характеристиками рифов, но со структурой сообщества, больше характерного для микробиальных холмов. Преобладающее строматолитовое строение, мелковод-ность и присутствие рифовой зональности позволяет отнести рифогенное образование Лопъю-Кырта к специфическому типу — строматолитовому рифу [6].
Обычно одной из важнейших характеристик ископаемых рифов считают их размеры. Так, нижнепалеозойские рифы имеют мощность от 147 м (верхнеордовикский риф Большая Косью [9]) до 1200 м (пражско-нижнеэмский риф на р. Лемва [1]), но в среднем — 300—500 м. Другой критерий обычно слабее освещен в литературе — это ширина рифа. Так, например, по схематическим картам [1] можно предположить ширину нижнепалеозойских рифов в первые километры. По сравнению с ними строматолитовый риф Лопъю-Кырта относительно небольшой: видимая мощность оценивается в 65—70 м. По одному пересечению органогенной постройки нет возможности установить истинную ширину строматолитового рифа. Однако, даже если предположить, что пересечение было сделано перпендикулярно палеоландшафтным зонам, то ширину строматолитового рифа можно оценить примерно в 270 м. Если же учесть преимущественно субмеридиональное простирание фациальных поясов на севере Урала, то ширина постройки Лопъю-Кырта может быть оценена не более чем в 130 м.
Современные рифы преимущественно метазой-ные, за исключением единичных примеров, известных на островной цепи Экзума Кэйс (Exuma Cays) в центральной части Большой Багамской банки [11]. Здесь риф Хайбёрн Кэй (Highborne Cay) имеет ширину до первых сотен метров (рис. 4, b) и, в целом, сходное со строматолитовым рифом Лопъю-Кырта строение. В обоих случаях постройки сложены микробиальными образованиями. Для Хайбёрн Кэй характерно развитие микробных матов на рифовом плато и столбчатых строматолитов в зарифовой области. В строении Лопъю-
Кырта ясно выраженное строматолитовое строение имеет тыловая часть рифового плато, а для центральной части, предположительно, также характерны микробиальные маты. Важные различия касаются строения гребня рифа и пэтч-рифов. Рифовый гребень (reef crest) Хайбёрн Кэй сложен красными кораллиновыми водорослями, а в постройке Лопъю-Кырта этот элемент представлен микробиальными образованиями. Пэтч-рифы Хайбёрн Кэй сложены исключительно строматолитами, а в позднелохковском аналоге кроме строматолитов в них развиты перекристаллизованные цено-стеумы строматопороидей. Приуроченность колоний кораллов в обоих случаях ограничена исключительно рифовым плато, а распространение проблематичных гидроидов Fistulella в строматолитовом рифе Лопъю-Кырта совпадает с распространением макроводорослей на рифе Хайбёрн Кэй (см. рис. 4, b). Макроводоросли в Хайбёрн Кэй разнообразны [15], широко развиты на рифовом плато и колонизируют вершины строматолитовых пэтч-рифов. По всей видимости, фистулеллы играли функционально ту же роль, что и макроводоросли — колонизаторы твердых поверхностей.
Этот факт выявляет различия в архитектуре мета-зойно-микробиальных каркасов позднего лохкова и луд-лова (верхний силур). Верхнесилурийские строматоли-товые рифы Аляски стали широко известны всему миру благодаря исследованиям К. Соджа и А. И. Антошкиной [17, 18]. Однако в их строении [1] не удалось «опознать» зональности, подобной Лопъю-Кырты. В строении же строматолитовых известняков лудловских рифов формации Хекета неоднократно подчеркивается, что ме-тазойные организмы часто « ...являются как бы ядрами концентров внутри строматолитовых матов» [1, с. 104]. Архитектуру основных типов каркасов верхнесилурийского рифа Илыч можно представить следующим образом (рис. 4, с): а) основу каркаса составляют фистулел-лы, на которые нарастают губки-афросальпингиды и различные цианобактерии; б) каркас обволакивается толстой (от первых миллиметров до 1.0 см) коркой строматолитовых карбонатов; в) межкаркасные полости заполняются биокластовым материалом, либо (чаще) залечиваются различными типами цементов. В таком случае, метазойные организмы и цианобактерии можно отнести к «гильдии конструкторов» [12], тогда как стро-матолитовые корки — явно к «гильдии склеивателей».
Иная картина складывается в архитектуре каркасов строматолитового рифа Лопъю-Кырта. Здесь строматолиты большей частью сами являются и конструкторами, и склеивателями. Фистулелл, в лучшем случае, можно отнести к «гильдии улавливателей», либо к «гильдии жителей» (Dweller Guild). Фистулеллы, подобно современным морским травам на Хайбёрн Кэй, колонизировали поверхность микробиальных матов, в том числе и на поверхности строматолитов, создавая затененные участки поверхности рифа и препятствуя дальнейшему развитию фототрофных цианобактериальных сообществ. Конечно, в итоге микробные сообщества снова одерживали верх: либо после смерти ме-тазойных организмов, либо в силу более быстрого роста. Как результат — образование множества участков с обилием цемента на месте бывших фистулелловых поселений (рис. 2, d, e).
Причины таких различий функциональной роли фистулелл пока сложно объяснить. Сейчас можно лишь констатировать факт, что основная каркасообразующая роль в позднем силуре принадлежала метазоям, а в начале девона (поздний лохков) роль метазой (стро-матолитовый риф Лопъю-Кырта) отошла на «вторые планы», а основную каркасообразующую роль взяли на себя микробные сообщества.
Заключение
Изучение верхнелохковской (нижний девон) органогенной постройки Лопъю-Кырта показало, что:
-
1. В строении постройки играют важную роль стро-матолитовые образования, реже метазойные разности, а также известняковые песчаники, гравелито-песчани-ки и брекчии. На основании преобладания строматолитовых разностей в строении Лопъю-Кырты постройка отнесена к специфическому для палеозоя типу строматолитовых рифов.
-
2. В строматолитовом рифе выделено два палеолан-дшафтных элемента: внешний склон и рифовое плато. В составе последнего элемента выделяются 4 зоны: гребень рифа, песчано-гравийные отмели, центральная и тыловая зона рифового плато.
-
3. Изученная постройка значительно уступает в размерах другим нижнепалеозойским рифам. Видимая мощность Лопъю-Кырты определяется 65—70 м, в сравнении со средней мощностью верхнеордовикско-ниж-недевонских рифов в 300—500 м. Поперечные размеры строматолитового рифа оцениваются между 130 и 270 м, тогда как для рифов они составляют первые километры.
-
4. Вероятным современным аналогом Лопъю-Кырты может быть преимущественно микробиальный риф Хайбёрн Кэй, расположенный в центральной части Большой Багамской банки. Они сходны по размерам, морфологическим элементам и распространению палеосообществ. Удивительна одинаковая приуроченность проблематичных гидроидов Fistulella в стромато-литовом рифе Лопъю Кырта и макроводорослей на рифе Хайбёрн Кэй. Это указывает на фунционально одинаковую их роль.
-
5. С другой стороны, казалось бы, стратиграфически и географически более близкие верхнесилурийские строматолитовые рифы Северного Урала и Аляски отличаются от Лопъю-Кырты архитектурой каркаса и ролью метазойных и бактериальных сообществ. Основная каркасообразующая роль в позднем силуре принадлежала метазоям, тогда как строматолитовые образования играли роль склеивателей. Однако, в позднелох-ковской постройке Лопъю-Кырта функциональная роль метазой отошла на «вторые планы», а основную каркасообразующую роль взяли на себя бактериальные сообщества.
Работа проводилась в рамках Госпрограммы № АААА-А17-117121270034-3 и при частичной финансовой поддержке Программы фундаментальных исследований УрО РАН, проект № 18-5-5-31.
Список литературы Строение верхнелохковской (нижний девон) органогенной постройки Лопъю-Кырта (р. Илыч, Северный Урал)
- Антошкина А. И. Рифообразование в палеозое (север Урала и сопредельные области). Екатеринбург: УрО РАН, 2003. 303 с.
- Антошкина А. И. Палеомикрокодии: новый взгляд на их генезис//Палеонтологический журнал, 2014. № 4. С. 15-31.
- Бактериальная палеонтология/Под ред. А. Ю. Розанова. М.: ПИН РАН, 2002. 188 с.
- Варсанофьева В. А. Геологическое строение территории Печоро-Илычского государственного заповедника//Тр. Печоро-Илычского заповедника, 1940. Вып. 1. С. 5-214.
- Пархета И. И., Гатовский Ю. А., Пономаренко Е. С., Архипова Е. В. Анализ условий обитания палеозойских биоценозов Северного Урала в районе бассейна р. Илыч//Вестник Международного университета природы, общества и человека «Дубна»/Сер. «Естественные и инженерные науки», 2016. № 3 (35). С. 36-43.
