Структура и динамика микробных комплексов филлосферы и ризосферы хвойных видов лесных биогеоценозов Приенисейской Сибири (на примере Предивинского лесхоза)
")
Автор: Сорокин Н.Д., Афанасова Е.Н.
Журнал: Вестник Красноярского государственного аграрного университета @vestnik-kgau
Рубрика: Биологические науки
Статья в выпуске: 8, 2016 года.
Бесплатный доступ
Объектом исследования являлась микро-флора хвои и ризосферы ели сибирской (Picea obovata), сосны обыкновенной (Pinus sylvestris), сосны кедровой (Pinus sibirica), произрастаю-щих в лесных биогеоценозах Предивинского лесхоза (Большемуртинский район). Возраст деревьев 30-35 лет. Целью работы было изу-чить сопряженную динамику формирования структуры микробных комплексов филлосфе-ры и ризосферы различных видов хвойных в лесных биогеоценозах Сибири. При исследова-нии динамики формирования эпифитной мик-рофлоры хвои ели сибирской отмечено макси-мальное развитие неспоровых форм бактерий в сентябре, что связано с прекращением ак-тивной вегетации растений и значительным снижением их фитонцидной активности. Ди-намика мицелиальных грибов, находящихся в минимуме, не зависит от температурных ус-ловий и условий влажности в течение вегета-ционного периода. Формирование эпифитной микрофлоры хвои сосны обыкновенной имеет отличие от динамики ели. Здесь регистриру-ется максимум развития неспоровых микроор-ганизмов в июне и сентябре, что связано с не-высокой фитонцидной активностью сосны в эти периоды. При формировании эпифитной микрофлоры хвои сосны обыкновенной следу-ет учитывать негативное воздействие на эпифиты патогенных микроорганизмов, кото-рые проявляют большую антагонистическую активность по отношению к ним из-за мень-шей фитонцидности хвои. При анализе сред-ней за вегетационный период численности ри-зосферных микроорганизмов на диагностиче-ских средах установлено, что максимальное развитие аммонифицирующих бактерий (рост на МПА) выявляется в почве-контроле и под кедром. Показатели численности микроорга-низмов, использующих минеральные источники азота (рост на КАА), имеют наибольшее зна-чение в контроле, а наименьшие под елью. Максимальные коэффициенты микробиологи-ческой минерализации (КАА/МПА) регистриру-ются под сосной. Эти значения численности и коэффициентов минерализации свидетельст-вуют о несбалансированности процессов син-теза-ресинтеза под этими видами древесных насаждений. Максимальные количества олиго-нитрофильных бактерий определяются под кедром, а коэффициенты олиготрофности (Эшби/МПА) имеют наибольшие значения под елью, что является свидетельством недос-татка элементов питания под этими видами деревьев. При сравнительном анализе сум-марной численности микроорганизмов ризо-сферы и филлосферы различных видов де-ревьев проявились следующие тенденции: аб-рисы динамики численностей для всех видов исследуемых деревьев практически совпада-ют в течение вегетационного периода. Это можно объяснить тем, что комплекс «почва- дерево-микроорганизмы ризосферы и филло-сферы» является экосистемой, компоненты которой связаны функционально и энергети-чески. Такая взаимосвязь и взаимозависимость компонентов сохраняют экосистему в со-стоянии гомеостаза.
Микробные комплексы, филлосфера, ризосфера, суммарная числен-ность микроорганизмов (счм), период веге-тации
Короткий адрес: https://sciup.org/14084775
IDR: 14084775 | УДК: 631.41
The structure and dynamics of microbial fillosfery and rhizosphere complexes of coniferous species of forest biogeocenosis of the Yenisey territory Siberia (on the example of Predivinsk forestry)
The object of research was the microflora of needles and rizosfer of a Siberian fir-tree of (Picea obovata), Scotch pine (Pinus sylvestris), a cedar pine (Pinus sibirica) growing in forest biogeocenoses of Predivinsk forestry (Bolshemurtinsky area). The age of trees was 30-35 years. The purpose of the work was to study the interfaced dynamics of formation of structure of microbic complexes of phyllosphere and rizosfer of different types coniferous in forest biogeocenoses of Siberia. In the research of dynamics of formation of epifit microflora of needles of Siberian fir-tree the maximum development of non sporous forms of bacteria in September connected with the termina-tion of active vegetation of plants and considerable decrease in their fitoncid activity was noted. The dynamics of the micelial mushrooms which were in minimum did not depend on temperature and hu-midity conditions during the vegetative period. The formation of epifit microflora of needles of an ordi-nary pine had the difference from the dynamics of a fir-tree. Here the maximum of development of non sporous microorganisms in June and September, i.e. connected with low fitoncid activity of an ordi-nary pine during these periods was registered. When forming epifit microflora of needles of a pine, it was necessary to consider negative impact on epifit of pathogenic microorganisms which show big antagonistic activity in relation to them because of a smaller fitoncid ability of needles. In the analysis of the number of rizosfern microorganisms, average for the vegetative period, on diagnostic environ-ments it was established that the maximum devel-opment of ammonifying bacteria (growth by MPA) comes to light in the soil control and under a cedar. The indicators of number of the microorganisms using mineral sources of nitrogen (growth on CMM), have the greatest value in control, and the smallest under a fir-tree. The maximum coefficients of a microbiological mineralization (CMM/MPA) were registered in a pine. These values of number and coefficients of mineralization testify to the im-balance of processes of synthesis resynthesis un-der these types of wood plantings. The maximum quantities of oligonitrofil bacteria were defined un-der a cedar, and coefficients of an oligotrof (Eshbi/MPA) have the greatest values under a fir-tree that is the evidence of a lack of batteries under these species of trees. In the comparative analysis of total number of microorganism’s rizosfer and phyllospheres of different types of trees were shown the following tendencies: outlines of dynam-ics of the total number for all species of the studied trees practically coincide during the vegetative pe-riod. It can be explained to that the "soils-trees-microorganisms rizosfer and phyllospheres" com-plex is an ecosystem which components are con-nected functionally and energetically. Such interre-lation and interdependence of components keep an ecosystem in the condition of homeostasis.
Текст научной статьи Структура и динамика микробных комплексов филлосферы и ризосферы хвойных видов лесных биогеоценозов Приенисейской Сибири (на примере Предивинского лесхоза)
Введение. Комплексы микроорганизмов, обитающих на поверхности наземных частей растений и в ризосфере, выполняют важную функцию биоредуцентов органических соединений. Образуемые ими простые минеральные соединения ассимилируются растениями, обеспечивая их рост, развитие и продуктивность фитоценоза. Помимо функции биоредуцентов, микроорганизмы филлосферы (эпифиты) и ризосферы являются фиксаторами атмосферного азота, необходимого для роста высших растений. Кроме того, микроорганизмы являются индикаторами состояния растения.
В настоящее время очевидно, что природный комплекс «почва–дерево–микроорганизмы» следует рассматривать как сложную экосистему, в которой ее компоненты находятся в постоянном функциональном взаимодействии и энергетическом обмене питательными элементами и продуктами метаболизма [3]. При этом состояние дерева во многом зависит от структуры, динамики и функциональной активности микробных сообществ филлосферы и ризосферы. Поэтому важно знать, насколько тесно сопряжены и сбалансированы структурно-динамические параметры филлосферы (эпифиты хвои) и ризосферы (микроорганизмы прикорневой почвы).
Цель работы . Изучение сопряженной динамики формирования структуры микробных комплексов филлосферы и ризосферы различных видов хвойных в лесных биогеоценозах Сибири.
Объекты и методы исследований. Объектом исследования являлась микрофлора хвои и ризосферы ели сибирской ( Picea obovata ), сосны обыкновенной ( Pinus sylvestris ), сосны кедровой ( Pinus sibirica ), произрастающих в лесных биогеоценозах Предивинского лесхоза (Боль-шемуртинский район). Возраст деревьев 30– 35 лет.
В лабораторных условиях была изучена эпифитная микрофлора исследуемых образцов. Для ее анализа бралась навеска в 1 грамм хвои, с которой проводился смыв в 100 мл воды. Для выявления грибов были произведены посевы методом Коха (0,1 мл) на плотную среду Чапека. Для роста бактерий использовался картофельный агар (КА), для актиномицетов – крахмало-аммиачный агар (КАА), для олигонитрофилов – среда Эшби.
Анализ микрофлоры ризосферы проводился по методике Е.З. Теппер [4, 5], адаптированной для древесных растений. Корневая система дерева обнажалась на глубине 10–15 см от слоя органогенного горизонта, и стерильными приборами извлекалась ризосферная почва (вместе с корешками 3–4-го порядка), которая закладывалась в стерильные бюксы. Навеску ризосферной почвы в 1 г помещали в колбу со 100 мл стерильной воды. Затем делали посев из разведения 1/10000. Для роста бактерий использовался мясо-пептонный агар (МПА) вместо КА для эпифитов [1]. Суммарная численность микроорганизмов (СЧМ) определялась как сумма КОЕ на МПА (КА), КАА, среде Чапека [2]. Кон-
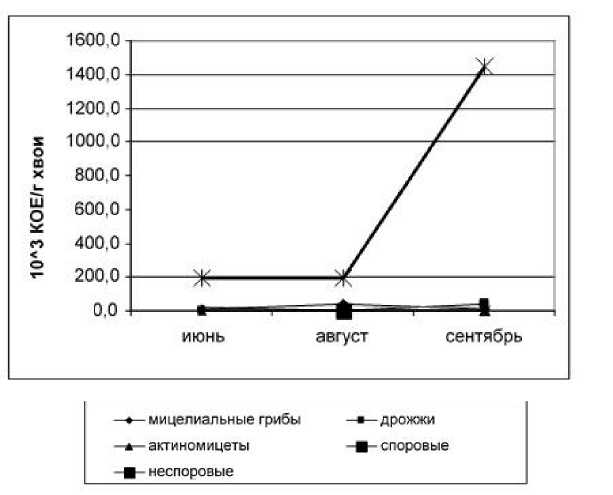
Рис. 1. Динамика формирования эпифитной микрофлоры хвои ели сибирской
тролем служили образцы почвы, взятой в «окнах» насаждений. Образцы хвои и ризосферной почвы отбирались на десяти модельных деревьях каждого вида.
Выросшие микроорганизмы микроскопиро-вались при увеличении х1350. Производился количественный учет колоний мицелиальных грибов, споровых и неспоровых форм бактерий, дрожжей и актиномицетов. С помощью метода Грезерсона определялась грам-принадлежность бактерий.
Результаты исследований и их обсуждение. Поскольку все процессы роста и развития дерева и связанные с ними микробиологические процессы в условиях лесных экосистем Сибири активно проходят в короткий период вегетации, исследования структуры и динамики комплексов эпифитных и ризосферных микроорганизмов, сосны обыкновенной и сосны сибирской проводились в июне, июле, августе и сентябре месяце, а ели сибирской – в июне, августе и сентябре.
При исследовании динамики формирования эпифитной микрофлоры хвои ели сибирской (рис. 1) отмечено максимальное развитие неспоровых форм бактерий в сентябре, что связано с прекращением активной вегетации растений и значительным снижением их фитонцидной активности. Динамика мицелиальных грибов, находящихся в минимуме, практически не зависит от температурных условий и условий влажности в течение вегетационного периода.
Формирование эпифитной микрофлоры хвои сосны обыкновенной (рис. 2) отличается от ели тем, что здесь регистрируется максимум разви- тия неспоровых микроорганизмов в июне и сентябре. Очевидно, это связано с невысокой фитонцидной активностью сосны в эти периоды.
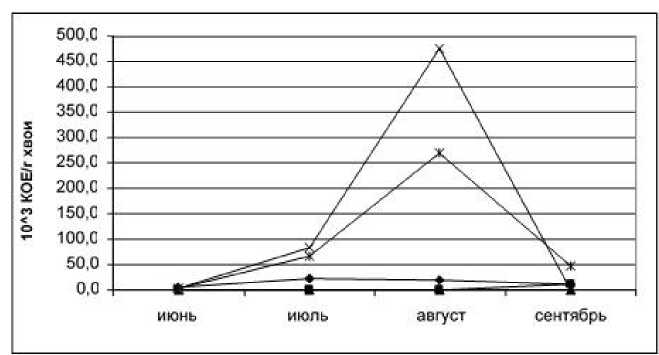
Рис. 2. Динамика формирования эпифитной микрофлоры хвои сосны обыкновенной (обозначения те же)
Кроме того, при формировании эпифитной микрофлоры хвои сосны обыкновенной, вероятно, следует учитывать негативное воздействие на эпифиты патогенных микроорганизмов, которые проявляют большую антагонистическую активность по отношению к ним из-за меньшей фитонцидности хвои.
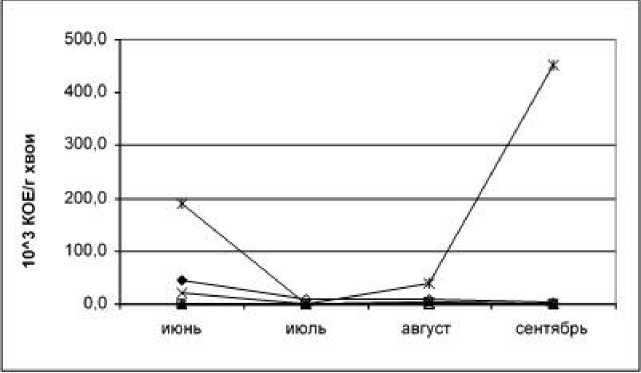
Рис. 3. Динамика формирования эпифитной микрофлоры хвои сосны кедровой (обозначения те же)
При анализе динамики формирования эпифитной микрофлоры хвои сосны сибирской (кедра) выявляется максимум развития споровых и неспоровых бактерий (с преобладанием споровых) в августе. Это можно связать с биологическими особенностями кедра (по сравнению с сосной обыкновенной). Максимальное развитие прокариот на хвое кедра в августе связано со значительными выделениями листо- вой поверхности и незначительной фитонцидной активностью в этот период (рис. 3).
Во всех случаях динамика мицелиальных грибов не имеет достоверных экстремумов развития, что объясняется меньшей зависимостью данной таксономической группы от гидротермических условий развития и фитонцидной активности дерева.
Исследования количественного состава мик- ределения суммарной численности (СЧМ) микрофлоры в ризосфере проводились путем оп- роорганизмов (табл. 1).
Таблица 1
Суммарная численность микроорганизмов ризосферы под различными видами деревьев в течение вегетационного сезона, тыс. кОе/ г воздушно-сухой почвы
|
Вид хвойных |
Июнь |
Июль |
Август |
Сентябрь |
|
Сосна |
4859 |
2447 |
2537 |
4942 |
|
Ель |
2986 |
2485 |
2676 |
3875 |
|
Кедр |
3237 |
3632 |
4870 |
2950 |
|
Контроль |
2438 |
2802 |
3070 |
5037 |
Примечание. Допустимая методическая ошибка определения не превышает 10–15 %.
Довольно высокая СМЧ в ризосфере наблюдалась в начале вегетационного сезона. Большое количество КОЕ в июне может быть обусловлено поздним сходом снежного покрова, началом активной фазы вегетации деревьев, ростом корней и размножением микроорганизмов ризосферы. Отмечены существенные различия в динамике численности микроорганизмов у таксономически близких сосны и кедра.
Почва вне зоны действия корней деревьев (контроль), в отличие от ризосферной почвы древостоев, при одинаковых температурных условиях имела более высокую влажность (до 40%), но в целом близкую с ризосферой суммарную численность микроорганизмов, и только в сентябре здесь зарегистрирован максимум количества микроорганизмов. Это связано, скорее всего, с поступлением «свежего» органического вещества в почву и уменьшением конкуренции за питательные элементы со стороны корневых систем.
При анализе средней за вегетационный пе- риод численности ризосферных микроорганизмов на диагностических средах (табл. 2) установлено, что максимальное развитие аммонифицирующих бактерий (рост на МПА) выявляется в контроле и под кедром. Показатели численности микроорганизмов, использующих минеральные источники азота (рост на КАА), имеют наибольшие значения в контроле, а наименьшие под елью. Максимальные коэффициенты микробиологической минерализации (КАА/МПА) регистрируются под сосной. Эти значения численности и коэффициентов минерализации свидетельствуют о несбалансированности процессов синтеза-ресинтеза под этими видами древесных насаждений. Максимальные количества олигонитрофильных бактерий определяются под кедром, а коэффициенты олиго-трофности (Эшби/МПА) имеют наибольшие значения под елью, что является свидетельством недостатка элементов питания под этими видами деревьев [3].
Таблица 2
Средняя численность микроорганизмов ризосферы под различными видами деревьев на разных диагностических средах, тыс. КОЕ/ г воздушно-сухой почвы
|
Вид хвойных |
МПА/ КА |
Эшби |
Чапека |
КАА |
КАА/ МПА |
Эш-би/МПА |
|
Сосна |
328,8 + 79,0 |
561,1 + 219,4 |
1870,0 + 134 |
766,2 + 369,3 |
2,83 |
1,94 |
|
Ель |
648,3 + 431,8 |
958,8 + 233,3 |
288,9 + 99,6 |
329,7 + 126,8 |
0,51 |
3,48 |
|
Кедр |
3291,9 + 2394,1 |
3827,6 + 1154,2 |
840,9 + 543,4 |
812,8 + 251,7 |
0,26 |
0,58 |
|
Контроль |
2017,4 + 607,5 |
782,9 + 376,7 |
1421,1 + 518,5 |
4204,7 + 1839,9 |
0,96 |
0,71 |
СЧМ филлосферы, определяемая на комплексной ГПД среде (табл. 3), в течение вегетационного периода имеет максимальные величины на хвое ели. Минимальные значения чис- ленности регистрируются на хвое сосны. На хвойных древостоях отмечены существенные колебания численности микроорганизмов по месяцам вегетационного периода. Поскольку исследуемые древесные насаждения находятся в одинаковых экологических условиях, выраженные колебания численности микроорганизмов филлосферы хвойных можно связать с из- менением фитонцидной активности в определенную фенофазу вегетации деревьев и с биологическими особенностями вида.
Таблица 3
Суммарная численность микроорганизмов филлосферы (эпифитов) различных видов деревьев в течение вегетационного сезона, тыс. КОЕ/ г воздушно-сухой почвы
|
Вид хвойных |
Июнь |
Июль |
Август |
Сентябрь |
|
Сосна |
780,8 |
293,3 |
301,7 |
533,3 |
|
Ель |
2673,3 |
1760,5 |
1950,7 |
1600,3 |
|
Кедр |
266,7 |
1080,7 |
1737,5 |
560,7 |
Примечание. СМЧ определена на глюкозо-пептоно-дрожжевой среде.
При сравнительном анализе СЧМ ризосферы и филлосферы различных видов деревьев проявились следующие тенденции: абрисы динамики численностей для всех видов исследуемых деревьев практически совпадают в течение вегетационного периода. Это, вероятно, связано с тем, что, как было сказано выше, комплекс «почва–дерево–микроорганизмы ризосферы и филлосферы» является экосистемой, компоненты которой связаны функционально и энергетически. Такая взаимосвязь и взаимозависимость компонентов сохраняют экосистему в состоянии гомеостаза.
Заключение. Исследование динамики численности различных групп эпифитных и ризосферных микроорганизмов на диагностических средах у сосны, ели и кедра свидетельствует об отличающихся максимальных и минимальных значениях численности микроорганизмов у разных видов исследуемых хвойных по фенофазам вегетации (июнь, июль, август, сентябрь). Очевидно, что динамика численности и структуры микробных комплексов у разных видов деревьев обусловлена, наряду с влиянием абиотических и биотических факторов, биологическими особенностями вида растений. При этом абрисы динамики суммарной численности микроорганизмов филлосферы (эпифитов) и ризосферы у каждого конкретного вида в течение периода вегетации совпадают, что свидетельствует о сопряженных взаимосвязях дерева и микробных комплексов.
Список литературы Структура и динамика микробных комплексов филлосферы и ризосферы хвойных видов лесных биогеоценозов Приенисейской Сибири (на примере Предивинского лесхоза)
- Методы почвенной микробиологии и био-химии. -М.: Изд-во МГУ, 1991. -304 с.
- Методы стационарного изучения почв. -М.: Наука, 1977. -248 с.
- Сорокин Н.Д. Микробиологическая диагно-стика лесорастительного состояния почв Средней Сибири. -Новосибирск: Изд-во СО РАН, 2009. -221 с.
- Теппер Е.З. Изучение бактерий корневой зоны растений и на корнях: практикум по микробиологии. -М.: Высш. шк., 1972. -198 с.
- Теппер Е.З. Практикум по микробиологии. -М.: Дрофа, 2004. -256 с.
