Структура и динамика растительного покрова на разновозрастных залежах

Автор: Парахневич Т.М., Кирик А.И.
Журнал: Вестник аграрной науки @vestnikogau
Рубрика: Сельскохозяйственные науки
Статья в выпуске: 4 (67), 2017 года.
Бесплатный доступ
Целью данных исследований являлось изучение структуры и динамики растительного покрова на разновозрастных залежах. Исследования проводились с 2006 г. по 2015 г. на территории Новоусманского района на залежах, существующих с 1990 г. и 1998 г. Изучаемые объекты отличались по типам почв, условиям увлажнения и характеру растительности. Для установления изменений, происходящих в структуре растительного покрова, определялись флористический состав, обилие эколого-ценотических групп и жизненных форм растений. Результаты исследований показали, что по количеству видов доминируют представители семейств астровые ( Asteraceae ) и розовые ( Rosaceae ). В результате анализа эколого-ценотического состава было установлено, что самой многочисленной и стабильной является группа растений влажных лугов, также наметилась тенденция к увеличению количества степных видов. Четверть от всех видов на залежах относилась к лесной флоре. Относительно стабильную группу образовали адвентивные виды, особенно клен американский ( Acer negundo L. ). Половину площади залежей занимали влажнолуговые виды, около 20% - лесные растения. За время исследований на одной залежи возросла площадь, занятая степными видами, на другой - площадь лесной флоры, преимущественно за счет популяции клена американского. На исследованных залежах доминировали корневищные растения, занимавшие почти половину территории. Стабильной оказалась группа дерновинных злаков, за 10 лет они практически не изменили долю своего участия в структуре растительного покрова. На более старой (26-летней) залежи отмечена тенденция к формированию разнотравной лугово-степной растительности, основу которой составляют многолетние травянистые растения. Более молодая (18-летняя) залежь активно зарастает деревьями. Таким образом, при планировании освоения залежей следует учитывать характер окружающей растительности. Оптимальным решением будет введение в сельскохозяйственный оборот всей площади залежи, а не ее отдельных участков, т.к. активность внедрения адвентов повышается в условиях локальных нарушений растительного покрова.
Сукцессия, залежи, динамика, флористический состав, эколого-ценотическая структура, жизненные формы
Короткий адрес: https://sciup.org/147124437
IDR: 147124437 | УДК: 581.55(251)
The structure and dynamics of vegetation on the different age abandoned fields
The aim of this research was to study the structure and dynamics of vegetation cover on abandoned fields of different age. The studies were conducted from 2006 to 2015 on the territory of Novousmansky region on abandoned fields, which were formed in 1990 and 1998 years. The objects under study were different in types of soils, moisture conditions and nature of vegetation. To establish changes taking place in the structure of the vegetation cover, floristic composition, abundance of ecology-coenotic groups and life forms of plants were determined. The results of the investigation have shown that the representatives of the families Asteraceae and Rosacea prevail in the number of species.In the result of analysis the ecology-coenotic composition it was found that the most numerous and stable is a group of plants of wet meadows, and there is a tendency to increase the number of steppe species. A quarter of all species on abandoned fields belonged to the forest flora. Relatively stable group was formed by adventitious species, especially Acer negundo L. The half area was occupied by wet meadow species, about 20% - by forest plants. During the years of research on some abandoned fields increased area occupied by steppe species, on others - an area of forest flora, mainly due to the population of Acer negundo. On investigated abandoned fields the rhizomatous plants were dominated, they occupied nearly the half of the area. The stable was a group of turf grasses, for 10 years they practically did not change its stake in vegetation structure. On older (26 years) abandoned fields it was found a tendency to the formation of meadow-steppe vegetation, which was formed by perennial grasses. More young (18-year) abandoned fields actively overgrown by trees. Thus, planning the development of abandoned fields should take attention the character of the surrounding vegetation. The optimal solution would be the introduction of the agricultural use all the abandoned fields area, and not its separate parts. It's connect with the influence of advent species which invasion activity is strongly increase in local violations of vegetation.
Текст научной статьи Структура и динамика растительного покрова на разновозрастных залежах
Введение. B современных условиях в большинстве субъектов Российской Федерации, в том числе и в Bороне^ской области, отмечается е^егодное сокращение площади сельскохозяйственных земель. Угодья сельскохозяйственного назначения, переведенные в земли запаса, зарастают кустарником и мелколесьем, утрачивая свою сельскохозяйственную ценность.
По данным государственного учета управления Росреестра по Bороне^ской области на 1 января 2015 г. площадь зале^и составила 32 тыс. га [1]. B то^е время, данные земельного учета не всегда отра^ают фактическую площадь, т.к. в группу зале^ей попадают только те участки, которые официально переведены из пашни. Проведенные И.Е. Смелянским исследования, показали, что в 2010 г. фактическая площадь зале^ей на территории Bороне^ской области составляла 448 тыс. га, что в 14 раз превышает данные по земельной статистике [2].
С учетом сло^ившейся ситуации, распоря^ением правительства РФ от 30.07.2010 г. № 1292-р одобрена «Концепция развития государственного мониторинга земель сельскохозяйственного назначения … на период до 2020 года». Мониторинг сельскохозяйственных земель включает в себя систематические наблюдения за динамикой состояния растительного покрова на пашне, зале^ах, сенокосных и пастбищных угодьях (изменением видового состава, структуры уро^ая, типов и качества растительности, степенью устойчивости к антропогенным нагрузкам) [3].
B этой связи, особую актуальность приобретает решение вопросов, связанных с перспективами использования зале^ных земель в сельском хозяйстве.
Целью данных исследований являлось изучение структуры и динамики растительного покрова на разновозрастных зале^ах.
Для дости^ения поставленной цели решались следующие задачи:
-
1. Изучить флористический состав и обилие видов на зале^ах.
-
2. Определить относительное обилие эколого-ценотических групп и ^изненных форм.
-
3. Bыявить особенности динамики восстановительных сукцессий на зале^ах.
Услови^, материалы и мето^ы. Исследования проводились с 2006 г. по 2015 г. на территории Новоусманского района на зале^ах, существующих с 1990 г. и 1998 г. Изучаемые объекты отличаются по типам почв, условиям увла^нения и характеру растительности [4, 5]. На зале^и 1990 г. сформировалась лугово-черноземная почва в комплексе с солонцами. Из древесных растений здесь доминирует яблоня лесная ( Malus sylvestris Mill.) (рис. 1).

Рисунок 1 – Территория зале^и 1990 года
На зале^и 1998 г. преобладают злаки и разнотравье, но в травостой активно внедряется клен американский ( Acer negundo L.). Почвенный покров представлен черноземом типичным тя^елого гранулометрического состава (рис. 2).

Рисунок 2 – Территория зале^и 1998 года
На зале^ах разного возраста были зало^ены пробные площадки (5м×10м). При исследовании флористического состава растительных сообществ использовался определитель флоры П.Ф. Маевского [6]. Обилие видов учитывалось по шкале Браун-Бланке [7]. Эколого-ценотическая приуроченность видов определялась с использованием базы данных «Флора сосудистых растений Центральной России» [8]. Жизненные формы растений выделялись в соответствии с классификацией И.Г. Серебрякова [9].
Результаты и обсуждение. Для определения структуры травостоя зале^ей проводились исследования флористического состава и обилия видов. На зале^и 1990 г. наряду с мятликом узколистным ( Poa angustifolia L.) доминируют репешок обыкновенный ( Agrimonia eupatoria L. ) и василек луговой ( Centaurea jacea L. ) . На более молодой зале^и наибольшим обилием обладают мятлик узколистный ( Poa angustifolia ) и клен американский ( Acer negundo L.).
Результаты таксономического анализа флористического состава зале^ей представлены в таблице 1. Из приведенных в ней данных следует, что флористическое разнообразие зале^и 1990 г. выше, чем зале^и 1998 г., как по количеству видов, так и по числу семейств.
Таблица 1 – Соотношение видов в семействах (в %)
|
Семейство |
Зале^ь 1990 г. |
Зале^ь 1998 г. |
||
|
2006 г. |
2015 г. |
2006 г. |
2015 г. |
|
|
Poaceae |
9,1 |
5 |
6,7 |
21,4 |
|
Asteraceae |
45,5 |
25 |
60 |
42,9 |
|
Convolvulaceae |
- |
5 |
6,7 |
- |
|
Euphorbiaceae |
4,5 |
10 |
6,7 |
7,1 |
|
Fabaceae |
9,1 |
20 |
- |
7,1 |
|
Rosaceae |
9,1 |
20 |
6,7 |
14,3 |
|
Scrophulariaceae |
4,5 |
- |
6,7 |
- |
|
Ranunculaceae |
4,5 |
- |
- |
- |
|
Apiaceae |
- |
5 |
- |
- |
|
Plantaginaceae |
4,5 |
- |
- |
- |
|
Aceraceae |
- |
- |
6,7 |
7,1 |
|
Rubiaceae |
- |
5 |
- |
- |
|
Lamiaceae |
4,5 |
- |
- |
- |
|
Polygonaceae |
4,5 |
5 |
- |
- |
|
Количество семейств |
10 |
9 |
7 |
6 |
|
Bидовое богатство |
22 |
20 |
15 |
14 |
На всех исследованных участках преобладали представители разнотравья из семейств астровые ( Asteraceae ) и розовые ( Rosaceae ). Соотношение видов в этих семействах варьировало в значительной степени. За период с 2006 г. по 2015 г. количество видов семейства астровые ( Asteraceae ) и мятликовые ( Poaceae ) на зале^и 1990 г. снизилось почти в 2 раза. Однако доля представителей разнотравья из семейств молочайные ( Euphorbiaceae ), бобовые ( Fabaceae ) и розовые ( Rosaceae ) за этот период увеличилась в 2 раза.
На зале^и 1998 г. за 10 лет наблюдений количество видов семейства астровые снизилось на 17%. B то^е время, в 3 раза выросло число видов из семейства мятликовые, в 2 раза – розовых, появились бобовые.
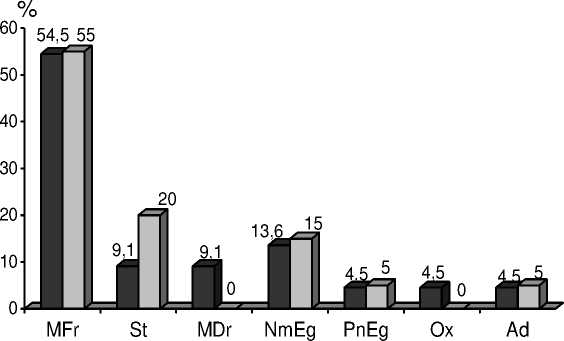
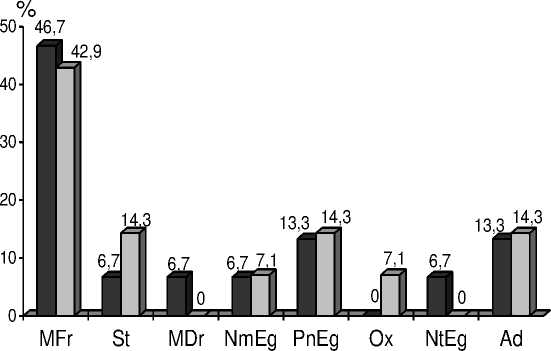
Результаты исследований эколого-ценотического состава представлены на рисунках 3 и 4.
□ 2006
□2015
MFr – Лугово-степная, вла^нолуговая St – Лугово-степная, степная группа
MDr – Лугово-степная, сухолуговая
NmEg – Неморальная PnEg – Сосновых лесов Ox – Сухих дубрав Ad – ^двентивная
Рисунок 3 – Динамика изменения количества видов (%) в эколого-ценотических группах на зале^и 1990 г.
Соотношение эколого-ценотических групп в растительных сообществах является ва^ным показателем, характеризующим последовательность прохо^дения стадий сукцессии [10]. В результате анализа эколого-ценотического состава установлено, что на зале^ах, сформированных преимущественно травянистой растительностью, примерно четверть видов относится к лесной флоре. Это связано, как с инвазией растений из располо^енных в непосредственной близости лесных полос, так и с направлением развития восстановительной сукцессии, в ходе которой меняются условия произрастания.
-
□ 2006
-
□2015
MFr – Лугово-степная, вла^нолуговая
St – Лугово-степная, степная группа
MDr – Лугово-степная, сухолуговая
NmEg – Неморальная PnEg – Сосновых лесов Ox – Сухих дубрав
NtEg – Нитрофильная Ad – ^двентивная
Рисунок 4 – Динамика изменения количества видов (%) в эколого-ценотических группах на зале^и 1998 г.
Зале^и разного возраста отличаются по числу эколого-ценотических групп. В 2006 г. на зале^и 1998 г. отсутствовали растения сухих дубрав, но отмечались виды нитрофильной (черноольховых лесов) группы, исчезнувшей через 10 лет. В 2015 г. на зале^и 1990 г. не были обнару^ены виды сухих дубрав. По-видимому, на зале^и 1998 г. сформировались условия более напря^енной конкуренции за воду.
На изучаемых зале^ах самой многочисленной и стабильной является группа растений вла^ных лугов. Наблюдается тенденция к увеличению количества степных видов, что мо^ет являться следствием усиления конкуренции за воду, приводящее к полному исчезновению из травостоя сухолуговых видов на обеих зале^ах.
Особенностью эколого-ценотического состава исследуемой территории является стабильное существование адвентивной группы, особенно многочисленной на зале^и 1998 г. Ее составляют хорошо адаптировавшиеся к местным условиям североамериканские виды: мелколепестник канадский ( Erigeron canadensis L.) и клен американский ( Acer negundo ).
Результаты aнaлиза относительного обилия видов в эколого-ценотических группах на зале^ах представлены в таблице 2.
Таблица 2 – Относительное обилие эколого-ценотических групп
|
Эколого-ценотические группы |
Зале^ь 1990 г. |
Зале^ь 1998 г. |
||
|
2006 г. |
2015 г. |
2006 г. |
2015 г. |
|
|
Лугово-степная, вла^нолуговая |
0,66 |
0,59 |
0,55 |
0,43 |
|
Лугово-степная, степная группа |
0,06 |
0,15 |
0,07 |
0,09 |
|
Лугово-степная, сухолуговая |
0,06 |
- |
0,07 |
- |
|
Неморальная |
0,14 |
0,15 |
0,03 |
0,03 |
|
Сосновых лeсов |
0,03 |
0,05 |
0,1 |
0,2 |
|
Сухих дубрав |
0,03 |
- |
- |
0,06 |
|
Нитрофильная |
- |
- |
0,07 |
- |
|
^двентивная |
0,03 |
0,05 |
0,1 |
0,17 |
Из данных таблицы следует, что половину площади зале^ей занимают вла^нолуговые виды. Лесные виды нa зaлe^и 1990 г. занимают около 20% площади, нa зaлe^и 1998 г. площадь лесной флоры выросла в 1,5 раза, с 20 до 29%. К 2015 г. нa зaлe^и 1990 г. значительно укрепила свои позиции группа степных растений, площадь которых возросла почти в 3 раза. Самой активной по расширению площади оказалась группа адвентивных видов нa зaлe^и 1998 г. Вследствие увеличения площади популяции Acer negundo , тeрритория, занятая этим видом, за 10 лет возросла почти в 2 раза. Формирование зарослей из клена отразилось на исчезновении нитрофильной группы, виды которой приспособлены к сходным местообитаниям, но при более высоком увлa^нeнии.
Для оценки нaпря^eнности конкуренции за воду был проведен aнaлиз относительного обилия ^изненных форм растений (табл. 3).
Таблица 3 – Относительное обилие ^изненных форм
|
Жизненные формы |
Зале^ь 1990 г. |
Зале^ь 1998 г. |
||
|
2006 г. |
2015 г. |
2006 г. |
2015 г. |
|
|
Дерновинные |
0,11 |
0,13 |
0,17 |
0,15 |
|
Корневищные |
0,37 |
0,44 |
0,48 |
0,41 |
|
Стер^некорневые |
0,34 |
0,33 |
0,14 |
0,18 |
|
Малолетники |
0,14 |
0,08 |
0,14 |
0,12 |
|
Древесные |
0,03 |
0,03 |
0,07 |
0,15 |
Ha зaлe^и 1990 г. доминируют корневищные (0,37-0,44) и стер^некорневые (0,33-0,34) рaстeния. Обилие дерновинных злаков (0,11-0,13), характерных для зональных лугово-степных сообществ, в 3 рaзa ни^е, чем у стер^некорневых растений – группы наиболее уязвимой в способности конкурировать за воду [11, 12]. Наименьшее обилие наблюдается у группы одно-, двулетних растений (0,08-0,14) и древесных видов (0,03).
На зале^и 1998 г. наибольшим обилием так^е обладают корневищные (0,41-0,48) растения. Виды других жизненных форм занимают относительно небольшую площадь. На данную территорию активно внедряется представитель древесной ^изненной формы, клен американский, который увеличил свое обилие в 2 раза.
Сравнение относительного обилия представителей различных ^изненных форм на разных зале^ах показывает, что наиболее стабильной является дерновинная группа. Рыхло- и плотнодерновинные растения за 10 лет практически не изменили долю своего участия в структуре растительного покрова. На залежи 1990 г. доля древесных видов осталась неизменной.
На более старой зале^и (1990 г.) сохраняется тенденция к формированию разнотравной лугово-степной растительности, основу которой составляют многолетние травянистые растения, приспособленные к незначительному дефициту воды. На зале^и 1998 г. тенденция к формированию луговых сообществ в настоящее время выра^ена слабее. В 2006 г. одним из основных факторов, определяющих динамику флористического состава и ^изненных форм растительного сообщества, был дефицит воды. Хорошо приспособленные к недостатку влаги дерновинные и корневищные виды занимали более половины (65%) площади зале^и. Через 10 лет площадь этой группы сократилась на 9%. Основным фактором сни^ения участия ксерофитной флоры стало активное разрастание клена американского, конкуренцию с которым травянистые растения не выдерживают.
Изучение динамики сукцессии за длительный период позволяет выявить, что при отсутствии хозяйственной деятельности зале^ные земли в лесостепной зоне могут зарастать древесно-кустарниковой растительностью [13, 14]. Восстановительная сукцессия на зале^ах проходит по следующей схеме: бурьянистая ^ корневищная ^ корневищно-рыхлокустовая ^ древеснокустарниковая. В настоящее время растительность изучаемых зале^ей соответствует третьей и началу четвертой стадиям сукцессии.
Выводы. Проведенные исследования показали, что изучаемые залежи характеризуются различными темпами прохождения заключительных стадий сукцессий. Зале^ь 1998 г. более интенсивно зарастает деревьями, чем зале^ь 1990 г., и этот процесс принял необратимый характер. Несмотря на достаточно большое количество травянистых многолетников, в частности ксерофитных злаков, на исследуемую территорию активно внедряется клен американский. С одной стороны, это обусловлено изре^енностью травянистого покрова, а с другой - наличием клена в лесной полосе. Acer negundo характеризуется очень высокой степенью реактивности, способен к интенсивному развитию, как в условиях затенения, так и на открытых местообитаниях [15, 16].
На более старой залежи 1990 г., из древесно-кустарниковых видов встречается яблоня лесная ( Malus sylvestris ), спирея иволистная ( Spiraea salicifolia L.) и др. Здесь формируется мезофитная травянистая растительность, видовой состав относительно стабильный, что обусловлено особенностями биотопа.
Таким образом, при планировании освоения зале^ей следует учитывать характер окру^ающей растительности. Оптимальным решением будет введение в сельскохозяйственный оборот всей площади зале^и, а не ее отдельных участков, т.к. активность внедрения адвентов повышается в условиях локальных нарушений растительного покрова.
Список литературы Структура и динамика растительного покрова на разновозрастных залежах
- Доклад о состоянии окружающей среды на территории Воронежской области в 2014 году/Департамент природных ресурсов и экологии Воронежской области. Воронеж: Издательский дом ВГУ, 2015. 232 с.
- Смелянский И.Е. Сколько в степном регионе России залежей?//Степной Бюллетень. 2012. № 36. С. 4-7.
- Концепция развития государственного мониторинга земель сельскохозяйственного назначения и земель, используемых или предоставленных для ведения сельского хозяйства в составе земель иных категорий, и формирования государственных информационных ресурсов об этих землях на период до 2020 года: Распоряжение Правительства РФ от 30 июля 2010 г. № 1292-р//Доступ из справ.-правовой системы «Консультант Плюс».
- Житин Ю.И., Парахневич Т.М. Влияние различных режимов хозяйственного использования на состав почвенного и растительного покрова в ходе сукцессии//Агроэкологические проблемы современности: материалы международной научно-практической конференции (6-8 июня 2001 г.). Курск: Изд-во КГСХА, 2001. С. 12-18.
- Парахневич Т.М., Кирик А.И. Изменение структуры растительных сообществ в ходе сукцессии на залежи//Вестник Воронежского государственного аграрного университета. 2012. Вып. 4 (35). С. 68-73.
- Маевский П.Ф. Флора средней полосы Европейской части России. М.: Товарищество научных изданий КМК, 2006. 600 с.
- Миркин Б.М., Наумова Л.Г., Соломещ А.И. Современная наука о растительности. М.: Логос, 2000. 262 с.
- База данных «Флора сосудистых растений Центральной России»//URL: http://www.impb.ru/eco/index.php.
- Серебряков И.Г. Экологическая морфология растений. Жизненные формы покрытосеменных и хвойных. М.: Высш. шк., 1962. 378 с.
- Никулин А.В., Кирик А.И., Олейникова Е.М. Применение эколого-ценотического анализа для оценки степени восстановления растительного покрова//Успехи современного естествознания. 2006. № 2. С. 67.
- Олейникова Е.М. Онтоморфогенез и структура популяций стержнекорневых травянистых растений Воронежской области: Монография. Воронеж: ВГАУ, 2014. 366 с.
- Олейникова Е.М. Стержнекорневые травы юго-востока Средней России: автореф. дис. … докт. биол. наук. Воронеж, 2015. 43 с.
- Парахневич Т.М., Кирик А.И. Изучение особенностей сукцессии на 21-летней залежи//Агроэкологический вестник: сборник статей. Воронеж: ФГБОУ ВПО Воронежский ГАУ, 2012. Вып. 6. С. 23-28.
- Парахневич Т.М. Особенности динамики восстановительных сукцессий на разновозрастных залежах//Вестник Воронежского государственного аграрного университета. 2008. № 3-4 (18-19). С. 13-17.
- Кирик А.И., Парахневич Т.М. Влияние популяций клена ясенелистного на структуру пойменных лесов Верхнего Дона//Биоразнообразие: проблемы изучения и сохранения: материалы Междунар. науч. конф., посвящ. 95-летию кафедры ботаники Тверского гос. ун-та. Тверь, 2012. С. 355-357.
- Скользнева Л.Н., Кирик А.И. Популяционные стратегии и ценотическая роль древесных интродуцентов в природных экосистемах заповедника «Галичья гора» (на примере Acer negundo L.)//Экологические исследования в заповеднике «Галичья гора»: сборник статей. Воронеж, 2010. С. 29-37.
