Структура населения и топические преферендумы дождевых червей (Oligochaeta, Lumbricidae) в почвах эталонных лесных формаций бассейнов рек Теберда и Большой Зеленчук (Тебердинский заповедник, Северо-Западный Кавказ)
 в почвах эталонных лесных формаций бассейнов рек Теберда и Большой Зеленчук (Тебердинский заповедник, Северо-Западный Кавказ)")
Автор: Рапопорт Ирина Борисовна, Цепкова Нэлли Лукинична
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Биологические ресурсы: фауна
Статья в выпуске: 6-1 т.17, 2015 года.
Бесплатный доступ
Изучены видовой состав и показатели обилия дождевых червей эталонных лесных биогеоценозов Тебердинского заповедника и прилегающих территорий. Наибольшее число видов отмечены в сосновых лесах. Наиболее «кавказский» облик отличает фауну пихтарников и грабовников, помимо стабильного космополитного блока, складывающуюся из двух крымско-кавказских эндемиков, кавказского по происхождению Dendrobaena tellermanica и видов, представляющих древнюю средиземноморскую фауну Кавказа. Дождевые черви грабовников характеризуются высоким общим обилием, максимальным средним биотопическим видовым богатством и близкой к максимальному значению выровненностью обилий дождевых червей. В пихтарниках меньше, чем в других биогеоценозах, представлены подстилочные виды и космополиты. Население кленовых, ольховых и березовых фитоценозов имеют схожую экологическую структуру. В березовых лесах отмечена наименьшая представленность собственно почвенных дождевых червей и частая встречаемость бореальных видов; в кленовниках - обширный спектр доминантов. В ольшаниках максимальна выровненность обилий люмбрицид. В почвенных пробах букняков отмечены только два вида, остальные люмбрициды стратифицированы под корой валежника, в зеленомошных синузиях и на гигрофитных участках. Индекс доминирования дождевых червей представляет верхний экстремум исследованных сообществ, биомасса, индексы биоразнообразия и выравненности обилий дождевых червей минимальны. Фауна лиственных и смешанных лесов и сообществ, включающих разнотравье в травяном ярусе, отличается от хвойных (мертвопокровных и злаковых) фитоценозов наиболее полной представленностью морфо-экологических форм. Наличие подзолов, или гидроморфный характер почв, ограничивают распространение норников и собственно почвенных видов. В березовых криволесьях, сосняках и ельниках отмечено замещение подстилочного D. octaedra средиземноморским подстилочным D. attemsi.
Дождевые черви, фауна, экология, топическая преференция, эталонные сообщества, тебердинский заповедник, северо-западный кавказ
Короткий адрес: https://sciup.org/148204248
IDR: 148204248 | УДК: 547.4:595.14
Population structure and topical preferendum of earthworms (Oligochaeta, Lumbricidae) in the soils of normal forest formations of the Teberda and Bolshoi Zelenchuk river basins (Teberda nature reserve, North-Western Caucasus)
The species composition and abundance rates of earthworms in normal forest biogeocenoses of Teberda nature reserve and adjacent territories have been investigated. The greatest species number is marked in pine forests. The most “caucasian” image distinguishes the fauna of fir and hornbeam forests in addition to stable cosmopolitan block, and is formed by two crimean-caucasian endemics - D. tellermanica, Caucasian by its origin and the species representing ancient Mediterranean fauna of the Caucasus. Hornbeam forest earthworms are characterized by general high population, maximum biotopiс species diversity and earthworm abundance evenness which is the closest to the maximum value. Debris epigeic and cosmopolites are less represented in fir forests than in other biogeocenoses. The soil population of maple, alder and birch formations have similar ecological structure. However, the minimum representation of endogenic earthworms and frequent occurrence of boreal species is marked in birch forests, a wide spectrum of dominants - in maple forests, and evenness of Lumbricidae abundance is maximum in alder forests. In soil samples of beech forests only two species are registered, the other Lumbricidae are stratified under the bark of brushwood, in green moss synusia and hygrophitic areas. Dominance index of earthworms represents the largest extremum of the researched communities, biomass, indexes of biodiversity and abundance evenness of earthworms are minimal. The fauna of deciduous and mixed forests and communities including motley grasses in the herbaceous layer differs from coniferous biogeocenoses (of dead soil coverings or gramineous) by the most total representation of morpho-ecological forms. Podzol occurrence or hydromorphic character of soils confines distribution of anecic and endogenic species. The substitution of epigeic D. octaedra by Mediterranean debris epigeic D. attemsi is registered in croocked birch, pine and spruce forests.
Текст научной статьи Структура населения и топические преферендумы дождевых червей (Oligochaeta, Lumbricidae) в почвах эталонных лесных формаций бассейнов рек Теберда и Большой Зеленчук (Тебердинский заповедник, Северо-Западный Кавказ)
заповедника (ТГПБЗ) проводилось с прошлого века [2-5], сведения о дождевых червях Архызского участка ТГПБЗ опубликованы недавно [6], неполными оставались данные о биотопической и высотнопоясной приуроченности видов.
Цель работы: выяснение особенностей топического и высотно-поясного распределения дождевых червей семейства Lumbricidae эталонных лесных сообществ Тебердинского заповедника и прилегающих территорий.
Материал и методика. Сбор дождевых червей проведен в 2011-2012 гг. в поясах темнохвойных, широколиственных лесов и субальпийском. Общий объем собранного материала составляет 215 почвенных проб. После проведения подробных геоботанических описаний, координатной и ландшафтной привязки к местности на площадках 30x30 м отбирали 5-8 проб. Раскопки проводили на глубину встречаемости дождевых червей. Почвенные монолиты 25х25 см2 разбирали вручную [7]. Дополнительно изучали распределение дождевых червей под корой валежника, в скоплениях опада и в зелено-мошных синузиях, под камнями по берегам ручьев. Определение люмбрицид и характеристика морфоэкологических форм приведены по работам Т.С. Перель [8, 9]. Для обработки полученных данных использованы: видовое богатство, средние общие численность и биомасса дождевых червей, численность видов, индексы Шеннона, Симпсона, Пиелу [10]. Экологическая характеристика древесных пород приведена согласно работам [11-13].
Работы выполнены в бассейнах рек Тебер-да, Большой Зеленчук и их притоков. Исследованы почвы фитоценозов с преобладанием:
-
1. ольхи черной (= о. клейкой), Alnus glutinosa (L.) Gaertn. (кленово-черноольхово-разно-травных; черноольхово-разнотравных, вязово-ольховоразнотравных);
-
2. клена Траутфеттера, Acer trautvetteri Medw. (кленово-разнотравных, березово-кленово-малиново-злаковых, осиново-пихтово-кленово-разнотравных, пихтово-кленово-разнотравных);
-
3. пихты Нордманна, Abies nordmanniana (Stev.) Spach (пихтарников кисличных и разнотравных, березово-пихтовых с кислицей, сосново-пихтовых с кислицей, буково-сосново-пихтово-редкопокровных, березово-пихтово-разнотравных, пихтово-ясменниковых, пихтово-ясменниковых с примесью ели, елово-пихтово-мертвопокровных с примесью сосны, пихтово-вейниковых, пихтово-мертво-покровных);
-
4. ели восточной Picea orientalis (L.) Link (ельников мертвопокровных, пихтово-елово-мертвопокровных);
-
5. сосны обыкновенной, Pinus silvestris L. (сосново-разнотравно-злаковых, елово-сосновозеленомошных, березово-сосновых злаково-разнотравных, березово-сосновыйх разнотравных, сосняков разнотравных, березово-сосново-высокотравных);
-
6. бука восточного, Fagus orientalis Lipsky (буково-мертвопокровных, буково-разнотравных);
-
7. граба кавказского, Carpinus caucasica Grossh. (буково-грабово-разнотравных с подростом клена, буково-грабово-разнотравных, грабово-разнотравных);
-
8. березы Литвинова, Betula litwinowii Doluch. (березово-разнотравных, березово-разнотравнозлаковых, сосново-березово-разнотравных, пихтово-березово-разнотравно-злаковых).
Краткое описание района исследований. Согласно типизации В.Е. Соколова, А.К. Темботова [14] рассматриваемая территория располагается в пределах кубанского варианта западно-северокавказского типа поясности, свойственного западной части северного макросклона Большого Кавказа и прилегающей равнины Западного Предкавказья. Климат более мягкий по сравнению с восточно-северокавказским типом поясности, т.к. формируется под влиянием воздушных масс Средиземно-Черноморского бассейна и Атлантического океана. Пояс широколиственных лесов кубанского варианта поясности расположен в пределах 10001500 м над ур. м. [14]. Доминируют бурые лесные и горно-лесные серые почвы [15]. За год в среднем выпадает 600-1500 мм осадков, лето теплое, зима умеренно холодная. Пояс темнохвойных лесов занимает склоны хребтов от 1000-1200 до 2000 м над ур. м. Климат влажный, умеренно холодный, преобладающие почвы - бурые лесные остаточно-оподзоленные, горно-лесные бурые, горно-лесные подзолистые [15]. Субальпийский пояс охватывает высоты 1700-1800 м до 2200 м над ур. м., отличается многоснежной зимой и среднегодовой суммой осадков около 1200 мм.
Результаты и их обсуждение. В лесных сообществах Тебердинского заповедника зарегистрированы 14 видов дождевых червей (табл. 1). Региональные эндемики отсутствуют, 2 вида -крымско-кавказские субэндемики, доминирующие по численности и биомассе в значительной части исследованных сообществ. Кавказским по проис-хождению является D. tellermanica . Дизъюнктивный ареал этого вида мы считаем восточ-ноевро-азиатским. 21% видов распространены в пределах средиземноморской области Голарк-тики. Наиболее редок из средиземноморских видов D. alpinа , на Западном Кавказе ранее известный по одному нахождению в районе Красной поляны (Кавказский заповедник) (Перель, 1979). Большин-ство отмеченных люмбрицид -космополиты (57%).
В лесных сообществах рек Теберда и Зеленчук встречаются все морфо-экологические группы дождевых червей. Крымско-кавказский эндемик D. schmidti представлен собственно почвенной, почвенно-подстилочной и редкой подстилочной морфо-экологическими формами, отличающимися размерами тела, наличием и характером пигментации, временем реакции на раздражение и глубиной стратификации в почвенном профиле [16]. По видовому богатству преобладают люмбрициды, питающиеся на поверхности почвы (62,5%).
Субальпийский пояс. В.З. Гулисашвили [17] выделял «зону сосновых и березовых лесов», хорошо выраженную на северных склонах Главного Кавказского хребта, начиная с бассейна Кубани, включая в него бассейн р. Теберда. Сосна и береза встречаются как в виде чистых древостоев, так и смешанных сосново-березовых и березово-сосновых лесов [17].
Леса с преобладанием березы Литвинова. Отмечены 7 видов люмбрицид (табл. 1, 2). Для дождевых червей березовых криволесий характерно содоминирование двух крымско-кавказских субэндемиков D. schmidti и D. mariupolienis и редкая встречаемость D. attemsi. D. octaedra и Dendro-drilus rubidus tenuis, широко распространенные в березовых криволесьях Центрального Кавказа [18], обычны на Тебердинском участке заповедника и не обнаружены в березняках в бас. р. Большой Зеленчук, возможно, в силу частичного замещения викари-рующим видом D. attemsi, или неполного охвата нами потен-циальных местообитаний. По берегам прогре-ваемых ручьев в изобилии встречаются E. Биотопическое видовое богатство невелико (табл. fetida и Eiseniella tetraedra, поднимающиеся здесь 2), видовые обилия имеют близкий порядок.
значительно выше, чем на Центральном Кавказе.
Таблица 1. Видовой состав, хорология и экология дождевых червей ТГПБЗ
|
Тип ареала |
Видовой состав |
Жизненная форма |
Бук |
Граб |
Клен |
льха |
Пихта |
Ель |
Сосна |
Береза |
|
1 |
Dendrobaena schmidti (Mich.) |
ПМ |
0,89 |
0,67 |
0,91 |
0,29 |
0,92 |
1 |
0,89 |
0,85 |
|
D. mariupolienis mariupolienis (Wyss.) |
Н |
- |
0,17 |
0,19 |
0,14 |
0,15 |
0,2 |
0,1 |
0,33 |
|
|
2 |
D. tellermanica Perel |
СП |
- |
0,17 |
0,09 |
- |
0,23 |
- |
0,11 |
- |
|
3 |
Octolasion lacteum (Örley) |
СП |
0,11 |
- |
- |
0,43 |
- |
- |
0,1 |
- |
|
Aporrectodea rosea (Sav.) |
СП |
- |
0,5 |
0,14 |
||||||
|
A. caliginosa trapezoidеs (Dug.) |
СП |
- |
- |
- |
- |
- |
- |
0,1 |
- |
|
|
Lumbricus rubellus (Hoffm.) |
ПП |
0,11 |
0,17 |
0,45 |
0,86 |
0,23 |
- |
0,14 |
- |
|
|
Eisenia fetida (Sav.) |
ПП |
0,11 |
- |
0,27 |
0,14 |
- |
- |
0,01 |
0,05 |
|
|
D. octaedra (Sav.) |
П |
0,67 |
0,67 |
0,55 |
0,14 |
0,54 |
0,5 |
0,1 |
0,2 |
|
|
Dendrodrilus rubidus tenuis (Eis.) |
П |
0,22 |
0,17 |
0,19 |
- |
0,23 |
1 |
0,1 |
0,1 |
|
|
Eiseniella tetraedra tetraedra (Sav.) |
П |
- |
- |
- |
- |
- |
- |
- |
0,05 |
|
|
4 |
D. attemsi Mich. |
П |
0,5 |
0,14 |
0,3 |
0,22 |
0,11 |
|||
|
D. alpina (Rosa) |
ПП |
- |
- |
- |
- |
0,08 |
- |
- |
- |
|
|
A. jassyensis (Mich.) |
СП |
- |
- |
- |
- |
0,08 |
- |
- |
Формации с преобладанием сосны обыкновенной часто произрастают на крутых склонах южной экспозиции на неполноразвитых олиготрофных почвах, отличаются разомкнутостью полога и малой густотой. Однако на пологих склонах с почвами среднемощными образуют густые одновозрастные древостои [17]. В сосновых лесах отмечены 10 видов люмбрицид (табл. 1, 2), из них 3 вида – Dendrodrilus rubidus tenuis, E. fetida, O. lacteum – исключительно под корой валежника и зеленомошными синузиями.
Таблица 2. Видов ое богатство и максимальная средняя численность доминирующих видов дождевых червей в порядке убывания встречаемости доминирования, в разных типах лесных формаций бассейнов рек Теберда и Большой Зеленчук
|
Преобладающая порода |
Видовое богатство |
Видовой состав и численность доминантов |
|
|
общее |
биотопи-ческое |
||
|
береза Литвинова |
7 |
1-4 |
D. schmidti ∗ , D. mariupolienis ∗ , D. octaedra , E. fetida ∗∗ |
|
сосна обыкновенная |
10 |
1-5 |
D. schmidti ∗ , L. rubellus ∗∗ , D. mariupolienis ∗ , D. octaedra, D. tellermanica |
|
ольха черная |
8 |
1-4 |
L. rubellus***, O. lacteum***, E. fetida**, D. schmidti* |
|
клен Траутфеттера |
7 |
1-4 |
D. schmidti*** , L. rubellus*** , E. fetida*** , D. octaedra*** , De. rubidus*** , D. mariupolienis |
|
бук восточный |
6 |
1-5 |
D. schmidti*, D. octaedra , De. rubidus, L. rubellus*** , O. lacteum** |
|
граб кавказский |
8 |
4-6 |
A. rosea*** , D. attemsi** , D. octaedra** , D. schmidti** , D. tellermanica |
|
ель восточная |
5 |
3-4 |
D. schmidti , D. attemsi , De. rubidus |
|
пихта Нордманна |
8 |
1-5 |
D. schmidti** , D. octaedra* , De. rubidus* |
Примечание: максимальная средняя биотопическая численность видов: без значка – 1-10 экз./м2; * - 11-50 экз./м2; ** - 51-100 экз./м2; *** - более 100 экз./м2
На вершинах водоразделов в сосняках вейниковых и редкотравно-злаковых дождевые черви, как правило, представлены 1 видом (D. schmidti, реже D. tellermanica, численность 3-5 экз./м2). В березово-сосновом злаково-разнотравном биогеоценозе, занимающем плакорную позицию, все черви относятся к виду D. mariupolienis (численность 48±11,31 экз./м2). Факт сам по себе интересен, т.к. почвы, в которых брались пробы - смытые песчаные, и плохо подходят для местообитаний норников, предпочитающих дренированные почвы с протяженным гумусовым горизонтом. Березово-сосново-разнотравный био-геоценоз отличается доминированием почвенно-подстилочного L. rubellus и представленностью как собственно почвенных (почвенная форма D. schmidti, O. lacteum), так и подстилочных (D. attemsi и D. octaedra) люм-брицид. Лишь однажды - на берегу минерального источника в сосняке разнотравном отмечен A. c. trapezoides. Наличие полидоминант-ного травяного яруса коррелирует с видовым богатством дождевых червей. Наибольшее число видов отмечено в почвах разнотравных сосновых фитоценозов (сосняках разнотравных и березово-сосноворазнотравных). Общая числен-ность дождевых червей варьирует от 2-3 до 200 экз./м2.
Пояс темнохвойных лесов. В формациях с преобладанием пихты (эдификатор - пихта кавказская, почвы - горно-лесные бурые) отмечены 8 видов (табл. 1, 2). Ядро фауны составляют D. schmidti , D. octaedra и D. tellermanica , часто встречается Dendrodrilus rubidus tenuis . Максимальное биотопическое видовое богатство - 5 видов . В пихтово-кисличных и сосново-пихтово-кисличных лесах отмечены 6 видов ( Dendro-drilus rubidus tenuis , L. rubellus , D. tellermanica , A. jassyensis , D. octaedra D. schmidti ), хотя в некоторой части этих сообществ встречается только D. schmidti , преимущественно почвенно-подстилочная, реже - собственно почвенная формы. В пихтарниках разнотравных отмечены виды, питающиеся на поверхности почвы ( D. alpina , D. octaedra , D. schmidti , Dendrodrilus rubidus tenuis , L. rubellus ) и отсутствуют собственно почвенные черви . Для злаковых формаций характерны D. schmidti и D. mariupolienis , численность которых имеет близкий порядок, для редкопокровных и мертвопокровных - содомини-рование D. Octaedra и Dendrodrilus rubidus tenuis , либо D. schmidti и D. tellermanica . В пихтовых биогеоценозах бассейна р. Теберда заметно присутствие средиземноморских видов - A. jassyensis и D. alpina и высокая численность Dendrodrilus rubidus tenuis (Eis.) . D. tellermanica и D. mariupolienis характерны для Архызского участка.
В ельниках отмечены 5 видов дождевых червей (табл. 1, 2), обилия которых имеют близкий порядок. На Архызском участке обычны почвенноподстилочная форма D. schmidti и 2 подстилочных вида: Dendrodrilus rubidus tenuis и D. attemsi. В.В. Зонн [15] почвы под еловыми лесами относит к типу горно-подзолистых. Отсутствие собственно почвенных видов в ельнике мертвопокровном скорее всего определяется промывным режимом почв и наличием оподзоленного горизонта с сильно кислой реакцией среды и бедным минеральным составом [19]. В пихтово-елово-мертвопокровном сообществе с примесью сосны (Тебер-динский участок) отмечен норник D. mariupolienis, почвенно-подстилочная форма D. schmidti и два подстилочных вида - D. octaedra и Dendrodrilus rubidus tenuis.
Пояс широколиственных лесов. Буковые леса. 1200-1400 м над ур. м. составляют верхнюю границу произрастания чистых буковых лесов на Западном Кавказе, хотя отдельные деревья бука встречаются до 2200-2400 м над ур. м. Бук восточный является мезофитом, однако плохо переносит переувлажнение и заболачивание. Буковые леса развиты на хорошо дренированных почвах, устойчивы к недостатку увлажнения. Букняки исследовались в июле, что, возможно, определило небогатый видовой состав дождевых червей. В почвенных пробах отмечены только два вида - D. schmidti (доминант) и D. octaedra . Dendrodrilus rubidus tenuis и E. fetida собраны под корой валежника, зеленомошных синузиях и по берегам ручьев. L. rubellus , O. lacteum доминируют на пойменных участках кленово-буковых лесов . Биотопическое видовое богатство дождевых червей является нижним экстремумом исследованных сообществ (табл. 2).
В формациях с преобладанием граба работы проводились на высоте 1000-1335 м над ур. м. Зарегистрированы 8 видов люмбрицид (табл. 1, 2). Как и в луговых биогеоценозах этого высотного уровня, доминирует A. rosea (бас. р. Большой Зеленчук) , на высотах от 1200-1300 м над ур. м. замещаемый собственно почвенной формой D. schmidti . Часто встречаются D. atemsi , D. octaedra, D. tellermanica и Dendrodrilus rubidus tenuis. Нор-ник D. mariupolienis характерен для хорошо дренированных почв с развитым гумусовым профилем, L. rubellus - для влажных участков. Среднее биото-пическое видовое богатство максимально. Грабовые леса отличаются высокой численностью люм-брицид (до 325 экз./м2).
Формации с доминированием клена. Клен, как правило, предпочитает глубокие, рыхлые, плодородные, глинисто-песчаные, слабокислые или нейтральные почвы. Не переносит чрезмер-ную сухость, засоление и заболачивание. В почвах кле-новников зарегистрированы 7 видов дождевых червей (табл. 1, 2). Наиболее часто доминирует D. schmidti, или D. octaedra , на более влажных участках - L. rubellus , на гигрофитных - L. rubellus и E. fetida. В кленовниках бассейна р. Теберда, в отличии от Архызского участка, часто встречались E. fetida и L. rubellus.
Интразональные формации с доминированием ольхи черной и ольхи серой формируются на полугидроморфных, плодородных, преимущественно глинистых кислых, или слабокислых почвах. Ольха черная является мезотрофом и гигрофитом. Встречается 8 видов дождевых червей. Для большинства ольшаников, развитых на днищах ущелий, характерно содоминирование O. lacteum и L. rubellus, на глинистых северных и северо-западных склонах - L. rubellus и E. fetida. В вязово-черноольховых сообществах высокой численности достигает D. schmidti, обычны подстилочные D. octaedra и D. attemsi. Нахождение норника D. mariupolienis в кленово-ольхово-разнотравном лесу свидетельствует о глубоко залегающих грунтовых водах и хорошем почвенном дренаже. В ольшаниках, развитых на низких надпойменных террасах, встречается A. rosea. Ольховые леса бас. р. Большой Зеленчук отличаются заметным участием A. rosea и D. attemsi. E. fetida отмечен в бас. р. Теберда, но пока не найден на Архызском участке. Максимальное видовое богатство (4 вида) наблюдалось на переувлажненных участках кленово-ольхово-разнотравного леса, в остальных биогеценозах отмечены 2-3 вида люмбрицид. В ольшанике разнотравном зарегистрирована высокая средняя численность - 304 экз./м2.
Индексы биоразнообразия, доминирования и выровненности. Максимальное биоразнообразие люмбрицид (индекс Шеннона) отмечено в почвах ельников, выровненность их обилий - в ольшаниках. Минимальны оба показателя в буковых лесах (рис. 1). Доминирование (индекс Симпсона) имеет обратный порядок.
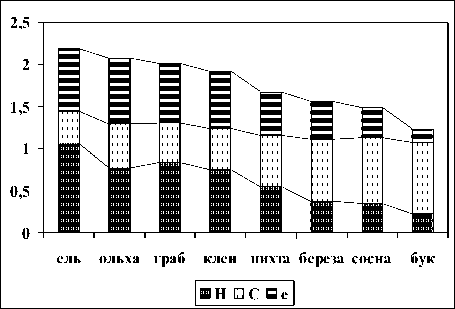
Рис. 1. Индексы Шеннона (Н), Симпсона (С) и Пие-лу (е) дождевых червей эталонных биотических сообществ в бассейнах рек Теберда и Большой Зеленчук
В целом, биогеоценозы, занимающие местоположения с близкими эдафо-климатическими условиями, характеризуются схожими значениями синэкологических индексов дождевых червей. Первый блок представлен сообществами грабовых, кленовых и ольховых лесов, произрастающих на хорошо увлажненных почвах с богатым минеральным составом. Более высокогорные березовые и сосновые леса - также обнаруживают схожесть структурной организации сообществ обитающих в них дождевых червей.
Показатели обилия дождевых червей. Наибольшие средние общие численность и биомасса люмбрицид наблюдались в грабовых лесах, наименьшие - в ельниках (рис. 2). Близкие величины показателей обилия от отмечены у дождевых червей, обитающих в почвах зональных формаций с преобладанием пихты, березы, сосны и бука. Сильнее всего варьируют средние численность и биомасса дождевых червей сосновых лесов.
Соотношение морфо-экологических форм дождевых червей. В большинстве рассмотренных биогеоценозов отмечены все жизненные формы люмбрицид (рис. 4). В ельниках мертвопокровных оподзоленный горизонт с сильно кислой реакцией среды и бедным минеральным составом [19] ограничивает распространение собственно почвенных видов.
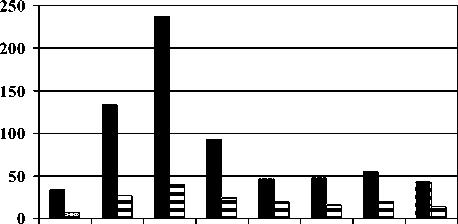
ель ольха граб клен пихта береза сосна бук
■ численность биомасса
Рис. 2. Общие численность (экз./м2) и биомасса (г/м2) дождевых червей в почвах эталонных биотических сообществ бассейнов рек Теберда и Большой Зеленчук
Хорологическая структура сообществ люм-брицид. Как и везде на Кавказе, самой наиболее многочисленны космополиты (рис. 3). «Кавказский» облик у населения люмбрицид пихтарников и грабовников, помимо стабильного космополитного блока, складывающийся из двух крымско-кавказских эндемиков, кавказского по происхождению восточноевро-азиатского D. tellermanica и представляющих наиболее древнюю средиземноморскую фауну Кавказа D. attemsi , D. alpina и A. jassyensis , из которых только A. jassyensis широко распространен на Северном Кавказе. В еловых лесах пока не найдены черви с восточноевроазиатским типом ареала, но соотношение космополитов и кавказского блока остается прежним. Сообщества люмбрицид кленовых, ольховых и березовых биогеоценозов имеют схожую структуру, за тем исключением, что в кленовниках средиземноморские виды замещаются восточноевроазиатским.
100%
80%
60%
40%
20%
0%
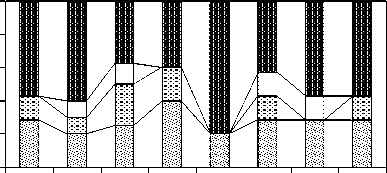
кавказские средиземноморские восточноевроазиатские космополитные
Рис. 3. Соотношение хорологических групп дождевых червей в почвах эталонных лесных формаций Тебердинского заповедника
В ельниках мертвопокровных оподзоленный горизонт с сильно кислой реакцией среды и бедным минеральным составом [19] ограничивает распространение собственно почвенных видов. Мало представлена эта морфо-эко-логическая группа и в субальпийских березовых криволесьях, но, в данном случае, лимитирующим фактором выступает краткость безморозного периода [20].
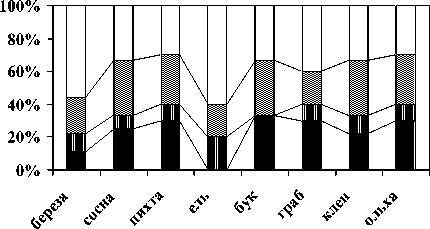
почвенные норники почвенно-подстилочные подстилочные
Рис. 4. Соотношение морфо-экологических форм дождевых червей в почвах эталонных лесных формаций Тебердинского заповедника
В буковых лесах не зарегистрированы норники, возможно, по причине засушливого лета 2011 г. Значительную роль сыграла так же высота над уровнем моря, на меньших высотах в мае-июне структура фауны и обилие дождевых червей буковых лесов Западного Кавказа могут существенно отличаться.
Выводы: в общей сложности отмечены 14 видов дождевых червей, из них в мелколиственных лесах – 7, в хвойных – 12 и широколиственных – 10 видов. Ядро фауны составляют поли-зональные D. schmidti , D. octaedra , Dendrodrilus rubidus tenuis , неморальные D. attemsi и L. rubellus и неморально-степной D. mariupolienis. D. schmidti – полиморфный вид с широкой экологической валентностью, отмеченный во всех типах рассмотренных сооб-ществ, в значительной части которых является доминантом. Занимает местообитания с широким диапазоном влажности – от мезоксеро-фитных сос-няков на вершинах водоразделов до гигрофитных участков ольховых сообществ. Молодые особи часто отмечаются под корой валежника и зеленомошными синузиями.
D. octaedra, Dendrodrilus rubidus tenuis и D. attemsi – подстилочные виды, экологические требования которых имеют некоторые отличия. Из них наиболее обычен D. octaedra, высокой численности достигающий в лиственных лесах и разнотравных хвойных формациях. Встречается этот вид и в почвах мертвопокровных букняков и ельников. Dendrodrilus rubidus tenuis и D. attemsi тяготеют к хвойным биогеоценозам. D. attemsi также отмечен на злаково-разнотравных участках буковых и ольховых сообществ. Dendrodrilus rubidus tenuis занимает сырые местообитания со значительным развитием зеленомошных участков, обычен в смешанных лесах, включающих хвойные породы, а также буковых и еловых мертвопокровных биогеоценозах. Все три вида на сухих участках лесов встречаются во влажной древесине упавших деревьев, подо мхом и по берегам ручьев. Немораль-но-степной D. mariupolienis обычен в субальпийском поясе в березовых лесах с полидоминантной структурой травяного яруса со значительной долей злаков. В поясе широколиственных лесов отмечен в разнотравных кленовых, ольховых лесах и пихтарниках. Другой неморально-степной собственно почвенный вид - D. tellermanica – зарегистрирован в редколесных с участием сосны и пихты биогеоценозах. L. rubellus, O. lacteum и E. fetida - индикаторы близкого залегания грунтовых вод. Чаще встречаются в ольшаниках и в полидоминантных сообществах. В других типах леса обычны по берегам ручьев, в западинах под влажным опадом, под корой валежника и подо мхом. 1000-1500 м над ур. м. представляют верхний предел распространения собственно-почвенного A. rosea, отмеченного исключительно в разнотравных грабовых и ольховых лесах на участках с периодическим или постоянным грунтовым увлажнением.
Четыре вида люмбрицид относятся в наших сборах к категории редких. A. caliginosa trapezoides отмечен в гигрофитной почве у минерального источника в сосняке разнотравном, A. jassyensis – в пихтарнике кисличном, D. alpina в пихтарнике травянистом, Eiseniella tetraedra – на высоте 2058 м над ур. м., в русле ручья в сосново-березовом разнотравном лесу. Из них, возможно, A. jassyensis и Eiseniella tetraedra распространены более широко и не представлены в наших сборах ввиду неполного охвата сообществ.
Искренне благодарим Т.С. Всеволодову-Перель, без постоянной творческой поддержки которой была бы не возможна проделанная работа, Ф.А. Темботову, Е.П. Кононенко за участие в работе над статьей и важные критические замечания. Ю.М. Саблирову – за помощь в геоботанической классификации фитоценозов Архызского участка.
Работа выполнена при финансовой поддержке программы фундаментальных исследований Президиума РАН «Живая природа: современное состояние и проблемы развития».
Список литературы Структура населения и топические преферендумы дождевых червей (Oligochaeta, Lumbricidae) в почвах эталонных лесных формаций бассейнов рек Теберда и Большой Зеленчук (Тебердинский заповедник, Северо-Западный Кавказ)
- Поливанова, H.H. Тебердинский заповедник Заповедники СССР//Заповедники Кавказа. -М., 1990. С. 18-34.
- Малевич, И.И. Дождевые черви Тебердинского Госзаповедника//Материалы III зоол. конф. педагогич. ин-тов РСФСР. -Волгоград, 1967. С. 313-315.
- Перель, Т.С. Дождевые черви в почвах лесов Северо-Западного Кавказа//Влияние животных на продуктивность лесных биогеоценозов. -М.: Наука, 1966. С. 146-165.
- Квавадзе, Э.Ш. Дождевые черви (Lumbricidae) Кавказа. -Тбилиси: Мецниереба, 1985. 283 с.
- Onipchenko, V.G. The structure of large soil invertebrate communities (Mesofauna) in the alpine ecosystems of the Teberda Reserve, the Northwestern Caucasus/V.G. Onipchenko, O.E. Znakova//Ecologia. -Montana, 1997. № 6. P. 35-38.
- Рапопорт, И.Б. Биотопическое распределение дождевых червей (Oligochaeta, Lumbricidae) в Тебердинской заповедной территории с наиболее высокой степенью охраны (Архызский участок, Северо-Западный Кавказ)//Современные проб-лемы особо охраняемых природных территорий регионального значения и пути их решения: Матер. Межрег. науч-практ. конф. -Воронеж, 2014. С. 214-218.
- Гиляров, М.С. Учет крупных беспозвоночных (мезофауны)//Методы почвенно-зоологических исследований. -М.: Наука, 1975. С. 12-29.
- Перель, Т.С. Распространение и закономерности распределения дождевых червей фауны СССР. -М.: Наука, 1979. 275 с.
- Всеволодова-Перель, Т.С. Дождевые черви фауны России. -М.: Наука, 1997. 102 с.
- Одум, Ю. Основы экологии. -М.: Мир, 1975. С. 186.
- Справочник лесничего. -М.: ВНИИЛМ, 2003. 640 с.
- Погребняк, П.С. Основы лесной типологии. -Киев: изд-во Академии наук Украинской ССР, 1955. 456 с.
- Сукачёв, В.Н. Идея развития в фитоценологии//Сов. ботаника. -М., 1942. №1-3. С. 5-17.
- Соколов, В.Е. Млекопитающие. Насекомоядные/В.Е. Соколов, А.К. Темботов. -М.: Наука, 1989. С. 3-27.
- Зонн, С.В. Горно-лесные почвы Северо-Западного Кавказа. -М.-Л.: Изд-во Академии наук СССР, 1950. 233 с.
- Rapoport, I.B. Morpfo-ecological forms of Dendrobaena schmidti Мichaelsen, 1907 (Oligochaeta, Lumbricidae) of North Caucasus//Fourth International Oligochaete Taxonomy Meetings. Book of Abstracts. -Diyarbakir, Turkey, 2009. Р. 39.
- Гулисашвили, В.З. Природные зоны и естественно-исторические области Кавказа. -М.: Наука, 1964. 325 с.
- Рапопорт, И.Б. Видовой состав и численность дождевых червей (Oligochaeta, Lumbricidae) пояса широколиственных лесов терского варианта поясности (Центральный Кавказ)//Животный мир горных территорий. -М.: Тов-во науч. изд-й КМК, 2009. С.118-123.
- Казеев, К.Ш. Биология почв Юга России/К.Ш. Казеев, С.И. Колесников, В.Ф. Вальков. -Ростов-на Дону: Изд-во ЦВВР, 2004. 350 с.
- Рапопорт, И.Б. Высотное распределение дождевых червей (Oligochaeta, Lumbricidae) в центральной части Северного Кавказа//Зоол. журн. 2013. № 1. С. 3-10.
