Структура населения пауков и сенокосцев (Arachnida, Araneae, Opiliones) модельных биоценозов Висимского биосферного заповедника
 модельных биоценозов Висимского биосферного заповедника")
Автор: С. Л. Есюнин, Н. Л. Ухова, А. М. Домолазова
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Зоология
Статья в выпуске: 3, 2021 года.
Бесплатный доступ
Группировки герпетобионтных пауков четырех местообитаний: двух семилетних гарей, еловоберезового и пихтово-елового лесов были исследованы в Висимском заповеднике (Свердловская обл.) в течение мая-сентября 2017 г. методом почвенных ловушек. Структура населения пауков гаревых биотопов существенно отличается от таковой лесов по видовому составу, попадаемости (экз./100 ловушко-суток), сезонной динамике и доминантным видам. Различия в структуре населения наиболее выражены в летний период. В летних группировках пауков гарей наиболее многочисленны Agyneta allosubtilis, Alopecosa pulverulenta, P. fulvipes, P. lugubris, P. riparia и Piratula hygrophila, в лесах – Allomengea scopigera и Haplodrassus soerenseni.
Пирогенные сообщества, сезонная динамика, мониторинг, горные леса, Средний Урал.
Короткий адрес: https://sciup.org/147235112
IDR: 147235112 | УДК: 595.44:581.524.3(1-924.95) | DOI: 10.17072/1994-9952-2021-3-191-205
Structure of spider-harvestman assemblages (Arachnida, Araneae, Opiliones) of model biotopes of Visimskiy natural reserve
The assemblages of herpetobion spiders from four habitats: two seven-year-old burns, spruce-birch and fir-spruce forests, were examined in the Visimskiy Reserve (Sverdlovsk Region) during May-September 2017 using pitfall-traps. The structure of the spiders population of burns significantly differs from that of forests in species composition, total occurrence rate (ind./100 trap-days), seasonal dynamics and composition of the dominant species. Differences in the structure of the population are most pronounced in the summer. In summer spider assemblages of burns, the most abundant are Agyneta allosubtilis, Alopecosa pulverulenta, P. fulvipes, P. lugubris, P. riparia and Piratula hygrophila, in forests - Allomengea scopigera and Haplodrassus soerenseni.
Текст научной статьи Структура населения пауков и сенокосцев (Arachnida, Araneae, Opiliones) модельных биоценозов Висимского биосферного заповедника
Таежные лесные пожары, являясь повторяющимися воздействиями, оказывают не только непосредственное влияние на животных, но и существенно изменяют среду их обитания [Niklasson, Granström, 2000; Wardle et al., 2003; Mordkovich et al., 2008; Gongalskij, Persson, 2013; Gorbunova et al., 2017].
Пауки являются удобной модельной группой для исследования последствий лесных пожаров, т.к. обычно они встречаются в большом количестве [Huhta, 1971; Coyle, 1981; Jennings et al., 1988; Coddington, Young, Coyle, 1996; Buddle, Spence, Langor, 2000], а распределение и численность отдельных видов тесно связаны со структурными характеристиками среды обитания [Uetz, 1979; Hatley, Macmahon, 1980; Pajunen et al.,
-
1995] . Пауки также являются ключевыми элементами детритных и пастбищных пищевых сетей лесных экосистем [Wise, 1993; Atlegrim, Sjoberg 1995; Nyffeler 2000].
Несмотря на наметившееся в последнее время увеличение интереса к изучению пирогенных группировок пауков бореальных лесов [Hauge, Kvamme, 1983; Aitchison-Benell, 1994; Buddle, Spence, Langor, 2000; Koponen, 2005; Larrivée, Fahrig, Drapeau, 2005; Samu et al., 2010], ощущается существенный недостаток исходной информации [Gongalskij, Persson, 2013].
Первые сведения о пауках и сенокосцах Висим-ского заповедника были получены нами в ходе комплексного изучения почвенной мезофауны коренных и производных лесных сообществ заповедника в 1989–1994 гг. Результаты данных исследо-
ваний послужили основой для организации мониторинговых наблюдений на данной территории. К настоящему времени в фауне Висимского заповедника выявлено 253 вида пауков и 5 видов сенокосцев, описано их биотопическое распределение [Есюнин, Ухова, 2011].
Катастрофические природные явления: массовый ветровал 1995 г. и последующие пожары 1998 и 2010 гг. в северо-восточной части заповедника, привели к существенным перестройкам группировок почвенно-подстилочных и герпетобионтных беспозвоночных. Нами было показано, что в населении пауков ветровальных участков увеличилось количество видов, характерных для лугов, начальных стадий сукцессий и термофильных видов [Есюнин, Ефимик, Мазура, 2000; Есюнин и др., 2001; Есюнин, Шумиловских, 2003]. Трансформация биоценозов после сильных пожаров оказывается более значительной, чем после ветровала. В первый год после пожара гаревые группировки пауков характеризовались высокой специфичностью структуры населения. Наблюдалось значительное снижение количества видов, а у таксонов, переживших пожар, - значимое снижение численности. Из паукообразных на свежей гари (месяц после пожара) были выявлены только два вида пауков: Robertas lividus и Centromerus arcanus . В конце лета число видов пауков увеличилось до десяти, и они составляли около 60% от обилия герпетобио-нтных беспозвоночных. Названная нами первичнопирогенной [Есюнин, Шумиловских, 2003], эта группировка беспозвоночных представляла из себя остаточный, сильно обедненный по количеству видов и особей комплекс исходного пихто-ельника.
В процессе восстановления кустарниковой и травяной растительности первично-пирогенная группировка трансформировалась в травянокустарниковую. Первые два года (1999 и 2000 гг.) наблюдалось быстрое изменение структуры и видового состава группировок беспозвоночных [Есюнин, Шумиловских, 2003]. В отличие от жужелиц, для которых характерна дискретность сукцессионных стадий, структура комплексов паукообразных изменялась континуально. Общим трендом было постепенное увеличение значимости паука Allomengea scopigera .
На второй и пятый годы после повторного пожара сохранились существенные различия между населением пауков горевших и контрольных участков как по количественным, так и по качественным показателям [Ухова и др., 2014; Ухова, Есюнин, 2016]. Если в населении пауков контрольного пихто-ельника наиболее обильными видами были A. scopigera, Agyneta allosubtilis, Alo-pecosa taeniata , то в населении послепожарного сообщества - Pardosa riparia и P. fulvipes .
Данное исследование является продолжением мониторинга изменений структуры населения паукообразных в послепожарных сообществах Висим-ского заповедника.
Материал и методы исследования
Мониторинг населения беспозвоночных животных в Висимском заповеднике ведется на пробных зоологических площадках (ПЗП). В 2017 г. сбор материала по структуре герпетобионтного комплекса производился непрерывно в течение всего вегетационного периода (середина мая - середина сентября). Для сбора материала использовались почвенные ловушки (ловушки Барбера) - стеклянные банки объемом 0.5 л., диаметром 73 мм, на треть наполненные фиксатором (этиленгликоль). Периодичность сбора материала - две недели в августе, 10 дней - в прочие месяцы. Наблюдения велись на четырех ПЗП. Подробное описание биотопов дано в работах Р.З. Сибгатуллина [2014, 2018]. Краткое описание обследованных площадок приводится ниже.
ПЗП-2 - малиново-кипрейно-вейниковое по-слепожарное сообщество (кв. 112; 57°23,757'N, 59°44,515'E). В 1995 г. коренной лес был нарушен катастрофическим ветровалом. Впоследствии оставшийся древостой усыхал и выпадал, а сохранившийся подрост из пихты сибирской ( Abies sibirica ) и ели сибирской ( Picea obovata ) активно развивался. В 2010 г. пробная площадь оказалась на границе пожара и выгорела мозаично, пятнами. В настоящее время живого древостоя нет. В 2013 г. началось активное возобновление березы пушистой ( Betula pubescens ), ивы козьей ( Salix caprea ). B травянистом ярусе доминируют иван-чай узколистный ( Chamaenerion angustifolium ), вейники Лангсдорфа ( Calamagrostis langsdorffii ) и притупленный ( Calamagrostis obtusata ).
ПЗП-20 - кипрейно-вейниковое послепожарное сообщество (кв. 112; 57°23,988'N, 59°43,833'E). В 1995 г. коренной лес был нарушен катастрофическим ветровалом; в 1998 г. - интенсивным низовым пожаром. Впоследствии оставшийся древостой усыхал и выпадал. В 2010 г. по участку снова прошёл пожар, он прошёл бегло, задерживаясь только на обгоревших ранее ветровальных деревьях, многие из которых на этот раз сгорели дотла. С 2013 г. отмечено возобновление ивы козьей, единично осины ( Populus tremula ). B травянистом ярусе доминируют вейники притупленный и Лангсдорфа, иван-чай узколистный.
ПЗП-7 - елово-березовый вейниково-высокотравный лес (кв. 123; 57°23,300'N, 59°44,624'E). Состав древостоя - 5Б4Е1П, подроста - 8Е2П. Подлесок редкий. В травяно-кустарничковом ярусе доминируют вейник притуплённый, перловник поникающий (Melica nutans), цицербита уральская (Cicerbita uralensis), сныть обыкновенная (Aego- podium podagraria), щучка дернистая (Deschampsia cespitosa), копытень европейский (Asarum europa-eum), кислица обыкновенная (Oxalis acetosella).
ПЗП-19 – пихто-ельник высокотравно-папоротниковый коренной (кв. 112; 57°23,908'N, 59°44,307'E). Площадь сохранившегося пихто-ель-ника около 1000 га. Он окружен гарями, ветровальными участками и разновозрастными березняками. Состав древостоя 6П4Е. Древостой разреженный, около 18% древостоя составляет сухостой. Подрост разреженный, представлен пихтой и елью. В подлеске господствует малина обыкновенная ( Rubus idaeus ), бузина сибирская ( Sambucus sibirica ), смородина щетинистая ( Ribes hispidulum ) и черёмуха обыкновенная ( Padus avium ). В травяном ярусе доминируют щитовник распростертый ( Dryopteris assimilis ), щучка дернистая ( Descha-mpsia cespitosa ), кислица обыкновенная, вейник притупленный.
Предварительный анализ показал, что первичные пробы паукообразных кластеризуются в сезонные группы. Это позволило сгруппировать данные, объединив их в весеннюю (середина мая – первая декада июня), летнюю (вторая декада июня – первая декада августа) и осеннюю (вторая декада августа – середина сентября) группы проб. В статье анализируются усредненные показатели для выделенных аспектов населения паукообразных.
В качестве показателя доминирования использовалась пятибалльная, ограниченная сверху лога-рифметическая шкала, предложенная Ю.А. Песен-ко [1982]. Данные по сезонной динамике апрокси-мировались встроенными средствами программы Microsoft Excel – методом полиномиальной регрессии. Структура населения пауков проанализирована посредством алгоритма Detrended correspondence analysis (DCA; без трендовый анализ соответствия), выполненного в программе PAST [Hammer, Harper, Ryan, 2001].
Население гарей
На малиново-кипрейно-вейниковой гари (ПЗП-2) отмечено максимальное количество видов пауков (65), и 5 видов сенокосцев (табл. 1).
В сезонной динамике попадаемости пауков отмечены два пика: первый приходится на июнь, второй – на первую декаду сентября (рис. 1А) с максимальной попадаемостью 210 и 246 экз./100 лов. сут., соответственно. Летом наиболее многочисленны половозрелые пауки-волки, такие как A. taeniata, P. lugubris, P. riparia, P. hygrophilus , и гнафозида H. soerenseni (табл. 1). Осенью обильны неполовозрелые особи A. taeniata и пауки-балдахинники: A. scopigera, D. concolor, T. mengei .
Таблица 1
Видовой состав, попадаемость (экз./100 лов. сут..) и некоторые показатели разнообразия населения пауков и сенокосцев малиново-кипрейно-вейниковой гари (ПЗП-2) и вейниково-кипрейной гари (ПЗП-20)
|
Таксон |
Биотопы |
|||||
|
ПЗП-2 |
ПЗП-20 |
|||||
|
Весна |
Лето |
Осень |
Весна |
Лето |
Осень |
|
|
Aranei |
||||||
|
Araneidae |
||||||
|
Araneus marmoreus (Clerck, 1757) |
- |
- |
0.3 |
- |
- |
- |
|
Cercidia prominens (Westring, 1851) |
1.2 |
- |
- |
- |
- |
- |
|
Clubionidae |
||||||
|
Clubiona kulczynskii (Lessert, 1905) |
- |
3.2 |
- |
- |
2.3 |
0.2 |
|
Clubiona subtilis (L. Koch, 1867) |
- |
- |
- |
- |
0.2 |
- |
|
Clubiona неполовозрелые |
- |
- |
0.4 |
- |
0.4 |
0.2 |
|
Dictynidae |
||||||
|
Mastigusa arietina (Thorell, 1871) |
- |
- |
0.4 |
- |
- |
- |
|
Gnaphosidae |
||||||
|
Arboricaria subopaca (Westring, 1861) |
- |
0.2 |
- |
- |
- |
- |
|
Gnaphosa неполовозрелые |
- |
0.7 |
0.4 |
- |
- |
- |
|
Haplodrassus cognatus (Westring, 1861) |
- |
0.2 |
- |
- |
- |
- |
|
Haplodrassus moderatus (Kulczynski, 1897) |
- |
0.2 |
- |
- |
- |
- |
|
Haplodrassus signifer (C. L. Koch, 1839) |
- |
- |
- |
- |
- |
0.2 |
|
Haplodrassus soerenseni (Strand, 1900) |
- |
11.7 |
4.0 |
- |
8.2 |
- |
|
Haplodrassus неполовозрелые |
- |
1.3 |
1.4 |
- |
0.3 |
1.0 |
|
Micaria pulicaria (Sundevall, 1831) |
- |
- |
- |
- |
0.3 |
- |
|
Zelotes azsheganovae Esyunin, Efimik, 1992 |
- |
- |
- |
- |
- |
0.4 |
|
Zelotes clivicola (L. Koch, 1870) |
- |
3.1 |
- |
- |
0.3 |
- |
Продолжение табл. 1
|
Таксон |
Биотопы |
|||||
|
ПЗП-2 |
ПЗП-20 |
|||||
|
Весна |
Лето |
Осень |
Весна |
Лето |
Осень |
|
|
Zelotes latreillei (Simon, 1878) |
- |
- |
- |
0.8 |
0.5 |
0.7 |
|
Zelotes subterraneus (C. L. Koch, 1833) |
- |
0.2 |
- |
- |
- |
0.2 |
|
Zelotes неполовозрелые |
- |
0.6 |
0.4 |
- |
- |
0.5 |
|
Linyphiidae |
||||||
|
Agyneta affinis (Kulczyński, 1898) |
- |
- |
- |
- |
0.5 |
- |
|
Agyneta allosubtilis (Loksa, 1965) |
1.2 |
6.5 |
1.6 |
- |
0.3 |
0.6 |
|
Agyneta conigera (O.P.-Cambridge, 1863) |
- |
0.3 |
- |
- |
0.3 |
- |
|
Agyneta olivacea (Emerton, 1882) |
- |
5.6 |
- |
- |
0.7 |
- |
|
Agyneta ramosa (Jackson, 1912) |
- |
0.7 |
- |
- |
- |
- |
|
Anguliphantes angulipalpis (Westring, 1851) |
- |
0.5 |
- |
- |
0.3 |
- |
|
Araeoncus humilis (Blackwall, 1841) |
- |
0.2 |
- |
- |
- |
- |
|
Allomengea scopigera (Grube, 1859) |
- |
0.4 |
13.2 |
- |
- |
- |
|
Bathyphantes gracilis (Blackwall, 1841) |
- |
2.8 |
0.8 |
- |
6.3 |
3.6 |
|
Bathyphantes nigrinus (Westring, 1851) |
- |
0.4 |
0.4 |
- |
- |
- |
|
Bolyphantes alticeps (Sundevall, 1833) |
- |
- |
1.1 |
- |
- |
- |
|
Centromerus arcanus (O.P.-Cambridge, 1873) |
2.4 |
0.5 |
0.3 |
0.8 |
1.6 |
- |
|
Centromerus sylvaticus (Blackwall, 1841) |
- |
- |
3.1 |
- |
- |
0.5 |
|
Ceratinella scabrosa (O.P.-Cambridge, 1871) |
- |
0.2 |
- |
- |
- |
- |
|
Cnephalocotes obscurus (Blackwall, 1834) |
- |
- |
- |
1.7 |
- |
- |
|
Decipiphantes decipiens (L. Koch, 1879) |
- |
- |
0.4 |
- |
- |
- |
|
Dicymbium tibiale (Blackwall, 1836) |
1.2 |
- |
- |
- |
- |
- |
|
Diplocentria bidentata (Emerton, 1882) |
- |
0.3 |
- |
- |
- |
- |
|
Diplostyla concolor (Wider, 1834) |
4.8 |
2.8 |
8.4 |
- |
- |
- |
|
Drapetisca socialis (Sundevall, 1833) |
- |
- |
0.7 |
- |
- |
- |
|
Erigone atra (Blackwall, 1833) |
1.2 |
- |
- |
- |
- |
- |
|
Erigonella hiemalis (Blackwall, 1841) |
1.2 |
0.3 |
- |
1.6 |
0.7 |
- |
|
Floronia bucculenta (Clerck, 1757) |
- |
- |
0.4 |
- |
- |
- |
|
Gonatium rubellum (Blackwall, 1841) |
- |
- |
0.9 |
- |
- |
- |
|
Gongylidiellum latebricola (O. P.-C., 1871) |
- |
- |
- |
- |
0.3 |
0.3 |
|
Helophora insignis (Blackwall, 1841) |
- |
- |
0.7 |
- |
- |
- |
|
Incestophantes kochiellus (Strand, 1900) |
1.2 |
- |
- |
- |
- |
- |
|
Kaestneria pullata (O.P.-Cambridge, 1863) |
- |
- |
- |
- |
- |
0.2 |
|
Metopobactrus prominulus (O. P.-C., 1873) |
- |
- |
- |
0.8 |
0.3 |
- |
|
Micrargus herbigradus (Blackwall, 1854) |
1.2 |
1.8 |
- |
- |
0.2 |
- |
|
Microneta viaria (Blackwall, 1841) |
- |
0.9 |
1.1 |
- |
- |
- |
|
Neriene clathrata (Sundevall, 1830) |
1.2 |
- |
- |
- |
- |
- |
|
Oedothorax agrestis (Blackwall, 1853) |
- |
- |
- |
- |
0.3 |
- |
|
Oryphantes angulatus (O. P.-C., 1881) |
1.4 |
0.2 |
- |
1.7 |
- |
0.2 |
|
Palliduphantes alutacius (Simon, 1884) |
- |
1.6 |
4.6 |
- |
- |
4.3 |
|
Panamomops dybowskii (O. P.-C., 1873) |
2.4 |
- |
0.5 |
- |
0.7 |
- |
|
Pelecopsis mengei (Simon, 1884) |
- |
- |
- |
- |
0.3 |
- |
|
Pocadicnemis pumila (Blackwall, 1841) |
- |
- |
- |
- |
2.8 |
0.2 |
|
Tapinocyba insecta (L. Koch, 1869) |
- |
- |
- |
- |
0.3 |
- |
|
Taranucnus setosus (O. P.-Cambridge, 1863) |
- |
- |
- |
21.7 |
0.2 |
- |
|
Tenuiphantes alacris (Blackwall, 1853) |
- |
0.8 |
0.8 |
- |
- |
- |
|
Tenuiphantes mengei (Kulczyński, 1887) |
- |
- |
17.3 |
- |
- |
8.4 |
|
Tenuiphantes tenebricola (Wider, 1834) |
- |
4.9 |
3.8 |
- |
- |
- |
|
Walckenaeria antica (Wider, 1834) |
- |
1.4 |
0.3 |
1.7 |
1.7 |
0.7 |
|
Walckenaeria atrotibialis (O. P.-C., 1878) |
- |
2.4 |
0.5 |
- |
- |
0.2 |
|
Walckenaeria mitrata (Menge, 1868) |
- |
- |
- |
0.8 |
- |
- |
|
Walckenaeria unicornis (O. P.-C., 1861) |
- |
0.6 |
- |
- |
0.2 |
- |
Продолжение табл. 1
|
Таксон |
Биотопы |
|||||
|
ПЗП-2 |
ПЗП-20 |
|||||
|
Весна |
Лето |
Осень |
Весна |
Лето |
Осень |
|
|
Wubanoides uralensis (Pakhorukov, 1981) |
- |
0.2 |
- |
- |
- |
- |
|
Zornella cultrigera (L. Koch, 1879) |
- |
0.2 |
0.4 |
- |
- |
- |
|
Linyphiidae неполовозрелые |
2.4 |
4.4 |
6.6 |
- |
1.6 |
2.8 |
|
Liocranidae |
||||||
|
Agroeca brunnea (Blackwall, 1833) |
1.2 |
0.8 |
0.8 |
- |
0.4 |
0.2 |
|
Agroeca proxima (O.P.-Cambridge, 1871) |
- |
- |
9.3 |
- |
- |
5.7 |
|
Agroeca неполовозрелые |
- |
- |
- |
- |
0.4 |
- |
|
Lycosidae |
||||||
|
Acantholycosa lignaria (Clerck, 1757) |
- |
0.7 |
0.4 |
- |
- |
- |
|
Acantholycosa norvegica (Thorell, 1872) |
- |
0.2 |
- |
- |
- |
- |
|
Acantholycosa неполовозрелые |
- |
0.2 |
0.7 |
- |
- |
0.2 |
|
Alopecosa pulverulenta (Clerck, 1757) |
- |
- |
- |
16.7 |
7.6 |
- |
|
Alopecosa taeniata (C.L.Koch, 1835) |
- |
13.1 |
17.5 |
- |
0.4 |
0.2 |
|
Alopecosa неполовозрелые |
- |
- |
- |
- |
0.8 |
3.6 |
|
Pardosa fulvipes (Collett, 1876) |
- |
0.2 |
0.5 |
- |
18.5 |
11.2 |
|
Pardosa lugubris (Walckenaer, 1802) |
1.2 |
27.4 |
2.4 |
- |
2.3 |
- |
|
Pardosa riparia (C.L.Koch, 1833) |
- |
20.6 |
1.9 |
- |
60.5 |
19.6 |
|
Pardosa sphagnicola (Dahl, 1908) |
- |
- |
- |
- |
2.5 |
- |
|
Pardosa неполовозрелые |
4.8 |
4.4 |
- |
12.5 |
8.3 |
1.1 |
|
Pirata hygrophilus (Thorell, 1871) |
- |
10.0 |
3.2 |
- |
3.4 |
0.8 |
|
Trochosa ruricola (DeGeer, 1778) |
1.2 |
0.2 |
1.2 |
2.5 |
0.5 |
0.7 |
|
Trochosa spinipalpis (F.O.P.-C., 1895) |
- |
- |
- |
0.8 |
0.2 |
- |
|
Trochosa неполовозрелые |
1.2 |
0.4 |
1.7 |
- |
1.1 |
2.0 |
|
Xerolycosa nemoralis (Westring, 1861) |
1.2 |
0.4 |
- |
- |
0.3 |
0.2 |
|
Mimetidae |
||||||
|
Ero furcata (Villers, 1789) |
- |
- |
- |
0.8 |
- |
0.9 |
|
Philodromidae |
||||||
|
Tibellus oblongus (Walckenaer, 1802) |
- |
- |
- |
- |
- |
0.2 |
|
Phrurolithidae |
||||||
|
Phrurolithus festivus (C. L. Koch, 1835) |
- |
- |
- |
- |
0.3 |
- |
|
Pisauridae |
||||||
|
Pisaura mirabilis (Clerck, 1757) |
- |
- |
- |
- |
0.6 |
- |
|
Sparassidae |
||||||
|
Micrommata roseum (Clerck, 1758) |
- |
- |
- |
- |
0.4 |
- |
|
Tetragnathidae |
||||||
|
Tetragnatha неполовозрелые |
- |
- |
- |
0.8 |
- |
- |
|
Metellina неполовозрелые |
- |
0.2 |
0.0 |
- |
- |
- |
|
Pachygnatha listeri (Sundevall, 1830) |
3.6 |
1.0 |
0.5 |
- |
- |
- |
|
Theridiidae |
||||||
|
Dipoena неполовозрелые |
- |
- |
- |
- |
0.3 |
- |
|
Euryopis flavomaculata (C. L. Koch, 1836) |
- |
- |
- |
- |
1.0 |
- |
|
Robertus lividus (Blackwall, 1836) |
2.4 |
1.0 |
1.5 |
1.6 |
0.4 |
3.0 |
|
Thomisidae |
||||||
|
Ozyptila trux (Blackwall, 1846) |
- |
- |
- |
- |
1.5 |
0.2 |
|
Xysticus cristatus (Clerck, 1757) |
- |
- |
- |
- |
0.3 |
- |
|
Xysticus luctuosus (Blackwall, 1836) |
- |
2.9 |
- |
- |
0.8 |
- |
|
Xysticus неполовозрелые |
- |
0.2 |
0.8 |
- |
- |
- |
|
Zoridae |
||||||
|
Zora spinimana (Sundevall, 1833) |
- |
0.4 |
- |
1 - 1 |
2.6 1 |
- |
Окончание табл. 1
|
Таксон |
Биотопы |
|||||
|
ПЗП-2 |
ПЗП-20 |
|||||
|
Весна |
Лето |
Осень |
Весна |
Лето |
Осень |
|
|
Opiliones |
||||||
|
Phalangiidae |
||||||
|
Lacinius ephippiatus (C.L. Koch, 1835) |
- |
6.3 |
- |
- |
0.6 |
0.4 |
|
Mitopus morio (Fabricius, 1779) |
- |
1.7 |
- |
- |
0.2 |
0.3 |
|
Oligolophus tridens (C. L. Koch, 1836) |
- |
- |
2.6 |
- |
- |
0.5 |
|
Rilaena triangularis (Herbst, 1799) |
1.2 |
0.2 |
- |
1.7 |
1.3 |
- |
|
Nemastomatidae |
||||||
|
Nemastoma lugubris (O.F. Müller, 1776) |
1.2 |
6.9 |
4.6 |
- |
0.3 |
1.2 |
|
Средняя попадаемость (пауки/сенокосцы) |
39.7/ 2.4 |
146.4/ 15.0 |
117.7/ 7.2 |
67.5/ 1.7 |
147.0/ 2.3 |
76.3/ 2.4 |
|
Кол-во видов (пауки/сенокосцы) |
18/2 |
48/4 |
40/2 |
16/1 |
47/4 |
28/4 |
Таблица 2
Видовой состав, попадаемость (экз./100 лов. сут.) и некоторые показатели разнообразия населения пауков и сенокосцев в березняке вейниково-высокотравном (ПЗП-7) и пихто-ельнике коренном (ПЗП-19)
|
Таксон |
Биотоп |
|||||
|
ПЗП-7 |
ПЗП-19 |
|||||
|
Весна |
Лето |
Осень |
Весна |
Лето |
Осень |
|
|
Aranei |
||||||
|
Clubionidae |
||||||
|
Clubiona неполовозрелые |
- |
- |
- |
- |
0.6 |
0.4 |
|
Dictynidae |
||||||
|
Dictyna неполовозрелые |
- |
- |
- |
- |
0.2 |
- |
|
Gnaphosidae |
||||||
|
Gnaphosa неполовозрелые |
- |
- |
- |
- |
- |
0.2 |
|
Haplodrassus cognatus (Westring, 1861) |
- |
- |
- |
- |
- |
0.4 |
|
Haplodrassus soerenseni (Strand, 1900) |
- |
4.8 |
- |
- |
5.9 |
- |
|
Haplodrassus неполовозрелые |
- |
- |
- |
- |
- |
0.5 |
|
Zelotes clivicola (L. Koch, 1870) |
- |
- |
- |
2.0 |
1.1 |
0.4 |
|
Zelotes subterraneus (C. L. Koch, 1833) |
- |
0.3 |
- |
- |
- |
- |
|
Hahniidae |
||||||
|
Cryphoeca silvicola (C. L. Koch, 1834) |
0.8 |
- |
- |
- |
- |
- |
|
Linyphiidae |
||||||
|
Abiskoa abiskoensis (Holm, 1945) |
- |
- |
- |
1.0 |
- |
0.7 |
|
Agyneta allosubtilis (Loksa, 1965) |
0.8 |
1.1 |
4.4 |
- |
3.6 |
27.2 |
|
Agyneta conigera (O.P.-Cambridge, 1863) |
- |
- |
- |
- |
2.5 |
0.6 |
|
Agyneta mossica (Schikora, 1993) |
- |
- |
- |
- |
0.9 |
- |
|
Agyneta olivacea (Emerton, 1882) |
- |
- |
- |
- |
4.1 |
- |
|
Agyneta tibialis (Tanasevitch, 2005) |
- |
- |
- |
- |
1.3 |
0.2 |
|
Allomengea scopigera (Grube, 1859) |
- |
24.5 |
112.5 |
- |
- |
51.6 |
|
Anguliphantes angulipalpis (Westring, 1851) |
0.8 |
- |
- |
- |
0.6 |
- |
|
Bathyphantes gracilis (Blackwall, 1841) |
- |
0.2 |
- |
- |
1.9 |
- |
|
Bathyphantes nigrinus (Westring, 1851) |
- |
0.8 |
0.3 |
- |
0.7 |
0.8 |
|
Bolyphantes alticeps (Sundevall, 1833) |
- |
- |
1.6 |
- |
- |
2.5 |
|
Centromerus arcanus (O. P.-Cambr., 1873) |
- |
- |
- |
- |
2.6 |
0.4 |
|
Centromerus sylvaticus (Blackwall, 1841) |
0.8 |
0.5 |
21.1 |
- |
- |
11.7 |
|
Продолжение табл. 2 |
||||||
|
Таксон |
Биотоп |
|||||
|
ПЗП-7 |
ПЗП-19 |
|||||
|
Весна |
Лето |
Осень |
Весна |
Лето |
Осень |
|
|
Ceratinella brevis (Wider, 1834) |
0.8 |
- |
- |
- |
- |
- |
|
Decipiphantes decipiens (L. Koch, 1879) |
- |
- |
0.4 |
- |
0.6 |
0.7 |
|
Diplocentria bidentata (Emerton, 1882) |
- |
- |
- |
- |
1.2 |
- |
|
Diplostyla concolor (Wider, 1834) |
0.8 |
0.4 |
0.5 |
2.0 |
3.8 |
4.2 |
|
Drapetisca socialis (Sundevall, 1833) |
- |
- |
- |
- |
- |
0.9 |
|
Erigonella hiemalis (Blackwall, 1841) |
- |
- |
- |
- |
0.2 |
- |
|
Flagelliphantes bergstroemi (Schenkel, 1931) |
0.8 |
- |
- |
- |
- |
- |
|
Gonatium rubellum (Blackwall, 1841) |
- |
- |
- |
- |
- |
0.4 |
|
Helophora insignis (Blackwall, 1841) |
- |
- |
- |
- |
- |
0.8 |
|
Hypselistes jacksoni (O. P.-Cambridge, 1903) |
- |
- |
- |
1.0 |
- |
- |
|
Incestophantes kochiellus (Strand, 1900) |
- |
- |
- |
3.1 |
0.5 |
1.7 |
|
Macrargus rufus (Wider, 1834) |
3.3 |
- |
- |
- |
- |
0.5 |
|
Micrargus herbigradus (Blackwall, 1854) |
- |
- |
- |
- |
2.3 |
0.6 |
|
Microneta viaria (Blackwall, 1841) |
0.8 |
0.4 |
- |
- |
- |
0.3 |
|
Oreonetides vaginatus (Thorell, 1872) |
0.8 |
- |
- |
- |
- |
- |
|
Oryphantes angulatus (O. P.-Cambr., 1881) |
- |
- |
- |
3.1 |
3.3 |
- |
|
Palliduphantes alutacius (Simon, 1884) |
0.8 |
1.1 |
0.7 |
1.0 |
1.8 |
10.2 |
|
Panamomops dybowskii (O. P.-Cambr., 1873) |
- |
- |
- |
1.0 |
0.3 |
- |
|
Pocadicnemis pumilla (Blackwall, 1841) |
- |
- |
- |
- |
- |
0.2 |
|
Savignya producta (Holm, 1977) |
- |
- |
- |
1.0 |
0.5 |
0.6 |
|
Tapinocyba insecta (L. Koch, 1869) |
- |
0.5 |
- |
- |
- |
- |
|
Tapinopa longidens (Wider, 1834) |
- |
- |
0.3 |
- |
- |
- |
|
Tenuiphantes alacris (Blackwall, 1853) |
- |
0.2 |
- |
5.2 |
4.3 |
0.9 |
|
Tenuiphantes nigriventris (L. Koch, 1879) |
- |
0.4 |
0.8 |
- |
0.3 |
4.9 |
|
Tenuiphantes tenebricola (Wider, 1834) |
0.8 |
2.3 |
10.2 |
- |
3.8 |
11.9 |
|
Tibioplus diversus (L. Koch, 1879) |
- |
- |
- |
- |
- |
0.8 |
|
Walckenaeria antica (Wider, 1834) |
- |
- |
- |
- |
0.9 |
|
|
Walckenaeria atrotibialis (O. P.-C., 1878) |
- |
0.7 |
0.5 |
- |
1.5 |
0.5 |
|
Walckenaeria mitrata (Menge, 1868) |
0.8 |
0.2 |
- |
- |
- |
- |
|
Walckenaeria nudipalpis (Westring, 1851) |
0.8 |
- |
- |
- |
- |
- |
|
Walckenaeria obtusa (Blackwall, 1836) |
- |
0.7 |
- |
- |
- |
- |
|
Wubanoides uralensis (Pakhorukov, 1981) |
- |
- |
- |
- |
0.2 |
- |
|
Zornella cultrigera (L. Koch, 1879) |
- |
- |
- |
1.0 |
- |
0.5 |
|
Linyphiidae неполовозрелые |
1.7 |
1.1 |
1.0 |
2.1 |
14.5 |
10.4 |
|
Liocranidae |
||||||
|
Agroeca brunnea (Blackwall, 1833) |
2.5 |
0.4 |
0.4 |
2.0 |
- |
- |
|
Agroeca proxima (O. P.-Cambridge, 1871) |
- |
- |
0.3 |
- |
- |
1.1 |
|
Lycosidae |
||||||
|
Acantholycosa norvegica (Thorell, 1872) |
- |
- |
- |
- |
0.3 |
- |
|
Acantholycosa неполовозрелые |
- |
0.3 |
- |
- |
- |
- |
|
Alopecosa taeniata (C.L.Koch, 1835) |
- |
1.1 |
- |
2.0 |
2.1 |
1.7 |
|
Alopecosa pinetorum (Thorell, 1856) |
- |
0.2 |
- |
- |
- |
- |
|
Alopecosa неполовозрелые |
- |
- |
- |
2.-1 |
0.2 |
- |
|
Pardosa lugubris (Walckenaer, 1802) |
- |
0.6 |
- |
- |
1.3 |
0.6 |
|
Pardosa неполовозрелые |
0.8 |
0.2 |
0.4 |
- |
0.3 |
- |
|
Piratula hygrophilus (Thorell, 1872) |
- |
1.0 |
0.3 |
- |
0.5 |
0.5 |
|
Trochosa ruricola (De Geer, 1778) |
0.8 |
0.4 |
- |
- |
- |
- |
|
Trochosa spinipalpis (F. O. P.-Cambr., 1895) |
- |
1.3 |
- |
- |
- |
- |
|
Trochosa terricola (Thorell, 1856) |
0.8 |
- |
- |
- |
- |
- |
|
Metidae |
||||||
|
Metellina mengei (Blackwall, 1869) |
- |
- |
- |
- |
- |
0.4 |
|
Metellina неполовозрелые |
- |
- |
- |
- |
- |
0.2 |
|
Mimetidae |
||||||
|
Ero furcata (Villers, 1789) |
- 1 |
- 1 |
0.3 |
- |
- 1 |
0.4 |
Окончание табл. 2
|
Таксон |
Биотоп |
|||||
|
ПЗП-7 |
ПЗП-19 |
|||||
|
Весна |
Лето |
Осень |
Весна 1 |
Лето |
Осень |
|
|
Tetragnathidae |
||||||
|
Pachygnatha listeri (Sundevall, 1830) |
- |
0.2 |
0.3 |
- |
- |
- |
|
Tetragnatha неполовозрелые |
- |
- |
- |
- |
0.2 |
- |
|
Theridiidae |
||||||
|
Robertus lividus (Blackwall, 1836) |
- \ |
0.9 1 |
1.1 |
- 1 |
1.4 1 |
0.5 |
|
Thomisidae |
||||||
|
Xysticus luctuosus (Blackwall, 1836) |
- |
- |
- |
- |
0.3 |
- |
|
Xysticus cristatus (Clerck, 1757) |
- |
- |
- |
1.0 |
- |
- |
|
Zoridae |
||||||
|
Zora spinimana (Sundevall, 1833) |
- I |
0.2 |
- |
- 1 |
- 1 |
- |
|
Opiliones |
||||||
|
Phalangiidae |
||||||
|
Lacinius ephippiatus (C.L. Koch, 1835) |
- |
5.8 |
2.0 |
- |
3.2 |
- |
|
Mitopus morio (Fabricius, 1779) |
- |
14.4 |
0.9 |
- |
4.1 |
0.4 |
|
Oligolophus tridens (C. L. Koch, 1836) |
- |
- |
8.8 |
- |
- |
2.9 |
|
Rilaena triangularis (Herbst, 1799) |
- |
- |
0.3 |
1.0 |
1.1 |
- |
|
Nemastomatidae |
||||||
|
Nemastoma lugubris (O.F. Müller, 1776) |
2.5 |
1.5 |
5.0 |
- |
0.3 |
4.0 |
|
Попадаемость (пауки/сенокосцы) |
20.8 2.5 |
46.8 21.7 |
157.1 16.9 |
31.3 1.0 |
72.0 8.7 |
153.5 7.3 |
|
Кол-во видов (пауки/сенокосцы) |
18/1 |
28/3 |
19/5 |
14/1 |
37/4 |
38/3 |
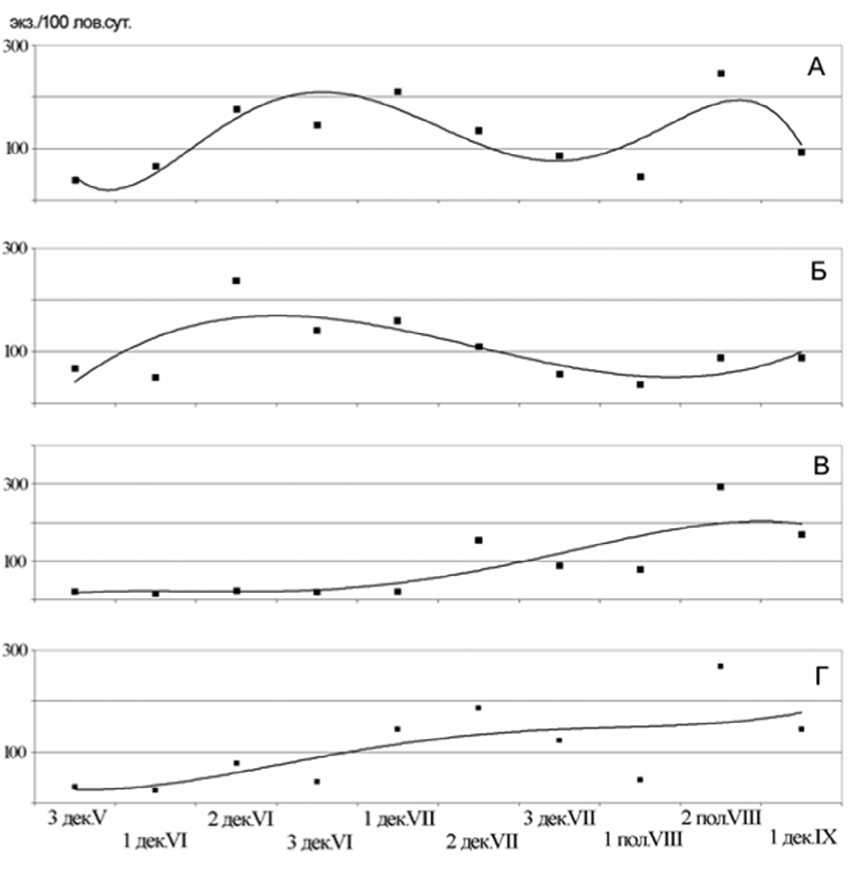
Рис. 1 . Сезонная динамика попадаемости пауков на малиново-кипрейно-вейниковой (А) и кипрейно-вейниковой (Б) гарях, в елово-березовом лесу (В) и в пихто-ельнике (Г)
В весенний сезон почти все виды пауков и сенокосцев были представлены единичными особями, обильные виды не зарегистрированы. Видовое
разнообразие пауков изменялось аналогично обилию группы.
Максимальное количество видов сенокосцев
отмечено в летний период (табл. 1). Попадаемость сенокосцев имеет один летний пик. Летом и осенью в данном биоценозе наиболее многочислен сенокосец N. lugubris .
Суммарное видовое разнообразие паукообразных кипрейно-вейниковой гари (ПЗП-20) незначительно отличается от такового ПЗП-2. Здесь было обнаружено 62 вида пауков и 5 видов сенокосцев (табл. 1). Как и на малиново-кипрейно-вейниковой гари максимальное количество видов зафиксировано в летний период. Однако осенний аспект населения кипрейно-вейниковой гари характеризуется заметным сокращением видового разнообразия пауков.
В сезонной динамике попадаемости пауков наблюдается летний пик, приходящийся на вторую половину июня и менее выраженный осенний подъем (рис. 1Б). Максимальная попадаемость, зафиксированная во второй декаде июня, составила 237 экз./100 лов. сут. В весенний период почти треть пойманных пауков представлена пауком-балдахинником T. setosus , кроме того, многочисленны пауки-волки A. pulverulenta и неполовозре-
лые Pardosa . Обильные весной виды летом редки, а основу населения герпетобионтных пауков составляют пауки-волки – P. fulvipes и P. lugubris , и гнафозида H. soerenseni (табл. 1). В населении пауков данного биотопа осенью преобладают те же два вида пауков-волков, которые были многочисленны летом, и паук-балдахинник T. mengei . По-падаемость сенокосцев низкая и незначительно меняется в течение сезона.
Население лесов
Фауна паукообразных березняка вейникововысокотравного (ПЗП-7) характеризуется относительно низким видовым разнообразием. Здесь обнаружено 45 видов: 39 пауков и 5 сенокосцев (табл. 2). Максимальное видовое разнообразие пауков, как и на гарях, наблюдается в летний период. Однако летний пик более выражен – количество видов летом в 1.5 раза больше, чем весной и осенью (табл. 2). Видовое разнообразие сенокосцев увеличилось от весны к осени с 1 до 5 видов (табл. 2).
Весной и в первой половине лета попадаемость пауков была стабильно низкой, рост попадаемости начинается со второй декады июля (рис. 1В). Во второй половине августа зафиксирован максимум попа-даемости – 293 экз./100 лов. сут. Такая динамика объясняется появлением и увеличением численности в этот период осенних лесных видов из сем. Linyphiidae: в основном, A. scopigera и, в меньшей степени, C. sylvaticus и T. tenebricola .
В пихто-ельнике высокотравно-папоротниковом коренном (ПЗП-19) отмечено 55 видов пауков и 5 видов сенокосцев (табл. 2). Количество видов пауков весной в 2.5 раза ниже такового летом и осенью. Динамика видового разнообразия сенокосцев имеет аналогичные тенденции.
Попадаемость пауков в течение сезона относительно равномерно увеличивается (рис. 1Г). Максимальное значение этого показателя, 268 экз./100 лов. сут., зафиксировано во второй половине августа. Как и в березняке, такой тип динамики определяют пауки-балдахинники, из которых наиболее многочислен A. scopigera , а осенью – A. olivacea и, в меньшей степени, C. sylvaticus, P. alutacius и T. tenebricola . Весной в этом биотопе обычны пауки-балдахинники I. kochiel-lus, O. angulatus и T. alacris .
Попадаемость сенокосцев в ельнике самая низкая из всех обследованных биоценозов и незначительно варьирует в летне-осенний период.
Обсуждение
До 2017 г. исследования структуры населения пауков и сенокосцев ветровальных и гаревых сообществ затрагивали поздне-летний и осенний периоды [Есю-нин, Ефимик, Мазура, 2000; Есюнин и др., 2001; Есюнин, Шумиловских, 2003; Ухова, Есюнин, 2016] или поздне-весенний и ранне-летний аспекты [Ухова и др., 2014]. Приведенные выше данные впервые позволяют сравнить все аспекты населения паукообразных по материалам, собранным в течение одного вегетационного сезона.
Весенний аспект . Во всех исследованных биотопах весенние группировки пауков и, особенно, сенокосцев характеризуются низкой попадаемо-стью (табл. 3). С одной стороны, это связано с низкой численностью, а с другой – пониженной уловистостью.
Низкая попадаемость в ловушки личиночных стадий пауков и сенокосцев на первых этапах их жизни обусловлена тем, что они, как правило, обитают в подстилке, т.е. являются стратобионтами. И только позднее, по мере увеличения размеров, меняют среду обитания, становясь компонентом гер-петобионтного комплекса. Другая общая закономерность – небольшое количество видов и отсутствие, за редким исключением, обильных видов. Выравненность численностей видов в весенних группировках пауков отражается в высоких значениях интегрированного показателя разнообразия (табл. 3: индекс Шеннона).
Таблица 3
Основные показатели весеннего аспекта населения пауков и сенокосцев на пробных зоологических площадках Висимского заповедника в 2017 г.
|
Показатель |
Биотоп |
|||
|
Гари |
Березняк |
Пихто-ельник |
||
|
ПЗП-2 |
ПЗП-20 |
ПЗП-7 |
ПЗП-19 |
|
|
Средняя попадаемость пауков (экз./100 лов.сут.) |
39.7 |
67.5 |
20.8 |
31.3 |
|
Кол-во видов пауков |
18 |
16 |
18 |
14 |
|
Индекс разнообразия (Шеннона, H’) |
2.96 |
2.49 |
3.10 |
2.94 |
|
Доля Linyphiidae (% от всех особей) |
55 |
46 |
72 |
70 |
|
Доля Lycosidae (% от всех особей) |
24 |
48 |
12 |
13 |
|
Средняя попадаемость сенокосцев (экз./100 лов.сут.) |
2.4 |
1.7 |
2.5 |
1.0 |
|
Кол-во видов сенокосцев |
2 |
1 |
1 |
1 |
|
Доминирующие виды |
- |
- |
- |
- |
|
Alopecosa pulverulenta (Clerck, 1757) |
- |
IV |
- |
- |
Примечание. IV – обильный вид [Песенко, 1982].
Причем значения индекса сопоставимы с таковыми многовидовых летних группировок (табл. 4). По прочим показателям гаревые и лесные группировки существенно различаются. Попадаемость пауков на гарях весной выше, чем в лесах (табл. 3). Различаются соотношения семейств в сообществах пауков. На гарях пауки-балдахинники (Linyphiidae) составляют около половины всех отловленных особей, тогда как в лесах доля линифи-ид не менее 70%. Обратную тенденцию демонстрируют пауки-волки (табл. 3), доля которых в населении пауков гари в два и более раз больше, чем в лесах.
Летний аспект. Увеличение попадаемости пауков и сенокосцев в этот период сопровождается ростом количества видов (табл. 4). Летнее население характеризуется наибольшим за сезон количеством видов. Это обусловлено тем, что в населении одновременно присутствуют представители трех фенологических групп – весенне-летние, летние и позднелетне-осенние виды.
Таблица 4
Основные показатели летнего аспекта населения пауков и сенокосцев на пробных зоологических площадках Висимского заповедника в 2017 г.
|
Показатель |
Биотоп |
|||
|
Гари |
Березняк |
Пихто-ельник |
||
|
ПЗП-2 |
ПЗП-20 |
ПЗП-7 |
ПЗП-19 |
|
|
Средняя попадаемость пауков (экз./100 лов.сут.) |
146.4 |
147.0 |
46.8 |
72.0 |
|
Кол-во видов пауков |
48 |
47 |
28 |
37 |
|
Индекс разнообразия (Шеннона, H’) |
2.96 |
2.30 |
2.46 |
3.11 |
|
Доля Linyphiidae (% от всех особей) |
28 |
13 |
75 |
80 |
|
Доля Lycosidae (% от всех особей) |
53 |
72 |
11 |
6 |
|
Средняя попадаемость сенокосцев (экз./100 лов.сут.) |
15.0 |
2.3 |
21.7 |
8.7 |
|
Кол-во видов сенокосцев |
4 |
4 |
3 |
4 |
|
Доминирующие виды |
||||
|
Allomengea scopigera (Grube, 1859) |
- |
- |
V |
- |
|
Alopecosa pulverulenta (Clerck, 1757) |
- |
IV |
- |
- |
|
Haplodrassus soerenseni (Strand, 1900) |
- |
- |
IV |
- |
|
Agyneta allosubtilis (Loksa, 1965) |
IV |
- |
- |
- |
|
Pardosa fulvipes (Collett, 1876) |
- |
IV |
- |
- |
|
Pardosa lugubris (Walckenaer, 1802) |
IV |
- |
- |
- |
|
Pardosa riparia (C. L. Koch, 1833) |
IV |
V |
- |
- |
|
Piratula hygrophila (Thorell, 1871) |
IV |
- |
- |
- |
|
Lacinius ephippiatus (C.L. Koch, 1835) |
IV |
- |
IV |
- |
|
Mitopus morio (Fabricius, 1779) |
- |
- |
V |
IV |
|
Oligolophus tridens (C. L. Koch, 1836) |
- |
- |
IV |
- |
|
Nemastoma lugubris (O.F. Müller, 1776) |
V |
- |
- |
- |
Примечание. IV – обильный и V – очень обильный вид [Песенко, 1982].
В летний период в наибольшей степени выражены различия в структуре населения изученных группировок. Динамика попадаемости пауков на гарях имеет четко выраженный пик (см. рис. 1А,
Б), тогда как в лесах наблюдается равномерное увеличение этого показателя в течение лета (см. рис. 1В, Г). Доминантный комплекс гаревых группировок пауков многовидовой, тогда как в лесах имеется только пара обильных видов, или виды представлены небольшим количеством особей. Особенность доминантного комплекса лесных группировок пауков обусловлена тем, что численность личинок основного доминанта A. scopigera летом еще не слишком велика.
Так или иначе, но выравненность летнего аспекта населения пауков высокая, что отражается в максимальных за сезон значениях индексов видового разнообразия (табл. 4: индекс Шеннона). Летом гаревые группировки существенно различаются друг от друга по составу доминантных видов пауков и численности сенокосцев. Несмотря на то, что в обоих типах гари возрастает численность пауков-волков, на кипрейно-вейниковой гари (ПЗП-
Таблица 5
Основные показатели осеннего аспекта населения пауков и сенокосцев на пробных зоологических площадках Висимского заповедника в 2017 г.
|
Показатель |
Биотоп |
|||
|
Гари |
Березняк |
Пихто-ельник |
||
|
ПЗП-2 |
ПЗП-20 |
ПЗП-7 |
ПЗП-19 |
|
|
Средняя попадаемость пауков (экз./100 лов. сут.) |
117.7 |
76.3 |
157.1 |
153.5 |
|
Кол-во видов пауков |
40 |
28 |
19 |
38 |
|
Индекс разнообразия (Шеннона, H’) |
2.83 |
2.40 |
0.88 |
2.08 |
|
Доля Linyphiidae (% от всех особей) |
58 |
29 |
98 |
96 |
|
Доля Lycosidae (% от всех особей) |
25 |
52 |
0 |
2 |
|
Средняя попадаемость сенокосцев (экз./100 лов.сут.) |
7.2 |
2.4 |
16.9 |
7.3 |
|
Кол-во видов сенокосцев |
3 |
4 |
5 |
3 |
|
Доминирующие виды |
||||
|
Agyneta allosubtilis (Loksa, 1965) |
- |
- |
- |
V |
|
Allomengea scopigera (Grube, 1859) |
- |
- |
V |
IV |
|
Centromerus sylvaticus (Blackwall, 1841) |
- |
- |
V |
IV |
|
Tenuiphantes mengei (Kulczyński, 1887) |
V |
IV |
- |
- |
|
Pardosa riparia (C. L. Koch, 1833) |
- |
IV |
- |
- |
Примечание. IV – обильный и V – очень обильный виды [Песенко, 1982].
В отличие от других периодов, осенью попада-емость пауков в лесах выше, чем на гарях (табл. 5). Практически все осеннее население пауков лесных биотопов состоит из представителей сем. Linyphiidae (табл. 5) с очень похожим доминантным комплексом видов. Различия между гаревыми группировками пауков, наметившиеся в летний период, осенью хорошо выражены: различны по-падаемость, доли семейств, доминантный комплекс (табл. 5).
Таким образом, по данным, полученным на 7-й год после последнего пожара, структура населения пауков гаревых биотопов, существенно отличается от таковой лесов в течение всего вегетационного сезона (рис. 2).
Описанные выше весенние группировки пауков значительно отличаются от таковых, обследованных четырьмя годами ранее [Ухова и др., 2014].
-
20) этот процесс более выражен. Летом пауки-волки здесь составляют более 70% отловленных особей (табл. 4). В лесах рост видового разнообразия и численности пауков происходит за счет видов из сем. Linyphiidae.
Осенний аспект. Осеннее население пауков характеризуется низким видовым разнообразием: уменьшаются количество видов и выравненность населения, в лесах существенно снижаются значения индексов разнообразия (табл. 5). Такие изменения обусловлены исчезновением видов летней фенологической группы и появлением очень обильных осенних видов. Последнее приводит к тому, что в лесах отмечается максимальная за сезон попадаемость пауков, а на гарях в большей или меньшей степени выражен второй за сезон осенний пик численности. Сенокосцы осенью были малочисленны во всех биотопах.
Изученные в 2013 г. арахнокомплексы в большей степени соответствуют летней структуре населения. В 2013 г. в конце мая – начале июня попадае-мость пауков была очень высокой на всех ПЗП, от 122 до 237 экз./100 лов. сут.; доминантный комплекс видов формировали пауки-волки A. taeniata, P. lugubris, P. fulvipes, P. riparia . Причем два первых вида были многочисленны как на гарях, так и в лесах, что не отмечается в 2017 г. Только население пауков березняка имело типичные лесные черты, т.к. здесь был обильным паук-балдахинник A. olivacea . Ранее мы уже указывали на тот факт, что структура весеннего населения пауков мало пригодна для целей мониторинга, т.к. она очень изменчива и в значительной степени определяется погодой в начале сезона [Есюнин, Шумиловских, 2003].
Для целей мониторинга населения пауков ра- ционально исследовать осенний аспект [Есюнин, Шумиловских, 2003].
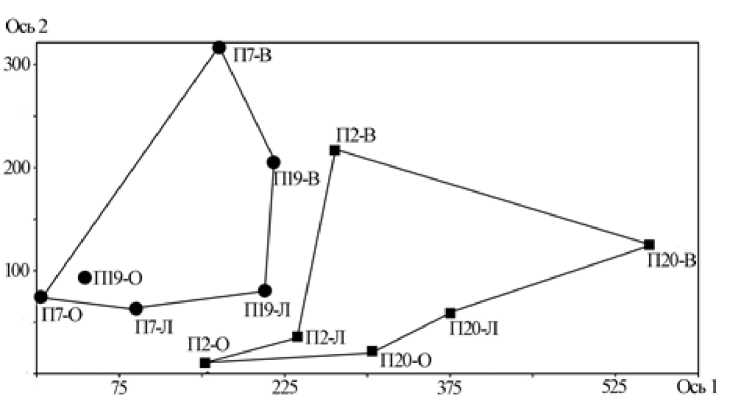
Рис. 2. Биплот анализа соответствия (метод Detrended correspondence analysis) населения пауков гаревых и лесных биотопов Висимского заповедника.
Номера обследованных биотопов: П2 и П20 – малиново-кипрейно-вейниковая и кипрейно-вейниковая гари, соответственно; П7 – елово-березовый лес, П19 – пихто-ельник. Фенологические периоды: В – весенний, Л – летний, О – осенний
Осенние группировки пауков в Приуралье демонстрируют поразительное единообразие структуры, а зачастую и доминантного комплекса в пределах одного типа растительности. Сравнивая приведенные выше результаты с данными, полученными в 2014 г. [Ухова, Есюнин, 2016], мы обнаруживаем следующее. В 2017 г. попадаемость осенью значительно выше: на гарях в среднем в 1.5 раза, в лесах – в 3 раза. Несмотря на это, состав доминантного комплекса демонстрирует одинаковые тенденции. В лесных группировках осенью доминирует A. scopigera , на гарях – пауки-волки, в частности, P. riparia . Однако в 2017 г. во всех биотопах в осеннем населении пауков попадались только единичные особи паука-волка A. taeniata – вида, который был очень многочислен в 2014 г. В заключение можно сделать вывод о том, что через 7 лет, прошедших со времени последнего пожара, сохраняются значительные отличия в структуре населения герпетобионтных пауков гаревых и лесных биоценозов.
Выражаем благодарность лаборанту научного отдела Висимского заповедника В.Д. Араповой и студенту 3 курса Пермского государственного национального исследовательского университета биологического факультета кафедры зоологии беспозвоночных и водной экологии К.С. Фефиловой за оказанную помощь в разборе проб.
Список литературы Структура населения пауков и сенокосцев (Arachnida, Araneae, Opiliones) модельных биоценозов Висимского биосферного заповедника
- Есюнин С.Л., Ефимик В.Е., Мазура Н.С. Структура населения мезофауны подстилки пихтоельников Висимского заповедника после ветровала // Последствия катастрофического ветровала для лесных экосистем: сб. науч. тр. Екатеринбург, 2000. С. 69–86.
- Есюнин С.Л., Козьминых В.О., Ухова Н.Л. Структура и разнообразие первичнопирогенных сообществ на месте коренных пихто-ельников Среднего Урала. 2. Герпетобионтные членистоногие // Вестник Пермского университета. 2001. Вып. 4. Биология. С. 144–153.
- Есюнин С.Л. и др. Динамика изменения структуры и разнообразия герпетобионтных беспозвоночных на травяно-кустарниковой стадии развития гарей пихто-ельников Висимского заповедника // Исследования эталонных природных комплексов Урала: материалы науч. конф. Екатеринбург, 2001. С. 284–294.
- Есюнин С.Л., Ухова Н.Л. Аннотированный список паукообразных (Arachnida) Висимского заповедника // Современное состояние и перспективы развития ООПТ Урала: материалы науч.-практ. конф. Екатеринбург, 2011. С. 124–138.
- Есюнин С.Л., Шумиловских Л.С. Аспектность населения беспозвоночных (замечания к проблемам мониторинга) // Экологические проблемы заповедных территорий России. Тольятти, 2003. С. 183–187.
- Песенко Ю.А. Принципы и методы количественного анализа в фаунистических исследованиях. М.: Наука, 1982. 287 с.
- Сибгатуллин Р.З. 7.2.4. Сукцессионные процессы // Летопись природы Висимского государственного природного биосферного заповедника за 2013 г. М., 2014. С. 70–82.
- Сибгатуллин Р.З. Динамика пихто-ельника высокотравно-папоротникового в Висимском заповеднике // Ботаника в современном мире: тр. XIV съезда РБО. Махачкала, 2018. Т. 2. Геоботаника. Ботаническое ресурсоведение. Интродукция растений. Культурные растения. С. 135–137.
- Ухова Н.Л., Есюнин Л.С. 8.2.3.2. Численность напочвенных беспозвоночных животных // Летопись природы Висимского государственного природного биосферного заповедника за 2014 г. Екатеринбург, 2016. С. 97–104.
- Ухова Н.Л. и др. 8.2.3.1. Численность почвенных и напочвенных беспозвоночных животных // Летопись природы Висимского государственного природного биосферного заповедника за 2013 г. М., 2014. С. 106–132.
- Aitchison-Benell C.W. Resposes to fire by taiga spiders // Proceedings of the Entomological Society of Ontario. 1994. Vol. 125. P. 29–41.
- Atlegrim O., Sjoberg K. Effects of clear-cutting and selective felling in Swedish boreal coniferous forest: response of invertebrate taxa eaten by birds // Annales Entomologicae Fennicae. 1995. Vol. 6. P. 79–90.
- Buddle C.M., Spence J.R., Langor D.W. Succession of boreal forest spider assemblages following wildfire and harvesting // Ecography. 2000. Vol. 23. P. 424–436.
- Coddington J.A., Young L.H., Coyle F.A. Estimating spider species richness in a southern Appalachian Cove hardwood forest // Journal of Arachnology. 1996. Vol. 24. P. 111–128.
- Coyle F.A. Effects of clearcutting on the spider community of a southern Appalachian forest // Journal of Arachnology. 1981. Vol. 9. P. 285–298.
- Gongalskij K.B., Persson T. Recovery of soil macrofauna after wildfires in boreal forests // Soil biology & Biochemistry. 2013. Vol. 57. P. 182−191.
- Gorbunova A.Yu. et al. Forest fires increase variability of soil macrofauna communities along a macrogeographic gradient // European Journal of Soil Biology. 2017. Vol. 80. P. 49–52.
- Hammer Ø., Harper D.A.T., Ryan P.D. PAST: Paleontological statistics software package for education and data analysis // Palaeontologia Electronica. 2001. Vol. 4, № 1. P. 1–9.
- Hatley C.L., Macmahon J.A. Spider community organization - seasonal variation and the role of vegetation architecture // Environmental Entomology. 1980. Vol. 9. P. 632–639.
- Hauge E., Kvamme T. Spiders from forest-fire areas in southeast Norway // Fauna Norvegica. 1983. Vol. B30, № 1. P. 39-45.
- Huhta V. Succession in the spider communities of the forest floor after clear-cutting and prescribed burning // Annales Zoologici Fennici. 1971. Vol. 8. P. 483–542.
- Jennings D.T. et al. Spiders (Araneae) associated with strip-clearcut and dense spruce–fir forests of Maine // Journal of Arachnology. 1988. Vol. 16. P. 55–70.
- Koponen S. Early succession of a boreal spider community after forest fire // Journal of Arachnology. 2005. Vol. 33. P. 230–235.
- Larrivée M., Fahrig L., Drapeau P. Effects of a recent wildfire and clearcuts on ground-dwelling boreal forest spider assemblages // Canadian Journal of Forest Research. 2005. Vol. 35. P. 2575–2588.
- Mordkovich V.G. et al. Soil arthropoda of post-fire successions in northern taiga of West Siberia // Contemporary Problems of Ecology. 2008. Vol. 1, № 1. P. 96–103.
- Moretti M. et al. The effects of wildfire on groundactive spiders in deciduous forests on the Swiss southern slope of the Alps // Journal of Applied Ecology. 2002. Vol. 39. P. 321–336.
- Niklasson M., Granström A. Numbers and sizes of fires, long-term spatially explicit fire history in a Swedish boreal landscape // Ecology. 2000. Vol. 81. P. 1484−1499.
- Nyffeler M. Ecological impact of spider predation: a critical assessment of Bristowe’s and Turnbull’s estimates // Bulletin of the British Arachnological Society. 2000. Vol. 11, № 9. P. 367–373.
- Pajunen T. et al. Ground-dwelling spiders (Arachnida, Araneae) in fragmented old forests and surrounding managed forests in southern Finland // Ecography. 1995. Vol. 18. P. 62–72.
- Samu F. et al. Differential ecological responses of two generalist arthropod groups, spiders and carabid beetles (Araneae, Carabidae), to the effects of wildfire // Community Ecology. 2010. Vol. 11, № 2. P. 129–139.
- Uetz G.W. The influence of variation in litter habitats on spider communities // Oecologia. 1979. Vol. 40. P. 29–42.
- Wardle D.A. et al. Longterm effects of wildfire on ecosystem properties across an island area gradient // Science. 2003. Vol. 300. P. 972−975.
- Wise D.H. Spiders in ecological webs. Cambridge: Cambridge University Press, 1993. 344 p.
