Структура популяции Hypogymnia physodes (L.) Nyl. в условиях гор Центрального Кавказа
 Nyl. в условиях гор Центрального Кавказа")
Автор: Ханов Залим Мирович
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Общая биология
Статья в выпуске: 4-2 т.17, 2015 года.
Бесплатный доступ
В статье рассматривается структура ценопопуляции вида лишайника Hypogymnia physodes в лесах Национального парка «Приэльбрусье» (Кабардино-Балкария). Использование методов описания структуры популяции, основанные на стадии развития и репродуктивном статусе индивидуальных талломов и их группировок (субпопуляций), позволили сделать вывод о текущем состоянии и перспективах развития популяции.
Эпифитные лишайники, популяция, популяционные структуры
Короткий адрес: https://sciup.org/148205709
IDR: 148205709 | УДК: 574.3:582.29
The population structure of Hypogymnia physodes (L.) Nyl. in the mountains of the Central Caucasus
The cenopopulation structure of the lichen species Hypogymnia physodes in the forests of the National Park “Prielbrusye” (Kabardino-Balkaria) is considered. The introduced description methods for the population structure, based on the development stage and reproductive status of individual thalli and their groups (subpopulations), allow to draw a conclusion on the current state and development prospects of the population.
Текст научной статьи Структура популяции Hypogymnia physodes (L.) Nyl. в условиях гор Центрального Кавказа
Лишайники являются одним из важнейших компонентов горных экосистем. В горных фитоценозах лишайники составляют большую часть видового разнообразия.
Одной из наименее разработанных областей лихенологии является популяционная биология лишайников. Лишь в последние двадцать лет рядом зарубежных исследовательских групп проводилось изучение популяционной организации лишайников [1, 2]. В России исследованиями популяционной структуры лишайников занимаются многие исследователи [3, 4, 5, 6, 7, 8]. Работ по изучению структуры ценопопуляций лишайников в Российской части Кавказа до настоящего времени нет.
В современной лихенологии исследования на популяционном уровне организации лихенобио-ты привлекает все большее внимание специалистов. Показано, что исследования численности и структуры популяций лишайников в разных экологических условиях могут пролить свет на причины субстратной и фитоценотической приуроченности лишайников [5, 2, 1], а также на популяционные механизмы устойчивости/чувстви-тельности лишайников к техногенным нагрузкам [4, 6]. Исследования структуры популяций редких и исчезающих видов лишайников являются необходимой основой для прогнозирования их будущего и разработки стратегии охраны [3, 9].
основных подхода к решению этой проблемы. В первом случае за элементарную единицу принимается таллом, который на момент исследования визуально регистрируется как дискретное образование [1, 2, 3, 4, 6]. Естественно, этот подход может быть применен только к видам, образующим дискретные талломы (многие листоватые виды, некоторые накипные и кустистые). Несмотря на существенные ограничения, этот подход на настоящий момент является преобладающим. Последователи второго подхода за элементарную единицу принимают так называемый «функциональный индивидуум», то есть совокупность талломов изучаемого вида, населяющих единицу субстрата (например, ствол дерева) [9]. Этот подход более универсален и может быть применен ко многим эпифитным и эпиксильным лишайникам, у которых трудно выделить индивидуальные талломы. Однако авторы подхода не предложили методов оценки репродуктивного статуса «функциональных индивидуумов»; при построении моделей развития популяции все «функциональные индивидуумы» признаются равнозначными [11].
Необходимо отметить, что единственное на сегодняшний день определение популяции лишайников [12] является «безразмерным»: популяция определяется как локальная группа талломов одного вида, более или менее пространственно отделенная от других таких групп. Совершенно очевидно, что при таком определении фактический объем популяции определяется масштабом и целями исследования: в качестве локальной группы могут выступать талломы, заселяющие один ствол дерева, его крону или отдельную ветвь; талломы в пределах одного фитоценоза или в пределах конкретного более или менее обширного географического региона.
Целью данной работы является оценка потенциала популяции и прогноз перспективы ее развития с помощью методики анализа популяционных структур эпифитных лишайников.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЙ
Исследования проведены на Центральном Кавказе (в пределах Кабардино-Балкарской республики, территория Национального парка «Приэльбрусье»), в поясе остепненных лугов [13], относящемуся Северо-Кавказскому горному району (43º10’N, 42º45’E). Климат умеренно континентальный, годовое количество осадков 791 мм. Среднегодовая температура – 0,2°C, средняя температура января и июля составляет соответственно –17,7°C и +17,0°C.
Пробная площадь №1 (ущ. Сылтран) расположена на высоте 1950 м над ур. м., в средней части южного склона г. Сарыкол (2931 м над ур. м.). Древостой представлен Pinus kochiana Klotzsch ex C. Koch со средним возрастом 56 лет. Сомкнутость крон – 1. В подлеске отмечены Juniperus sabina, J. communis, Berberis vulgaris , Rosa baksanica Почва хорошо дренированная, глубокая и каменистая; классифицирована как горная лесо-луговая [14].
На площадке №2 (ущ. Адыр-Су) древостой двухярусный, образован Pinus kochiana высотой до 26 м. Средний диаметр древостоя 38 см. Насаждение разновозрастное, средний возраст составляет 67 лет. Сомкнутость крон – 0,6. В составе подлеска встречаются Berberis vulgaris L., Sorbus aucuparia L., Juniperus communis, Lonicera steveniana Fisch. et Pojark . Подрост не развит. В травяном покрове преобладают следующие виды сосудистых растений: Geranium sylvaticum L, Lathyrus pratensis L., Moneses uniflora (L.) A. Grey и др., Valeriana tilifolia Troitzk ., Carum meifolium (Bieb.) Boiss, Calamagrostis arundinacea (L.) Roth, Vicia cracca L., Poa nemoralis L., Polygonatum verticillatum (L.) All., Primula macrocalyx Bunge и др. Сосняк разнотравный произрастает на cеверо-восточных склонах на высоте 1960 м над ур. м. Почвы горные серые лесные. Верхний почвенный слой каменистый, супесчаный [14].
Объект исследования – листоватый лишайник Hypogymnia physodes (L.) Nyl. – широко распространен в Европе, Африке, Северной Америке и Азии. Вид обладает средней чувствительностью к любому типу антропогенного воздействия и часто доминирует в эпифитном покрове основных лесообразующих пород [4].
Размножение и распространение H. physodes происходит преимущественно с помощью вегетативных диаспор – соредий, реже фрагментами таллома. Споры полового размножения формируются на старых талломах в апотециях [15].
На участке 10х10 м были зарегистрированы все деревья с H. physodes ( Pinus kochiana ). На каждом дереве выполнено описание всех талломов H. physodes (всего 46 талломов). Для каждого таллома указано его местоположение на стволе (высота и экспозиция), а также на каждом талломе было подсчитано количество соралей каждого из четырех типов [4].
По количеству и степени соралей выделены следующие группы талломов по [4]: 1) асореди-озные (as) – полное отсутствие соралей; 2) крип-тосоредиозные (cs) – все сорали на начальном этапе развития; 3) гипосоредиозные (S1) – сорали на трубчатых и 1-2 на шлемовидных и/или губо-видных лопастях; 4) мезосоредиозные (S2) – до 10 соралей на шлемовидных и/или губовидных лопастях; 5) гиперсоредиозные (S3) – более 10 соралей на шлемовидных и/или губовидных лопастях.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
На данной площадке высота поднятия лишайников по стволу варьирует от 1,5 до 2,5 м. Плотность популяции H. physodes варьирует в широких пределах – 16-60%, и в среднем составляет 32,7133,60 экз./ствол. Общее проективное покрытие ствола лишайниками – 15,00-16,49% (табл. 1).
Размерная структура популяций H. physodes свидетельствует о преобладании небольших талломов с размерами 30-45 мм, лишайники максимальных размеров (более 50 мм) встречаются редко и составляют 6,90 % от их общего числа. Масса талломов в данной популяции колеблется в довольно широких пределах (табл. 2) и в среднем составляет 2,58±0,11.
Из данных таблицы 2 видно, что образцы, собранные в ущ. Сылтран преобладают как по размеру, так и по массе талломов над образцами собранными в ущ. Адыр-Су. Видимо это объясняется тем, что микроклиматические условия ущ. Сылтран для данной популяции благоприятны, что в свою очередь играют большую роль в проведении всех физиологических процессов лишайника.
Важной популяционной характеристикой является возрастное состояние, определяющееся степенью развития соралей, которая показывает вклад в воспроизводство популяции. На начальном этапе развития соралей продукция соредий минимальна [4]. В данной популяции в функционально-возрастном отношении доминируют гиперсоредиозные талломы, редко встречаются мезо- и гипосоредиозные талломы (рис. 1).
Таб. 1. Характеристика эпифитного лишайникового покрова Pinus kochiana в разных условиях среды
|
Показатели |
ущ. Адыр-Су |
ущ. Сылтран |
|
X ± m |
X ± m |
|
|
Общее проективное покрытие, (%) |
16,49±0,43 |
15,00±0,26 |
|
Высота поднятия по стволу, см |
184,00±2,49 |
- |
|
Плотность популяции H. physodes, экз./ствол |
33,60±7,70 |
32,71±5,90 |
Таб. 2. Результаты статистического анализа массы и длины талломов H. physodes в разных условиях среды
|
Параметры |
Длина, мм |
Масса, мг |
||
|
Адыр-Су |
Сылтран |
Адыр-Су |
Сылтран |
|
|
Минимум |
18,91 |
29,56 |
0,50 |
2,70 |
|
Максимум |
58,47 |
184,64 |
6,00 |
3,12 |
|
Среднее арифметическое |
36,91 |
79,19 |
2,58 |
10,25 |
|
Среднее квадратическое отклонение |
8,21 |
32,36 |
1,27 |
6,67 |
|
Стандартная ошибка |
0,88 |
6,60 |
0,13 |
1,36 |
|
Коэффициент вариации, % |
22,25 |
40,86 |
49,21 |
65,14 |
Причем эта функционально-возрастная группа довольно неоднородна по размеру, от 18,91 мм до 58,47 мм. Мезосоредиозные талломы по размерам практически не отличаются от гиперсоре-диозных, 37,45 и 37,57 мм, соответственно. Выше сказанное характеризует популяцию обитающей в ущ. Адыр-Су в целом как зрелую, а гидротермический режим среды как благоприятный (рис. 1).
В другой популяции , обитающей в ущ. Сыл-тран на высоте около 2000 м над ур.м. наблюдается некоторое снижение проективного покрытия H. physodes на стволе (Р≤0,026) по сравнению с таковой из ущ. Адыр-Су, при этом плотность популяции в двух выборках на одном уровне.
Сравнение частотных распределений массы и длины талломов в двух популяциях H. physodes по непараметрическому критерию Краскела-Уоллиса приводит к выводу, что они различаются на высоком уровне значимости (Р≤0,000), что связано с увеличением данных параметров в ущ. Сылтран, по сравнению с выше таковыми в ущ. Адыр-Су. При этом cреди функциональ- но-возрастных групп преобладают лишайники с гиперсоредиозными талломами, как и в популяции из ущ. Адыр-Су. Однако в небольшом количестве – 4,17 % здесь представлены более молодые гипосоредиозные талломы.
Кроме того, мы попытались рассмотреть дальнейший путь развития субпопуляций. В качестве примера, мы основывались на работе [5]. Колонизирующая субпопуляция возникает при успешном прорастании и развитии диаспоры (или нескольких диаспор), занесенных извне. Успешно развивающийся таллом формирует соредии, значительная часть которых попадает на тот же ствол. При условии их успешного прорастания образуется растущая субпопуляция. Если развития соредий не происходит, талломы колонизирующей субпопуляции с возрастом начинают разрушаться с образованием многочисленных более или менее мелких лопастей разного репродуктивного статуса – возникает либо ложнорастущая, либо регрессивная субпопуляция. Растущая субпопуляция переходит в
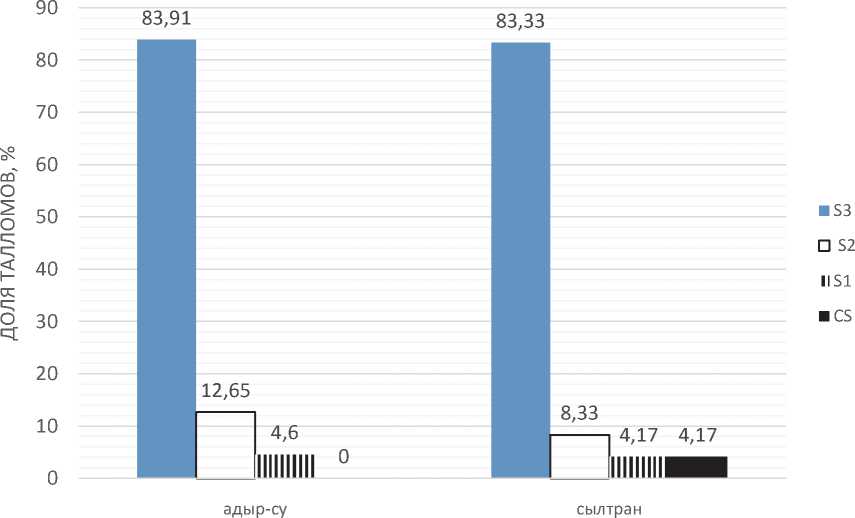
Рис. 1. Возрастной спектр популяций H. physodes в разных условиях среды
стабильную в результате старения талломов и уменьшения свободного пространства на стволе (что препятствует образованию молодых талломов из диаспор). Дальнейшим этапом развития стабильной субпопуляции является ее переход в ложнорастущую – в отличие от показанного выше механизма образования ложнорастущей популяции, этот переход представляется более «благополучным».
Преобладание колонизирующих субпопуляций свидетельствует об относительной биологической молодости популяции лишайников в ущ. Адыр-Су. При этом сам факт успешной колонизации новых субстратов является доказательством достаточно высокого репродуктивного потенциала популяции [5].
В ущ. Сылтран, преобладает ложнорастущая субпопуляция с переходом в стабильное состояние, на что указывает увеличение массы и размера талломов.
Таким образом, нами была впервые проанализирована на начальном этапе структура популяции широко распространенного лишайника H. physodes на территории Национального парка «Приэльбрусье». Было выяснено, что соседние талломы Hypogymnia physodes сливаются в процессе роста с талломами других видов (например, виды рода Parmelia ), однако используют нижерасположенные талломы в качестве точки опоры, в результате чего образуются весьма сложные комплексы. Поэтому чем выше численность субпопуляции, тем большая доля субъективизма присутствует в процессе выделения индивидуальных талломов. Также не свободен от субъективизма и процесс типизации субпопуляций, несмотря на сделанную попытку его формализации путем использования соотношения долей талломов разных функционально-возрастных групп. Это касается главным образом трактовки расположения талломов на стволе.
Тем не менее, анализ и индивидуальных талломов, и субпопуляций свидетельствует об одном – биологической молодости исследуемой популяции. Представляется достаточно вероятным, что при условии отсутствия внешнего вмешательства и катастрофических событий популяция будет способна к дальнейшему росту и колонизации новых субстратов.
Автор выражает искреннюю благодарность сотрудникам лаборатории по мониторингу лесных экосистем Саблировой Ю.М., Пшегусову Р.Х. и Моллаевой М.З. за сбор образцов и консультации при геоботанических описаниях пробных площадок.
Список литературы Структура популяции Hypogymnia physodes (L.) Nyl. в условиях гор Центрального Кавказа
- Rhoades, F.M. Distribution of thalli in a population of the epiphytic lichen Lobaria oregana and a model of population dynamics and production/F.M.Rhoades//Bryologist. 1983. V. 86. -P. 309-331.
- Gauslaa, Y. Population structure of the epiphytic lichen Usnea longissima in a boreal Picea abies canopy/Y.Gauslaa//Lichenologist. 1997. V. 29. -P. 455-469.
- Истомина, Н.Б. Биология Lobaria pulmonaria (L.) Hoffm. и Menegazzia terebrata (Hoffm.) Massal. в южной тайге европейской части России: автореф. дис. канд. биол. наук. 03.00.24/Истомина Нина Борисовна. -М., 1996. -24 с.
- Михайлова, И.Н. Размерная и возрастная структура популяций эпифитного лишайника Hypogymnia physodes (L.) Nyl. в условиях атмосферного загрязнения/И.Н. Михайлова, Е.Л. Воробейчик//Экология. -1999. -№ 2. -С. 130-137.
- Михайлова, И.Н. Анализ субпопуляционных структур эпифитных лишайников (на примере Lobaria pulmonaria (L.) Hoffm. /И.Н. Михайлова//Вест. ННГУ, 2005. № 1 (9). -С. 124-134. -URL: http://www.unn.ru/pages/vestniki_journals/9999-0191_west_bio_2005_1(9)11 (дата обращения 23.07.2015).
- Суетина, Ю.Г. Онтогенез и популяционная структура Xanthoria parietina (L.) Th. Fr. в различных экологических условиях/Ю.Г. Суетина//Экология. -2001. -№ 3. -С. 203-208.
- Плюснин, С.Н. Изменчивость и структура популяций лишайников рода Stereocaulon в тундровых экосистемах: автореферат дис. канд. биолог. наук. 03.02.08/Плюснин Сергей Николаевич. -Сыктывкар, 2004. -23 с.
- Теплых, А.А. Структура популяций лишайника Pseudevernia furfuracea (L.) Zopf в сосняках Республики Марий Эл: автореферат дис. канд. биолог. наук. 03.02.08/Теплых Алексей Александрович. -Казань, 2011. -23 с.
- Scheidegger, C. Monitoring lichens for conservation: Red Lists and conservation action plans/C.Scheidegger, T.Goward, L.Nimis, W.P.Dordrecht//Monitoring with lichens -monitoring lichens. -Netherlands: Kluwer Academic Publishers. 2002. -P. 163-182.
- Гиляров, А.М. Популяционная экология/А.М. Гиляров. М.: Изд-во МГУ, 1990. -191 с.
- Scheidegger, C. Reintroduction and augmentation of populations of the endangered Lobaria pulmonaria: methods and concepts/C.Scheidegger, B.Frey, J.-C.Walser//Lobarion lichens as indicators of the primeval forests of the Eastern Carpathians (Eds. S. Kondratyuk, B. Coppins) (Darwin International Workshop, 25-30 May, Kostrino, Ukraine). -Kostrino, 1998. -P. 33-52.
- Fahselt, D. Individuals, populations and population ecology/D.Fahselt//Lichen biology -Cambridge, 1996. -P. 181-198.
- Соколов, В.Е. Позвоночные Кавказа. Млекопитающие. Насекомоядные/В.Е. Соколов А.К. Темботов. -М.: Наука, 1989. -547 с.
- Молчанов, Э.Н. Почвенная карта Кабардино-Балкарской АССР/Э.Н.Молчанов, В.Д. Калмаков, А.К. Романова. -Новосибирск: Роскартография, 1984.
- Определитель лишайников СССР. Вып. 1. Пертузариевые. Леканоровые. Пармелиевые. Л.: Наука, 1971. -412 с.
