Структура популяций гриба Puccinia triticina на сортах и селекционных линиях мягкой пшеницы на опытном поле Омского ГАУ в 2013-2015 гг

Автор: Гультяева Елена Ивановна, Шаманин Владимир Петрович, Шайдаюк Екатерина Львовна, Потоцкая Инна Владимировна, Игнатьева Алена Игоревна
Журнал: Вестник Омского государственного аграрного университета @vestnik-omgau
Рубрика: Сельскохозяйственные науки
Статья в выпуске: 2 (22), 2016 года.
Бесплатный доступ
Проведен мониторинг вирулентности популяций гриба P. triticina, собранных на опытном поле Омского ГАУ с районированных и перспективных сортов и селекционных линий мягкой пшеницы в 2015 г. Все тестированные изоляты были авирулентны на линиях Tc Lr19 и Tc Lr24. Вариабельность по типу инфекции наблюдали на линиях Тс Lr2a, Тс Lr9, Тс 14b, Тс Lr16, Тс Lr20 и Тс Lr26. Среди изученных 210 монопустульных изолятов выявлено 8 фенотипов вирулентности. Наиболее представлены фенотипы TGR, TQR, THR, различающиеся вирулентностью к генам Lr9 и Lr26. Генетические различия по виру- лентности между популяциями, собранными с разных образцов пшеницы, незначительны, за исключени- ем популяции с сорта Столыпинская. В результате многолетнего анализа омских популяций (2013-2015) выявлена их высокая вирулентность (частота 80-100 %) к генам Lr1, Lr2a, Lr3a, Lr3bg, Lr3ka, Lr10, Lr11, Lr14a, Lr14b, Lr15, Lr16, Lr17, Lr18, Lr30 и авирулентность к генам Lr19, Lr24, Lr41, Lr42, Lr51, Lr53, Lr64, Lr65. Существенный полиморфизм по вирулентности отмечен на линиях Тс Lr9, Тс Lr26 и Тс Lr20. Две группы фенотипов TGQ и TGR, THQ и THR, различающиеся вирулентностью к гену Lr20, превали- ровали во все годы исследований. Фенотип TQR, авирулентный на линиях Tc Lr19, Tc Lr24 и Tc Lr26, встречался ежегодно с частотой от 19,2 до 10 %. В целом не выявлено существенных изменений в струк- туре популяций на опытном поле Омского ГАУ в течение трехлетнего периода.
Мягкая пшеница, бурая ржавчина, вирулентность, lr-гены
Короткий адрес: https://sciup.org/142199177
IDR: 142199177 | УДК: 632.911.2
Structure of P. triticina fungus populations on varieties and breeding lines of bread wheat on Omsk SAU experimental field in 2013-2015
Monitoring of virulence of fungus P. triticina populations collected on Omsk SAU experimental field, on released and promising varieties and breeding lines of bread wheat in 2015 was performed. All tested isolates were avirulent on Thatcher near-isogenic lines (Tc) Lr19 and Lr24. The variability of the infection type on lines TsLr2a, TsLr9, Ts14b, TsLr16, TsLr20 and TsLr26 was observed. Among 210 analysed monopustuled isolates 8 virulence phenotypes were revealed. Phenotypes TGR, TQR, THR with different virulence to genes Lr9 and Lr26 were the most represented. In general, genetic differences on virulence between populations collected from different wheat entries were not significant with exception of variety Stolypinskaya population. As a result of durable analysis of Omsk populations (2013-2015), their high virulence (frequency 80-100%) to genes Lr1, Lr2a, Lr3a, Lr3bg, Lr3ka, Lr10, Lr11, Lr14a, Lr14b, Lr15, Lr16, Lr17, Lr18, Lr30 and avirulence to genes Lr19, Lr24, Lr41, Lr42, Lr51, Lr53, Lr64, Lr65 were detected. A significant polymorphism on virulence on lines TsLr9, TsLr26 and TsLr20 was observed. Two phenotype groups TGQ and TGR, THQ and THR with different virulence to gene Lr20 were predominated for all years of research. Phenotype TQR with avirulence on lines TcLr19, TcLr24 and TcLr26 was observed annually with frequency of 19.2 to 10%. In general, significant changes in the structure of populations on Omsk SAU experimental field were not revealed during three years.
Текст научной статьи Структура популяций гриба Puccinia triticina на сортах и селекционных линиях мягкой пшеницы на опытном поле Омского ГАУ в 2013-2015 гг
Июнь, 2012 [Электронный ресурс]. URL : (Дата обращения 12.02.2016).
Иммуногенетическая защита мягкой пшеницы от вредоносных болезней является базовой составляющей ресурсосберегающих и экологически чистых технологий производства зерна. Для осуществления успешной иммуногенетической защиты необходима информация о расовом составе популяций фитопатогенов в разных регионах РФ и его изменениях, в том числе обусловленных возделыванием новых сортов пшеницы [1].
На опытном поле Омского государственного аграрного университета (Омский ГАУ) ежегодно проводится иммунологическая оценка изучаемых сортов и селекционных линий по устойчивости к комплексу болезней, в том числе и к актуальному в регионе заболеванию – бурой ржавчине. С 2013 г. эти исследования дополнены анализом вирулентности популяций гриба Puccinia triticina Erikss. В результате выявлены высокоэффективные в регионе ювенильные Lr -гены и охарактеризована вирулентность и расовый состав популяций возбудителя на разных сортах и селекционных линиях мягкой пшеницы [2, 3].
Цель работы – продолжение мониторинга вирулентности популяций гриба P. triticina на опытном поле Омского ГАУ и оценка изменчивости структуры популяций в 2013–2015 гг.
Объекты и методы
Листья пшеницы с урединиопустулами были собраны с 21 образца пшеницы, выращиваемого на опытном поле Омского ГАУ в 2015 г. в условиях естественного инфекционного фона. Источником инфекции служили: 1) сорта яровой пшеницы, рекомендованные для выращивания в Западно-Сибирском регионе – Терция (с 1995 г.), Памяти Азиева (с 2001 г.), Дуэт (с 2003 г.), Сибаковская юбилейная (с 2010 г.), Омская 35 (с 2004 г.), Омская 36 (с 2007 г.), Павлоградка (с 2015 г.); 2) находящиеся в Госсортоиспытании Столыпинская (с 2014 г.) и Элемент 22 (с 2015 г.), созданные в Омском ГАУ; 3) перспективные селекционные линии, изучаемые в рамках международного сотрудничества КАСИБ – Лютесценс 141/03-2, Лютесценс 90-12, Лю-тесценс 12/93-01-4, Лютесценс 15-12, Лютесценс 7/04-26, Лютесценс 220/03-83, Лютесценс 106-11, Лютесценс 1101-12, Лютесценс 96-12, Лютесценс 27-12, Астана 2, Фитон 82. Степень развития болезни на данных сортах и линиях составляла от 5 до 80 %.
Популяции с сухих листьев были реанимированы на восприимчивом сорте Инна. С использованием сборной омской популяции P. triticina оценена эффективность генов Lr1 , Lr2a, Lr2b, Lr2c, Lr3a, Lr3bg, Lr3ka, Lr9, Lr10, Lr11, Lr14a, Lr14b, Lr15, Lr16, Lr17, Lr18, Lr19, Lr20, Lr21, Lr24, Lr26, Lr28, Lr29, Lr30, Lr41, Lr42, Lr44, Lr45, Lr47, Lr48, Lr49, Lr51, Lr53, Lr57, Lr64, Lr65 и Lr67 . Для определения фенотипического (расового) состава популяции были клонированы и из каждой получено и размножено по 10 монопустульных изолятов. Для обозначения фенотипов использована буквенная номенклатура [4], основанная на определении вирулентности к группам Lr -линий. Первые две группы в наших исследованиях соответствовали классическому набору (группа 1: Lr1, Lr2a, Lr2c, Lr3а ; группа 2: Lr9, Lr16, Lr24, Lr26 ); а в третью были включены линии с генами отмеченной дифференциацией омских изолятов P. triticina : Lr14b , Lr18 , Lr19 , Lr20 .
Для размножения инфекционного материала, получения монопустульных изолятов и анализа вирулентности использовали методы лабораторного культивирования гриба на отрезках листьев пшеницы, помещенных в раствор бензимидазола [5]. Для сравнения результатов с использованием отрезков листьев по 3 изолята с каждой популяции дополнительно протестированы на интактных растениях. Все изоляты охарактеризованы по признаку виру- лентности. Тип реакции определяли по шкале E.B. Mains, H.S. Jackson [6]. Для определения буквенного кода фенотипов, вычисления индексов внутрипопуляционного разнообразия и различий между популяциями по вирулентности использовали пакет программ Virulence Analysis Tool (VAT) [7]. Для построения UPGMA-дендрограммы – пакет программ NTSYSpc, Version 2.2 (Exeter Software, Setauket, NY).
Результаты и их обсуждение
При инокуляции сборной омской популяцией P. triticina в 2015 г. высокая эффективность была характерна для генов Lr19, Lr24, Lr28, Lr41, Lr42, Lr45 , Lr51, Lr53, Lr57, Lr64 и Lr65 . При анализе монопустульных изолятов вариабельность по вирулентности наблюдали на линиях Thatcher (Тс) c генами Lr2a, Lr9, Lr14b, Lr16, Lr18, Lr20 и Lr26 . Все тестируемые изоляты были авирулентны на линиях Тс Lr19 , Тс Lr24 и вирулентны на Тс Lr1 , Тс Lr2с , Тс Lr3а (табл. 1) . Результаты оценок отрезков листьев и интактных растений идентичны. У изолятов, выделенных с сорта Столыпинская, установлено меньшее число аллелей вирулентности по сравнению с другими изученными популяциями.
Среди изученных 210 монопустульных изолятов выявлено 8 фенотипов (рас) (табл. 2). Наиболее представлены родственные фенотипы TGR, TQR, THR, различающиеся вирулентностью к генам Lr9 и Lr26 . Фенотипы группы Р-, авирулентные к гену Lr2а, зафиксированы в популяциях с сортов Столыпинская и Элемент 22. Индексы генетических различий (Нея N , Роджерса R , Космана, KBm ) незначительно варьировали между изученными популяциями, что указывало на их определенное сходство, за исключением популяций с сортов Столыпинская и Элемент 22. UPGMA-дендрограмма генетического сходства по вирулентности между омскими популяциями по индексу Нея представлена на рис. 1.
Таблица 1
Вирулентность P. triticina на опытном поле Омского ГАУ в 2015 г. (%)
|
Популяции P. triticina с образцов пшеницы |
Лин ии Thatch er c геном |
||||||||
|
Lr19, Lr24 |
Lr2а |
Lr9 |
Lr14b |
Lr16 |
Lr18 |
Lr20 |
Lr26 |
Lr1, Lr2c, Lr3a |
|
|
Дуэт, Терция |
0 |
100 |
100 |
100 |
100 |
100 |
100 |
0 |
100 |
|
Сибаковская юбилейная |
0 |
100 |
100 |
70 |
100 |
100 |
70 |
0 |
100 |
|
Столыпинская |
0 |
0 |
80 |
100 |
90 |
0 |
100 |
0 |
100 |
|
Элемент 22 |
0 |
0 |
0 |
100 |
100 |
100 |
100 |
100 |
100 |
|
Фитон 82 |
0 |
100 |
60 |
100 |
100 |
100 |
100 |
0 |
100 |
|
Л27-12 |
0 |
100 |
50 |
100 |
100 |
100 |
100 |
0 |
100 |
|
Л12/93-01-4 |
0 |
100 |
0 |
100 |
100 |
100 |
100 |
60 |
100 |
|
Л220/03-83 |
0 |
100 |
30 |
100 |
100 |
100 |
100 |
0 |
100 |
|
Памяти Азиева, Л90-12, Л106-11, Л7/04-26 |
0 |
100 |
0 |
100 |
100 |
100 |
100 |
100 |
100 |
|
Омская 35, Омская 36, Астана, Павлоградка, Л15-12, Л141/03-2, Л1101-12, Л96-12 |
0 |
100 |
0 |
100 |
100 |
100 |
100 |
0 |
100 |
Таблица 2
Фенотипический состав P. triticina на опытном поле Омского ГАУ в 2015 г. (%)
|
Популяции P. triticina с образцов пшеницы |
Фенотип |
|||||||
|
TQR |
TQG |
TGR |
THR |
PQL |
PGL |
PBL |
PHR |
|
|
Дуэт, Терция |
100 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
Сибаковская юбилейная |
70 |
30 |
0 |
0 |
0 |
0 |
0 |
0 |
|
Столыпинская |
0 |
0 |
0 |
0 |
80 |
10 |
10 |
0 |
|
Элемент 22 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
100 |
|
Фитон 82 |
60 |
0 |
40 |
0 |
0 |
0 |
0 |
0 |
|
Л27-12 |
50 |
0 |
50 |
0 |
0 |
0 |
0 |
0 |
|
Л12/93-01-4 |
0 |
0 |
40 |
60 |
0 |
0 |
0 |
0 |
|
Л220/03-83 |
30 |
0 |
70 |
0 |
0 |
0 |
0 |
0 |
|
Памяти Азиева, Л90-12, Л106-11, Л7/04-26 |
0 |
0 |
0 |
100 |
0 |
0 |
0 |
0 |
|
Омская 35, Омская 36, Астана, Павлоградка, Л96-12, Л15-12, Л141/03-2, Л1101-12 |
0 |
0 |
100 |
0 |
0 |
0 |
0 |
0 |
0.40 0.35
0.30 0.25 0.20 0.15
Nei distance N
Fiton L27-12 L220/03-83
Astana L141/03-2 L1101-12 Omskaya35 Omskaya36 L96-12 Pavlogradka L15-12
Duet
1 Terciya Sibakovskayaj. P.Azieva L106-11 L7/04-26 L90-12 L12/93-01-4
Element
Stolupinskaya 0.10 0.05 0.00
Рис. 1. UPGMA-дендрограмма генетического сходства омских популяций P. triticina по вирулентности в 2015 г.
В результате многолетнего анализа омских популяций (2013–2015) выявлена их высокая вирулентность (80–100 %) к генам Lr1, Lr2a, Lr3a, Lr3bg, Lr3ka, Lr10, Lr11, Lr14a, Lr14b, Lr15, Lr16, Lr17, Lr18 , Lr30 и авирулентность к генам Lr19, Lr24, Lr41, Lr42, Lr51, Lr53, Lr64, Lr65. Существенный полиморфизм по вирулентности во все годы исследований наблюдали на линиях Тс Lr9, Тс Lr26 и Тс Lr20 (рис. 2). В 2013 г. варьирование по частотам вирулентности также отмечено на линиях с генами Lr44, Lr50 и LrW(52) [2], однако в последующий период установлены умеренно устойчивый тип реакции (балл 2; 2+) на линии с геном Lr50 и восприимчивый (3–4 балла), или гетерогенный (балл 2–3, Х), на линиях с генами Lr44 и LrW (52).
Гены Lr9, Lr20 и Lr26 распространены в сортах пшеницы, выращиваемых в Западной Сибири, и могут оказывать определенное влияние на естественный отбор по вирулентности. Вирулентность к гену Lr9 преимущественно отмечена на сортах – его носителях. На восприимчивых сортах пшеницы без гена Lr9 вирулентные изоляты либо отсутствовали, либо встречались с более низкой частотой. До недавнего времени ген Lr9 относился к группе высокоэффективных в России [8]. Однако массовое его использование при создании сортов в Западной Сибири и на Урале в 2000–2010 гг. предопределило появление вирулентных изолятов [9]. Гены Lr26 и Lr20 относятся к группе неэффективных в России, вирулентность к ним варьирует как между регионами, так и в пределах популяций в одном регионе [8, 10]. Аналогичные различия по частотам вирулентности к генам Lr26 и Lr20 наблюдали в омских популяциях, собранных с разных сортов пшеницы (табл. 1), и в популяциях с одного сорта (рис. 3).
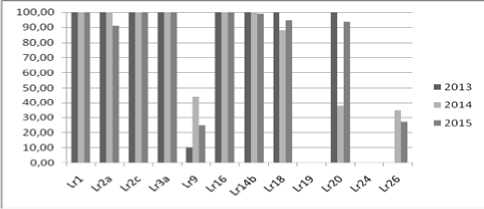
Рис. 2. Вирулентность P. triticina на опытном поле Омского ГАУ в 2013–2015 гг. (%)

Рис. 3. Вирулентность P. triticina к генам Lr20 и Lr26 на сортах Павлоградка, Памяти Азиева и Терция на опытном поле Омского ГАУ в 2013–2015 гг. (%)
Например, инфекционный материал с сортов Памяти Азиева, Терция, Павлоградка ежегодно анализировали и наблюдали варьирование по вирулентности к генам Lr26 и Lr20 на сортах Памяти Азиева и Терция, и его отсутствие – на сорте Павлоградка (рис. 3). Две группы фенотипов TGQ и TGR, THQ и THR, различающиеся вирулентностью к гену Lr20, превалировали в омских популяциях во все годы исследований (табл. 3).
Таблица 3
Фенотипический состав P. triticina на опытном поле Омского ГАУ в 2013–2015 гг. (%)
|
Фенотип |
Авирулентность на Tc Lr -линиях* |
2013 г. |
2014 г. |
2015 г. |
|
PBL |
Lr2a, Lr9, Lr16, Lr18, Lr19, Lr20, Lr24, Lr26 |
0,5 |
||
|
PGL |
Lr2a, Lr9, Lr18, Lr19, Lr20, Lr24, Lr26 |
0,5 |
||
|
PHR |
Lr2a, Lr9, Lr19, Lr24 |
4,8 |
||
|
PQL |
Lr2a, Lr18, Lr19, Lr20, Lr24, Lr26 |
3,8 |
||
|
TGQ |
Lr9, Lr19, Lr20, Lr24, Lr26 |
20,8 |
||
|
TGR |
Lr9, Lr19, Lr24, Lr26 |
90 |
47,6 |
|
|
THQ |
Lr9, Lr19, Lr20, Lr24, Lr26 |
10 |
||
|
THR |
Lr9, Lr19, Lr24 |
25 |
21,9 |
|
|
TQG |
Lr14b, Lr19, Lr20, Lr24, Lr26 |
1,4 |
||
|
TQL |
Lr18,Lr19, Lr20, Lr24, Lr26 |
12,5 |
||
|
TQQ |
Lr19, Lr20, Lr24, Lr26 |
19,2 |
||
|
TQR |
Lr19, Lr24, Lr26 |
10 |
12,5 |
19,5 |
*Определение фенотипов проведено с использованием 12 линий-дифференциаторов: Тс Lr1, Тс Lr2a, Тс Lr2c, Тс Lr3а, Тс Lr9, Тс Lr14b , Тс Lr16, Тс Lr18 , Тс Lr19 , Тс Lr20 , Тс Lr24 и Тс Lr26.
Фенотип TQR, авирулентный на линиях Tc Lr19 , Tc Lr24 и Tc Lr26 , встречался ежегодно с частотой от 19,2 до 10 %. Генетические различия по вирулентности согласно индексу Нея N были выше (0,06) между популяциями, собранными в 2013 г. и 2014 г., и ниже между популяциями 2013 г. и 2015 г. (0,01), 2014 г. и 2015 г. (0,04). Аналогична картина и по фенотипическому составу. Индекс Роджерса R составил 0,9, 0,42 и 0,66 соответственно.
В первую очередь это было обусловлено низкой частотой встречаемости изолятов, вирулентных к гену Lr26 в 2013 г. Несмотря на это, в целом не выявлено существенных изменений в структуре популяций на опытном поле Омского ГАУ в течение трехлетнего периода.
Заключение
В результате многолетнего мониторинга вирулентности популяций гриба P. triticina на сортах яровой пшеницы, выращиваемых на опытном поле ОмГАУ, выявлены эффективные Lr -гены и охарактеризован фенотипический состав. Выделенные эффективные гены Lr24, Lr41, Lr42, Lr51, Lr53, Lr64, Lr65 могут представлять потенциал для использования в качестве доноров устойчивости.
Несмотря на разный уровень полиморфизма омских популяций в 2013–2015 гг., не выявлено существенных различий по вирулентности и фенотипическому составу за трехлетний период. Вирулентность к гену Lr9 преимущественно встречалась на сортах – носителях этого гена и реже на других анализируемых сортах. Частоты вирулентности к другим Lr -генам варьировали как по годам исследований, так и в пределах одного года между анализируемыми сортами пшеницы. Включение в анализ большего количества сортов-источников инфекционного материала предопределило более высокое разнообразие популяций по вирулентности.
Список литературы Структура популяций гриба Puccinia triticina на сортах и селекционных линиях мягкой пшеницы на опытном поле Омского ГАУ в 2013-2015 гг
- Интегрированная защита озимой пшеницы/В.А. Павлюшин //Защита и карантин растений. -2015. -№ 5. -С. 38-71.
- Вирулентность гриба Puccinia triticina на сортах и селекционных линиях мягкой пшеницы на опытном поле OмГАУ в 2013 г./В.П. Шаманин //Вестн. Алт. ГАУ. -2014. -№ 6. -С. 36-42.
- Мониторинг вирулентности популяций гриба Р. triticina на опытном поле ОМГАУ/В.П. Шаманин //Вестн. Алт. ГАУ. -2015. -№ 5 (127). -С. 70-75.
- North American system of nomenclature for Puccinia recondita f.sp. tritici/D.L. Long, J.A. Kolmer//Phytopathology. -1989. -Vol. 79. -P. 525-529.
- Михайлова, Л.А. Методы исследования генетического разнообразия популяций возбудителя бурой ржавчины пшеницы Puccinia reconditа Rob.ex Desm. f. sp. tritici/Л.А. Михайлова, Е.И. Гультяева, Н.В. Мироненко. -СПб.: РАСХН, ВНИИЗР, Инновац. центр защиты растений, 2003. -24 с.
- Physiologic specialization in the leaf rust of wheat; Puccinia triticina Erikss./E.B. Mains, H.S. Jackson//Phytopathology. -1926. -Vol. 16. -P. 89-120.
- Virulence Analysis Tool (VAT)/E. Коsman //User Manual. -2008.
- Гультяева, Е.И. Методы идентификации генов устойчивости пшеницы к бурой ржавчине с использованием ДНК-маркеров и характеристика эффективности Lr-генов/Е.И. Гультяева.-СПб.: РАСХН, отделение защиты растений, ГНУ ВНИИЗР, 2012. -72 с.
- Вирулентность патотипов возбудителя бурой ржавчины пшеницы к ТhLr9 в регионах Сибири и Урала/Л.В. Мешкова //Современные проблемы иммунитета растений к вредным организмам: материалы 2-й Всерос. конф. 29 сентября -2 октября 2008 г. -СПб., 2008. -С. 70-73.
- Cтруктура российских популяций гриба Puccinia triticina Eriks/Е.И. Гультяева //Вестн. защиты растений. -2015. -Вып. 85. -№ 3. -С. 5-10.
