Структура популяций Hedysarum gmelinii Ledeb. на западной границе и в центральной части ареала

Автор: Ильина В.Н.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Наземные экосистемы
Статья в выпуске: 1 т.9, 2007 года.
Бесплатный доступ
Обобщены данные мониторинга ценопопуляций копеечника Гмелина (Hedysarum gmelinii Ledeb.) в бассейне Средней Волги и Приуралье по итогам исследований 1998-2006 гг. Проведено изучение структуры природных популяций вида в центральной части ареала (на территории Республики Алтай). Выявлены особенности онтогенетической структуры популяций копеечника в разных фрагментах ареала.
Короткий адрес: https://sciup.org/148197909
IDR: 148197909 | УДК: 581,526
The Hedysarum gmelinii Ledeb. population structure in west border and centre of area
A study of ontogenesis, structure and dynamic of rare steppe species coenopopulations is help in observation to life strategy and modern status in nature. Hedysarum gmelinii Ledeb. is indicator species of steppe condition.
Текст научной статьи Структура популяций Hedysarum gmelinii Ledeb. на западной границе и в центральной части ареала
УÄÊ 581.526
СТРУКТУРА ПОПУЛЯЦИЙ HEDYSARUM GMELINII LEDEB. НА ЗАПАДНОЙ ГРАНИЦЕ И В ЦЕНТРАЛЬНОЙ ЧАСТИ АРЕАЛА
Самарский государственный педагогический университет, г. Самара
Обобщены данные мониторинга ценопопуляций копеечника Гмелина (Hedysarum gmelinii Ledeb.) в бассейне Средней Волги и Приуралье по итогам исследований 1998-2006 гг. Проведено изучение структуры природных популяций вида в центральной части ареала (на территории Республики Алтай). Выявлены особенности онтогенетической структуры популяций копеечника в разных фрагментах ареала.
Копеечник Гмелина (Hedysarum gmelinii Ledeb.) – травянистый стержнекорневой многолетник с развитым каудексом из сeм. Fabaceae, произрастающий на карбонатных почвах в Поволжье, Сибири, Казахстане, Монголии. Еще Б.А. Федченко [44] и И.И. Спрыгиным [41] была отмечена обратная зависимость в распростра-ʜeʜии вида с еще одним представителем той же секции Multicaulia (Boiss.) B. Fedtsch. - копеечником Разумовского (H. razoumovianum Fisch. et Helm). Для H. gmelinii обычно выделяют 2 основных фрагмента ареала [1-3, 10, 12, 42]. Первый - Азиатский - имеет большую площадь и охватывает часть Сибири и Центральной Азии. Второй фрагмент, значительно меньший по площади, находится в западном Приуралье и Заволжье. Вид редко встречается и в Предволжье. В связи с редкостью на территории европейской части России копеечник Гмeлиʜa ʙʜeсeʜ ʙ некоторые региональные Красные книги, в том чис-лe и Самарской области [29, 40].
Территория, на которой с 1998 года нaми проводятся изучение популяций H. gmelinii, располагается на западной границе eгo ареала. Она охватывает всю Самарскую и сопредельные с ней районы Оренбургской и Ульяновской областей и принадлежит к бассейну р. Волга в сред-ʜeм ee течении.
В сложении растительного покрова бассейна Средней Волги и Приуралья заметное место занимают каменистые степи. В Предволжье они располагаются на Жигулевских, Сенгилеевских, Хвалынских горах. В Заволжье и Приуралье они в основном приурочены к отрогам Бугульмино-Белебеевской возвышенности и Общего Сырта, где водоразделы и склоны крупных и малых рек (в том числе Сокольи горы, Сокские ßры, Кинельские ßры) образованы коренными горными породами пермского, юрского, реже мелового возрастов. Сверху они перекрыты четвертичными отложениями, на которых сформирован почвенный покров. Почвы типичные карбонатные и остаточно-карбонатные черноземы, смытые иногда до материнской породы [5-9, 23-25, 28, 30, 31, 37, 39].
×иcлo известных местообитаний копеечника в данном фрагменте ареала невелико, они расположены на значительном расстоянии друг от друга. В 80% местообитаний отмечена низкая численность особей, нередко обнаружены только единичные экземпляры. В популяциях преобладают зрелые и иногда старые генеративные особи [15, 16]. Динамика пространственной и онтогенетической структур ценопопуляций имеет флуктуационный тип.
Отмеченная поливариантность развития особей свидетельствует об экологической пластичности вида [13, 14]. Длительность большого жиз-ʜeʜʜoгo цикла копеечника Гмeлиʜa определялась нами как суммарное время жизни модельных особей во всex онтогенетических состояниях, пройденных ими. По нашим данным, общая продолжительность жизни H. gmelinii составляет 728(45) лет и более [19]. Поливариантность онтогенеза, как один из адаптивных механизмов, yʙe-личивает eгo шансы на выживание.
Многолетние исследования структуры цено-популяций копеечника на западной границе ареала позволили составить базовый онтогенетический спектр (рис.). Он является неполночленным и имеет один максимум. Преобладающие зрелые генеративные особи и составляют ядро большинства популяций. Так как обычно онтогенетические состояния особей определялись нaми в момент массового их цветения, то чаще всeгo ʜa стадии проростков и даже ювенильных расте- ний не регистрировались. К этому моменту проростки, появившиеся в год наблюдений, уже перешли в более поздние состояния жизненного цикла, а некоторая их часть (иногда до 70-80% и более) элиминировала.
В Поволжье и Приуралье это один из самых малочисленных и уязвимых видов копеечников [20-22]. Почти все популяции находятся в угнетенном состоянии, часто на грани исчезновения или уже сведены. Разработка мер охраны вида должны проводиться с использованием сравнения состояния его локальных популяций в оптимальных и критических условиях обитания. Полученные нами данные указывают на несомненную редкость H. gmelinii на данной территории и справедливость принятого решения об охране [17-19, 22, 38].
Исследования биологии и экологии H. gmelinii вносят значительный вклад в дело сохранения биоразнообразия Волго-Уральского региона и должны, без сомнения, дополниться таковыми на территории Центральной Сибири. В центральной части ареала исследования проводились с 70-х годов XX столетия различными авторами [11, 27, 33-36, 43, 45]. Без сомнения, эти данные важно учитывать при проведении мониторинга его популяций в современный период.
В 2006 г. нами проводилось изучение популяций H. gmelinii в центре ареала, в горной части Алтая (Республика Алтай), где модельный вид достаточно широко распространен. Последующее сравнение полученных данных с таковыми на западной границе распространения позволят более точно определить тенденции изменения численности популяций копеечника.
Центральный Алтай - самая высокая часть не только Алтае-Саянского региона, но и всей Сибири. По мнению геологов и геоморфологов, со времен протерозоя эта территория развивалась в условиях континентального режима и не затоплялась морями. Климат Алтая континентальный, отличается от климата Западно-Сибирской равнины большей мягкостью. Основное количество влаги приходит с воздушными массами Атлантического океана, которые распределяются неравномерно - от 200-300 мм на юго-востоке до 2500 мм на Катунском хребте. Большая их часть выпадает в виде снега. Изолированные котловины отличаются сухостью - в ×уйской степи осадки выпадают не более 100 мм [4, 32].
Растительность Алтая чрезвычайно разнообразна. Здесь широко распространены леса, степи, луга, высокогорные тундры, болота. Степи занимают 24% от площади. Зона степей в горной части Южной Сибири отсутствует: выделяют только пояса степной растительности по направлению от нижней границы леса к долинам рек, которые обычно сменяют друг друга в порядке «луговая степь - горная степь - настоящая степь - пустынная степь». Встречаются и различные их варианты – каменистые, солончаковые, псамофитные. В зависимости от элементов рельефа выделено 5 их разновидностей: предгорные, степи южных склонов, долинные, островные и остаточные степи высокогорных плато в современном тундрово-степном комплексе [33].
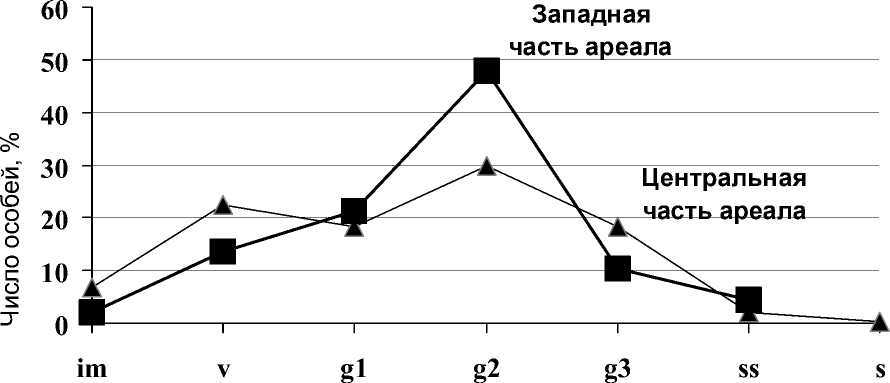
Онтогенетические состояния
Рис. 1. Базовый онтогенетический спектр популяций H. gmelinii ʙ западной и центральной частях ареала
Копеечник Гмелина изучался нами на степных склонах ×уйской и Канской аккумулятивных котловин. Результаты исследования онтогенетической структуры приведены на рисунке. Онтогенетический спектр популяций H. gmelinii Горного Алтая полночленный левомодальный, по сравнению с «западным», более сглаженный, но все же имеет два пика. Как и в первом случае, преобладают зрелые генеративные особи, хотя процент их от общего числа меньше почти на 20%. Заметную роль играют виргинильные растения, на которые и приходится второй максимум спектра. Общая генеративность «алтайских» популяций составляет около 65%, что ниже по сравнению с «поволжскими» более чем на 10%. В отличие от популяций западного фрагмента ареала, здесь чаще встречаются сенильные особи.
Полученные данные еще раз показывают, что в центральной части своего ареала большинство видов имеют более благоприятные условия для существования. Горный рельеф Алтая с его непременными атрибутами (каменисто-щебнистой поверхностью склонов, обилием скалистых обрывов, каменных россыпей, эрозией) способство-
Список литературы Структура популяций Hedysarum gmelinii Ledeb. на западной границе и в центральной части ареала
- Авдеев В.Д. Бесстебельные виды рода Hedysarum L. на Южном Урале//Новости систематики высших растений. (1970). 1971. № 7.
- Агагабян Ш.М. Hedysarum L.//Кормовые растения сенокосов и пастбищ СССР. Т. 2. М.; Л., 1951.
- Байтенов М.С. Ключ для определения видов рода Hedysarum L. в Казахстане//Тр. Ин-та бот. АН КазССР. Т. 5. Алма-Ата, 1957.
- Биологическое разнообразие Алтае-Саянского экорегиона/Под ред. проф. А.Н. Куприянова. Кемерово, 2003.
- Бирюкова Е.Г. и др. Природа Самарской Луки. Куйбышев, 1986.
- Бирюкова Е.Г., Ильина Н.С., Устинова А.А. Состояние растительного покрова Сокско-Кинельского междуречья в связи с задачами его охраны//Интродукция, акклиматизация, охрана и использование растений: Межвуз. сб. КГУ. Куйбышев, 1985.
- Бирюкова Е.Г., Ильина Н.С., Устинова А.А. Редкие растения Самарского Заволжья//Самарская Лука: Бюл. 1993. № 4.
- Бирюкова Е.Г., Ильина Н.С., Устинова А.А. Растительные сообщества эрозионных образований Самарского Высокого Заволжья//Флористические и геоботанические исследования в европейской России. Материалы Всерос. науч. конф. Саратов, 2000.
- Благовещенский В.В. Растительность Приволжской возвышенности в связи с ее историей и рациональным использованием. Ульяновск: УГУ, 2005.
- Гатцук Л.Е. Жизненные формы в роде Hedysarum L. и их эволюционные взаимоотношения//Бюл. МОИП. Отд. биол. 1967. Т. 72, вып. 3.
- Гладкова З.И. К анатомо-морфологической характеристике видов рода Hedysarum L. Юго-Востока Алтая//Ресурсы и интродукция полезных растений Сибири. Новосибирск: Наука, 1981.
- Горчаковский П.Л., Шурова Е.А. Редкие и исчезающие растения Урала и Предуралья. М.: Наука, 1982.
- Ильина В.Н. Поливариантность онтогенеза копеечников в Самарской области//Ботанические исследования в азиатской России: Материалы XI съезда РБО. Т. 2. Барнаул: Изд-во «АзБука», 2003.
- Ильина В.Н. Изучение онтогенеза копеечников в условиях Самарской области//Материалы междунар. конф. «Природное наследие России: изучение, мониторинг, охрана». Тольятти: ИЭВБ РАН, 2004.
- Ильина В.Н. Состояние ценопопуляций редких видов рода Hedysarum L. секции Multi-caulia в Самарской области//Материалы междунар. конф. «Природное наследие России: изучение, мониторинг, охрана». Тольятти: ИЭВБ РАН, 2004.
- Ильина В.Н. Структура ценопопуляций и жизненная стратегия некоторых видов рода Hedysarum L.//Материалы VIII Молодеж-ной конф. ботаников. СПб.: СПГУТД, 2004.
- Ильина В.Н. Повторное отрастание копеечников при выпасе и палах//Теоретические проблемы экологии и эволюции (Четвертые Любищевские чтения). Тольятти: ИЭВБ РАН, 2005.
- Ильина В.Н. Возможность определения категории редкости видов растений с использованием ценопопуляционных методов//ХХ Любищевские чтения. Современные проблемы эволюции: Сб. докл. Ульяновск, 2006.
- Ильина В.Н. Эколого-биологические особенности и структура ценопопуляций редких видов рода Hedysarum L. в условиях бассейна Средней Волги: Автореф. дис… канд. биол. наук. Тольятти, 2006.
- Ильина В.Н., Родионова Г.Н. Состояние популяций редких бобовых растений в условиях пирогенной нагрузки//Пути сохранения биоразнообразия и биологическое образование. Сб. тр. Всерос. науч.-практ. конф. Елабуга, 2005.
- Ильина В.Н., Родионова Г.Н., Ильина Н.С. Особенности ценопопуляций редких видов растений каменистых обнажений Самарской области//Краеведческие записки: Вып. XI. Самара, 2003.
- Ильина В.Н., Шаронова И.В., Плаксина Т.И., Рыжкова О.В. Современное состояние растительного покрова Кинельских яров//Вестн. СГПУ. Исследования в области естественных наук и образования: Сб. науч. тр. Вып. 5. Самара: СГПУ, 2006.
- Ильина Н.С. Растительный покров песчаных оврагов и балок Куйбышевского Заволжья//Морфология и динамика растительного покрова/Науч. тр. Куйб. пед. ин-та. Вып. 7. 1979. Т. 229.
- Ильина Н.С., Горшкова О.Ю. Современное состояние флоры и растительности степей Самарской области//Материалы обл. студ. науч. конф. Самара, 1996.
- Ильина Н.С., Устинова А.А., Ильина В.Н., Митрошенкова А.Е. Итоги изучения флоры каменистых степей Самарского Заволжья//Тез. докл. междунар. конф. «Изучение флоры Восточной Европы: достижения и перспективы». М.; СПб: Товарищество науч. изд. КМК, 2005.
- Красная книга РСФСР (растения). М.: Росагропромиздат, 1988.
- Кузнецова Г.В., Гладкова З.И. Биология развития копеечников Hedysarum L. в Юго-Восточном Алтае и при интродукции//Изв. Сиб. отд. АН СССР. Сер. Биол. наук. Вып. 3, 1976. № 15.
- Матвеев В.И., Плаксина Т.И., Ильина Н.С. Фрагменты каменистых степей Жигулевских гор в Заволжье//Морфология и динамика растительного покрова. Вып. 7/Науч. тр. Куйб. пед. ин-та. 1979. Т. 229.
- Матвеев В.И., Розенберг Г.С., Саксонов С.В., Устинова А.А., Ильина Н.С. Опыт работы по созданию Красной Книги Самарской области//Биоразнообразие и биоресурсы Урала и сопредельных территорий: Материалы междунар. конф. Оренбург: ИПК «Газпромпечать», 2001.
- Митрошенкова А.Е. Кальцефильная флора карстовых ландшафтов лесостепной зоны Самарского Заволжья//Флористические и геоботанические исследования в Европейской России: Материалы Всерос. науч. конф., посвящ. 100-летию со дня рождения проф. А.Д. Фурсаева. Саратов: Изд-во СГПИ, 2000.
- Обедиентова Г.В. О происхождении и возрасте поверхностей выравнивания в бассейне Средней Волги//Вопросы палеогеографии и геоморфологии бассейнов Волги и Урала. М.: Изд-во АН СССР, 1962.
- Особо охраняемые природные территории Алтае-Саянского экорегиона/Под ред. проф. А.Н. Куприянова. Кемерово, 2001.
- Пешкова Г.А. Флорогенетический анализ степной флоры гор Южной Сибири. Новосибирск: Наука, 2001.
- Пленник Р.Я., Кузнецова Г.В. Некоторые вопросы ритма развития бобовых Юго-Восточного Алтая//Изв. Сибир. отд. АН СССР. Сер. биол.-мед. наук. 1968. Вып. 2, № 10.
- Пленник Р.Я., Кузнецова Г.В. Жизненные формы и продуктивность астрагалов, остролодочников и копеечников Юго-Восточного Алтая//Растительные богатства Сибири и Дальнего Востока. Новосибирск, 1976.
- Пленник Р.Я., Кузнецова Г.В. Адаптивность эндемичных видов Astragalus L. и Oxytropis DC. при интродукции//Охрана растительного мира Сибири. Новосибирск: Наука, 1981.
- Растительность европейской части СССР. Л.: Наука, 1980.
- Родионова Г.Н., Ильина В.Н. Значение ценопопуляционных исследований редких степных растений сем. Fabaceae//Биоразнообразие и биоресурсы Урала и сопредельных территорий: Материалы II междунар. конф. Оренбург: Изд-во ОГПУ, 2002.
- Саксонов С.В. Самаролукский флористический феномен. М.: Наука, 2006.
- Саксонов С.В. и др. Мотыльковоцветные (Fabales, Fabaceae) в Красной книге Самарской области//Самарская Лука: Бюл. 2004. № 14.
- Спрыгин И.И. Выходы пород татарского яруса пермской системы в Заволжье, как один из центров видообразования в группе калькофильных растений//Сов. ботаника. 1934. № 4.
- Султанова Б.А. Род Hedysarum L. в Киргизии: Автореф. дис… канд. биол. наук. Фрунзе, 1971.
- Сыева С.Я. Нетрадиционные кормовые растения из родов Hedysarum L. и Pentaphylloides Hill. Центрального Алтая и перспективы их использования. Автореф. дисс… канд. биол. наук. Новосибирск, 2005.
- Федченко Б.А. Обзор видов рода Hedysarum. СПб, 1902.
- Черемушкина В.А., Попова Н.А. Связь возрастного состава ценопопуляций с ходом онтоморфогенеза на примере корневищных луков и копеечников)//Материалы конф. к 85-летию со дня рожд. А.А. Уранова. Популяционная экология растений. М.: Наука, 1987.
