Структура речных сообществ на перекатах, населяемых русской быстрянкой (Alburnoides rossicus Berg) в восточной части ареала
 в восточной части ареала")
Автор: Котельникова Валентина Сергеевна, Поздеев Иван Викторович, Огородов Сергей Павлович, Мартыненко Никита Александрович, Крайнев Евгений Юрьевич, Безматерных Валентин Владимирович
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Зоология
Статья в выпуске: 4, 2019 года.
Бесплатный доступ
Впервые приведена комплексная характеристика речных сообществ перекатов, населяемых русской быстрянкой, на примере разнотипных рек в восточной части её ареала. Выявлены ключевые виды водорослей, высшей водной растительности, зоопланктона, зообентоса, сиртона и рыб, составляющие специфичный «облик» сообщества. Высшая водная растительность, планкто- и ихтифауна отличались низким видовым богатством, альгофлора, бентофауна и сиртон были очень богаты в видовом отношении. Исследованный биотоп сильно зарастал к концу лета, формируя растительное сообщество, образованное урутью колосистой, рдестом пронзённолистным и нитчатыми водорослями (улотрикс или кладофора). Максимальные качественные и количественные показатели развития донных животных отмечены в июле-августе. В ихтиоценозах большинства водотоков русская быстрянка преобладала по численности и/или биомассе.
Эпилитон, зоопланктон, зообентос, сиртон, ихтиофауна, реки, бассейн камы, русская быстрянка
Короткий адрес: https://sciup.org/147227110
IDR: 147227110 | УДК: 574.58 | DOI: 10.17072/1994-9952-2019-4-441-451
Riverine communities structure in riffles, which inhabited Russian sprilin (Alburnoides rossicus Berg) in the eastern part of its area
The comprehensive description of the riverine communities of riffles which inhabited by the russian sprilin by the example of different types of watercources in the eastern part of its range have been given for the first time. The key species of algae, higher aquatic plants, zooplankton, zoobenthos, invertebrate drift and fishes, which make up the specific “appearance” of the community, have been identified. Higher aquatic vegetation, plankto- and ichthyofauna were characterized by low species richness, algoflora, benthic fauna and syrton were very rich in species. By the end of summer, the studied biotope was heavily overgrowning with the plant association of meakin, clasping-leaved pondweed and filamentous algae (Ulotrix or Cladophora). The maximum species richness and abundance of benthic invertebrates were observed in July-August. The russian sprilin was dominated by the number and/or biomass in the fish community of most rivers.
Текст научной статьи Структура речных сообществ на перекатах, населяемых русской быстрянкой (Alburnoides rossicus Berg) в восточной части ареала
Речные системы относятся к числу наиболее динамичных и характеризуются активным взаимодействием организмов и среды их обитания [Богатов, 1994].
По эколого-гидрологическим характеристикам все текучие воды принято делить на ритраль и по-тамаль [Illies, Botosaneanu, 1963, Богатов, 1994]. К ритрали чаще всего относят примыкающую к роднику часть водотока с каменистым или гравийногалечным грунтом, высокой скоростью течения, водой, насыщенной кислородом, и амплитудой среднемесячных температур до 20°. К потамали
относят примыкающую к ритрали нижнюю часть водотока с песчаным, заиленным или илистым грунтом, сравнительно небольшой скоростью течения, амплитудой среднемесячных температур выше 20° и с частыми проявлениями дефицита кислорода.
По отношению к продольному распределению основных групп речных организмов, всё население условно делится на ритрон и потамон, что соответствует делению водотоков на зоны ритрали и по-тамали. Ритрон обычно представлен фито- и зо-обентосными организмами, а также сообществом рыб с преобладанием лососевых. Фитобентос в зоне ритрали состоит из водорослевых обрастаний камней, мхов и немногих видов высших водных растений, а зообентос – преимущественно из организмов эпифауны, среди которых важную роль играют личинки амфибиотических насекомых отрядов подёнок, веснянок, ручейников и двукрылых. Из последних наибольшего количественно развития могут достигать личинки хирономид. Из организмов инфауны к макробентосу ритрали, в первую очередь, относятся олигохеты.
Население потамали состоит обычно из планктонных и бентосных организмов, а также из сообщества, главным образом, карповых рыб. В составе растительного компонента в потамали высокое развитие получают планктонные водоросли. Из планктонных животных – коловратки, веслоногие и ветвистоусые раки. Среди бентосных животных потамали ведущее значение могут иметь крупные двустворчатые и брюхоногие моллюски. Из других бентосных животных потамаль населяют те же группы беспозвоночных, что и ритраль, но заметного развития могут достигать личинки стрекоз и некоторые мелкие двустворчатые моллюски семейства Pisidiidae, а также олигохеты и личинки «красных» хирономид [Hynes, 1970; Богатов, 1994]
Цель работы – определение видового богатства и количественного соотношения видов тех биоценозов рек, одним из ключевых компонентов которых выступает русская быстрянка Alburnoides ros-sicus (Berg).
Материалы и методы исследования
На предмет наличия русской быстрянки Al-burnoides rossicus обследованы малые, средние и крупные реки бассейна Верхней и Нижней Волги.
Материал для данной работы собран на следующих водотоках: Ласьва, Буй (на территории Пермского края), Чепца, Вала (Удмуртская Республика), Юрюзань (Республика Башкортостан), Илеть (Республика Марий Эл), Мёша (Республика Татарстан) (рисунок).
Река Буй – приток Волги II порядка. Длина реки составляет 228 км, общая площадь водосбора – 6 530 км², средняя высота водосбора – 153 м.
Средний уклон – 0.4 м/км. Исследования на р. Буй проводились в 2010 и в 2013 гг. на одном и том же участке (N56.407903º E055.465690º). Данный перекат расположен на 171-м км от устья в Куедин-ском р-не Пермского края. В районе исследований ширина русла р. Буй в бровках пойменных берегов составляет 5–13 м. Дно на перекатах – песчаногравийное или гравийно-галечное. Скорости течения р. Буй изменяются от 0.1 м/с в межень до 1.5 м/с в паводки. Во время отбора проб на перекате скорости течения составляли около 1 м/c.
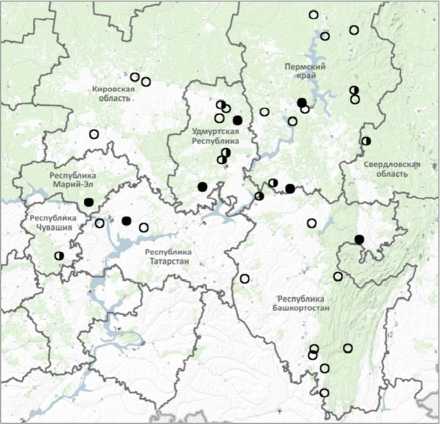
Карта-схема района проведения работ ○ – русская быстрянка не обнаружена; о– наличие русской быстрянки; • – места отбора материала
Река Чепца – приток Волги III порядка длиной 501 км, площадь её бассейна – 20 400 км², среднее падение реки 0.1 м/км. Район исследования расположен на территории Дебёсского района Удмуртской Республики на 419-м км от устья – в верхнем течении реки (N57.726988º E053.622222º). Ширина исследованного переката реки 22–27 м, скорость течения составляла 0.5–0.7 м/с на стрежнях переката при средней глубине 0.5 м. Грунт песчано-гравийно-галечный с преобладанием песка.
Река Юрюзань – приток Волги IV порядка, длина реки составляет 404 км, площадь водосборного бассейна – 7 240 км², Средний уклон реки значительно различается на отдельных участках (0.4–1.2м/км), составляя в районе проведения работ: 0.6–0.8 м/км. Исследованный участок
(N55.355497º E057.989065º) расположен в среднем течении р. Юрюзани на 144-м км от устья в Сала-ватском р-не Республики Башкортостан и представляет собой перекат с песчано-гравийногалечным дном и скоростью течения около 0.2 м/с. Средняя глубина составляла 0.3 м.
Река Вала – приток Волги IV порядка. Длина реки – 196 км, площадь бассейна – 7 360 км². Исследованный участок (N56.453008º E052.369820º) расположен в верхнем течении реки на 152-м км от устья в Мо-жгинском р-не Удмуртской Республики. Скорость течения на стрежне переката достигала 0.25 м/с, грунт – гравийно-галечный. Русло реки глубоко врезано, берега обрывистые. Средняя глубина составляла 0.6 м, максимальная – 1.5 м.
Река Ласьва – приток Волги II порядка протяженностью 78 км, площадь водосбора 418 км². Исследованный участок (N58.045240º E055.932278º) расположен в среднем течении р. Ласьвы на 33м км от устья в Краснокамском р-не Пермского края и представляет собой перекат с песчано-гравийно-галечным дном и скоростью течения около 0.7 м/с. Средняя глубина составляла 0.5 м.
Река Илеть – левый приток Волги I порядка. Длина реки – 204 км, площадь водосбора – 6 471 км². Исследованный участок (N56.135791º E048.333173º) расположен в нижнем течении р. Илеть на 49-м км от устья, в Звениговском р-не Республики Марий Эл. Ширина русла – 70–100 м, в межень оно разделено большим количеством небольших песчаных островов, перевалено топляками. Брёвна и ветки, погружённые в воду, обильно покрыты прикреплёнными нитчатыми водорослями, тогда как б о льшая площадь дна свободна от обрастаний. Грунт песчаный. Скорость течения – 0.1 м/с в прибрежье и 0.4 м/с на стрежне. Преобладающая глубина – 0.3–0.5 м, максимальная – 1.0 м.
Река Мёша – приток Волги II порядка длиной 204 км и площадью водосбора 4 180 км². Средний уклон водотока составляет 0.4 м/км. Исследованный участок водотока (N55.750765 E49.671744º) расположен на 60-м км от устья в Пестречинском р-не Республики Татарстан. Данный перекат представляет собой протяжённый прямой участок реки длиной 300 м. Грунт – песчано-гравийногалечный, местами лежат отдельные валуны. Ширина русла – 30–40 м. Скорость течения составляли 0.2–0.5 м/с, глубина не превышала 0.75 м, средняя глубина – 0.5 м.
Во всех водотоках исследованные перекаты в течение вегетационного периода обильно зарастали зелёными нитчатыми водорослями и высшей водной растительностью.
По гидрологическим и гидробиологическим параметрам исследованные нами участки можно разделить на три типа:
o ритраль : р. Буй. Высокие скорости течения (до 1.0 м/с), значительная затенённость русла с шириной 5–13 м и глубинами 0.3–0.5 м. Большое количество древесной растительности на обрывистых берегах;
o потамаль малых и средних водотоков: р. Вала, Ласьва, Мёша и Илеть. Глубоко врезанное русло (10–25 м) с ярко выраженным стрежнем и преобладающими глубинами 0.5–0.7 м. Скорости течения от 0.2–0.5 м/с;
o потамаль крупных водотоков : р. Чепца и Юрюзань. Довольно широкие участки (20–70 м) без ярко выраженного стрежня. Преобладающие глубины – 0.3–0.5 м. Скорости течения 0.2–0.7 м/с.
Материалом для работы послужили пробы фитопланктона и фитоперифитона (6), зоопланктона (76), зообентоса (78), сиртона (53) и ихтиофауны (15), отобранные в период 2010–2017 гг. в ходе экспедиционных и стационарных исследований.
Пробы фитопланктона (водотоки с песчаным грунтом) объёмом 1 л отбирали с поверхностного слоя воды, фитоперифитона (водотоки с каменистым грунтом) – путём смыва с камней с последующим измерением их площади проективного покрытия. Сгущение проведено фильтрационным методом. Количественное изучение проводилось в камере «Учинской» объёмом 0.02 см³. Биомасса определялась счётно-объёмным методом, при котором 109 мкм³ соответствует 1 мг сырой биомассы. Стереометрическим методом определялись средние размеры для каждого вида; плотность водорослей считали равной 1 г/м³. [Методические рекомендации …, 1984; Водоросли, 1989].
При отборе проб зоопланктона воду в объеме 50 л процеживали через сеть Джеди с ячеёй 0.1 мм, фиксировали 4%-ным формалином [Методика изучения …, 1975].
Сбор проб макрозообентоса осуществляли при помощи бентометра 625 см², грунт промывали через газ с размером отверстий 0.21 мм. Пробы дрифта отобраны двумя сачками-ловушками с тем же размером отверстий газовой ткани, имеющими входное отверстие 1000 см² и глубину мешка 75 см, которые одновременно устанавливали на 5 мин.; таким образом, общая экспозиция каждой пробы составляла 10 мин. Все организмы выбирали из проб живыми и фиксировали 8%-ным формалином [Методика изучения …, 1975; Богатов, 2005]. Донные животные определялись до вида, в случае невозможности точной идентификации – до «низшего определяемого таксона» (НОТ).
Для сбора ихтиофауны использовали два мальковых невода. Первый – длиной 5 м, высотой 1.5 м, диаметром ячеи 4 мм использовался на участках рек, ширина которых не превышала 5– 7 м. Второй – длиной 10 м, высотой 1.8 м, диаметром ячеи 3 мм применяли на участках рек с большей шириной.
Для выделения доминантных комплексов сообществ и выявления значимости отдельных видов использовали модифицированный индекс плотности Арнольди [Щербина, 1993], к числу доминан-тов относили виды, величина индекса для которых составила 15% и более, к субдоминантам – 10– 15%.
Структуру сообществ анализировали при помощи числа видов ( S ), коэффициента видового сходства Чекановского-Серенсена ( I CzS ), индекса видового разнообразия Шеннона по численности ( H N , бит/экз.) и биомассе ( H B , бит/г) и индекса разности выравненностей ( D E’ ). Также использовали показатель доминирования Симпсона, рассчитанный как по численности, так и по биомассе ( C N , C B ) и показатель видового разнообразия ( d ) Мар-галефа [Одум, 1975; Песенко, 1982; Алимов, 2001; Денисенко и др., 2013]. Расчёт индексов и их стандартных ошибок произведён в среде r версии 3.4.3 [R Core Team, 2018].
Анализ распространения видов рыб, состав их пищи и характер питания (кроме русской быстрян-ки) оценивался по обобщающим литературным источникам [Никольский, 1971, 1974; Kottelat, Frey-hof, 2007].
Результаты
Альгофлора
Видовой состав водорослей и цианопрокариот биотопа, занимаемого быстрянкой, в изученных водотоках весьма разнообразен. Так, в отдельных водотоках зарегистрировано от 51 до 103 таксонов рангов ниже рода из 52 родов, 32 семейств, 19 порядков, 9 классов и 7 отделов.
Ведущим отделом по видовому богатству выступал Bacillariophyta, формируя 62–88% видового списка альгофлоры. Среди диатомовых водорослей по количеству видов лидировали семейства Naviculaceae (20 внутриродовых таксонов), Cymbellaceae (12) и Fragilariaceae (11). Наиболее богатым в видовом отношении оказался род Navicula (18 внутриродовых таксонов), относительно велико разнообразие родов Placoneis и Cymbella (по 5), Fragilaria , Nitzschia , Gomphonema (по 4 внутриродовых таксона в каждом).
Второе место по числу внутривидовых таксонов, обеспечивая от 8 до 26% этого показателя, занимал отдел Chlorophyta. Ключевыми семействами среди зелёных водорослей следует считать Scenedesmaceae (8), Selenestraceae (5) и Hydrodictyaceae (5). Ведущими родами становились Monoraphidium и Desmodesmus (5 и 4 внутри-родовых таксона, соответственно).
На третьем месте по видовому богатству выступал отдел Streptophyta (3–10%), основу которого слагали представители семейства Desmidiaceae (7) c родом Cosmarium (6 внутриродовых таксонов).
Незначительно уступает отдел Cyanoprocaryota, формировавший в разных водотоках от 1 до 7% видового разнообразия альгофлоры. В нём наибольшим числом видов было представлено семейство Merismopediaceae (3) c родом Merismopedia (3).
Отделы Euglenophyta, Chrysophyta и Xan-thophyta суммарно обеспечивали менее 9% от общего числа видов.
Численность эпилитона на разных участках дна биотопа, населяемого русской быстрянкой в середине лета (июль), варьировала от 321 до 1990 млн. кл./м², биомасса – от 1.20 до 7.82 г/м².
По численности преобладали мелкоклеточные цианопрокариоты Pseudanabaena galeata Böcher, Merismopedia punctata Meyen, зелёная водоросль Dictyosphaerium pulchellum H.C. Wood, одношовная диатомея Cocconeis pediculus Ehrenb., навикулоидные диатомовые водоросли Navicula tripunctata (O. Müll.) и Navicula lanceolata Ehrenb. и бесшовная диатомея Diatoma vulgare Bory. По биомассе доминировали крупноклеточные диатомеи, относящиеся к разным порядкам: амфороидная Amphora ovalis (Kütz.) Kütz, гомфоцимбеллоидная Cymbella neolanceolata W. Silva, навикулоидная Gyrosigma attenuatum (Kütz.) Rabenh., одношовная Cocconeis pediculus , бесшовные Diatoma vulgare и Ulnaria ulna (Nitzsch) Compère, навикулоидные Navicula tripunctata , Navicula lanceolata и десмидиевая водоросль Cosmarium botrytis Menegh.
В конце лета (август) в наиболее крупных из изученных водотоков развитие получали зелёные нитчатые водоросли родов Ulotrix и Cladophora , создавая обильные куртины, превышающие по площади зарастания высшую водную растительность. Уже к концу августа их талломы сильно заиливались, а в середине сентября – начинали отмирать и, отрываясь, сносились течением вместе с населяющими их гидробионтами.
Высшая водная растительность
На исследованных биотопах водотоков произрастали отдельные куртины рдестов и урути колосистой – Potamogeton alpinus Balb., Potamogeton acutifolius Link, Potamogeton perfoliatus L. и Myriophyllum spicatum L. Занимая в июне-июле от 10 до 50% площади дна, к августу покрывали не менее четверти акватории биотопа, а в крупных реках – более половины.
В большинстве рек плёсовые участки выше и ниже исследованных перекатов обычно зарастали рдестами Potamogeton perfoliatus , Potamogeton gramineus L., Potamogeton lucens L. и Potamogeton praelongus Wulf. На наиболее спокойных участках дна плёсов произрастали отдельные побеги Po-tamogeton natans L., Nuphar lutea (L.) Smith, Elodea canadensis Michx. и Batrachium trichophyl-lum (Chaix) Van den Bosch. Площадь зарослей достигала 90%, оставляя свободной только узкую полосу стрежня водотока.
Зоопланктон
Планктофауна биотопа, занимаемого быстрян-кой, в изученных водотоках достаточно однообразна и включает 13 видов коловраток и 6 видов ветвистоусых раков. Веслоногие раки были представлены только науплиальными и копеподитными стадиями развития, их видовая принадлежность не определена. Наибольшую частоту встречаемости проявили виды коловраток Cephalodella gibba (Ehrenberg), Testudinella patina (Hermann) и кладоцер Alona costata Sars, Bosmina longirostris (O.F. Muller), Chydorus sphaericus (O.F. Muller). Видовое богатство планктонных беспозвоночных изученного биотопа в отдельных водотоках варьировало от 5 до 12 таксонов.
Биомасса зоопланктона изученных перекатов рек изменялась в среднем от 0.05 до 0.75 мг/м³, численность – от 1.06 до 778 тыс.экз./м³. В целом, количественные показатели развития планктонных сообществ увеличивались с протяжённостью водотока. Основу численности планктонных животных создавали коловратки (55–87%), основу биомассы – коловратки или кладоцеры (38–53%).
Видами, обеспечивающими наибольшие показатели численности и биомассы планктонных животных выступали коловратки Cephalodella gibba , Brachionus quadridentatus Hermann , Euchlanis dila-tata Ehrenberg, Euchlanis oropha Gosse и Testudi-nella patina , реже – кладоцеры Bosmina longirostris и Chydorus sphaericus .
Зообентос
В бентофауне исследованных участков рек зарегистрированы 171 вид и НОТ, принадлежащие 5 классам: малощетинковые черви (8 видов), пиявки (5), двустворчатые моллюски (14), брюхоногие моллюски (12) и насекомые (132 вида и НОТ). Среди последних наибольшим видовым богатством отличались хирономиды (59 видов и НОТ), подёнки (18 видов) и ручейники (17), остальные группы насекомых – стрекозы, веснянки, клопы, жуки, вислокрылки, бабочки, комары-долгоножки, болотницы, мошки, мокрецы, слепни, атерициды, толкунчики и настоящие мухи – насчитывали меньшее количество видов и НОТ.
Видовое богатство донных животных исследованного биотопа в разных реках варьировало от 10 видов на чистых песках до 90–95 – на грунтах с высоким разнообразием гранулометрических фракций. Численность и биомасса зообентоса также характеризовались минимальными величинами на чистых песчаных грунтах – в среднем 8.5 тыс.экз./м² и 8.49 г/м², соответственно. На песчано-гравийно-галечных грунтах обилие донных животных возрастало в среднем до 22.34 г/м² и 6.0 тыс.экз./м². Грунты с крупной галькой и валунами, а также с наличием перифитоных водорос- левых матов характеризовались максимальными показателями численности и биомассы зообентон-тов – в среднем 99.06 г/м² и 8.5 тыс.экз./м².
К постоянным видам, частота встречаемости которых на биотопе, занимаемом быстрянкой, превысила 50%, относятся шесть: ручейники Psy-chomyia pusilla (Fabricius) и Hydropsyche contube-rnalis McLachlan, клоп Aphelocheirus aestivalis (Fabricius), подёнка Caenis macrura Stephens, хи-рономиды Thienemannimyia fusciceps (Edwards) и Polypedilum scalaenum (Schrank).
Высокую частоту встречаемости (25–50%) проявляли 12 видов: болотница Hexatoma bicolor Meigen, хирономиды Cladotanytarsus mancus (Walker), Microtendipes chloris (Meigen), Potthastia gaedii (Meigen), Orthocladius rhyacobius Kieffer, подёнки Ephemera lineata Eaton, Potamanthus lute-us (Linnaeus), Baetis vernus Curtis, двустворчатые моллюски Neopisidium tenuilineatum (Stelfox), Eu-glesa casertana (Poli) и олигохета Lumbriculus var-iegatus (Muller).
Для ряда видов встречаемость более 50% отмечена лишь в отдельных водотоках, это олигохеты Tubifex newaensis (Michaelsen) и Tubifex tubifex (Muller), веснянка Isogenus nubecula Newman ручейник Hydroptila tineoides Dalman, жук Elmis maugetii Latreille, хирономиды Thienemannimyia carnea (Fabricius), Paracladopelma camptolabis (Kieffer) и Stictochironomus sticticus (Fabricius).
Структура донных сообществ на перекатах изученных рек, оцененная при помощи индексов Шеннона и Симпсона, характеризуется высокой сложностью (табл. 1).
Минимальные величины индексов отмечены в р. Илети, где перекат, населяемый быстрянкой, отличается чистыми песчаными грунтами, и, соответственно, низким видовым богатством. Индексы видового разнообразия Шеннона и Симпсона принимали в среднем наименьшие величины, а их стандартные ошибки были максимальными.
Донные сообщества перекатов всех прочих исследованных водотоков более разнообразны в видовом отношении. В условиях ритрали (р. Буй) отмечена наибольшая амплитуда варьирования рассчитанных параметров в ходе сезонной динамики. Так, в июне величины индексов по биомассе (HB, CB) в среднем превышали таковые по численности (HN, СN). Это связано с массовым появлением молоди хирономиды Orthocladius rubicundus (Meigen) после весеннего вылета. В августе, при максимальном видовом богатстве, величина индекса Шеннона, рассчитанная по численности, уже более чем в 2 раза превосходила таковую по биомассе. В это время в донных сообществах увеличивается количество других групп зообентонтов – подёнок, ручейников, клопов и др. В октябре, при снижении видового богатства бентосных беспозво- ночных, величины индексов, рассчитанных по лее чем в 2 раза в течение летне-осеннего сезона, численности и биомассе, выравниваются относи- величины индексов Шеннона и Симпсона остают-тельно друг друга и остаются на высоком уровне. ся высоки. Разнообразие донных сообществ по
В потамали (на примере р. Чепцы), несмотря на численности в течение всего летне-осеннего периварьирование видового богатства бентофауны бо- ода превышает таковое по биомассе (табл. 1).
Таблица 1
Видовое богатство и разнообразие донных сообществ перекатов изученных рек
|
Водоток |
Месяц |
S |
H N |
H B |
D E’ |
C N |
C B |
|
Чепца |
VI |
42 |
4.16±0.01 |
3.21±0.02 |
–0.18 |
0.92±0.03 |
0.79±0.05 |
|
VII |
48 |
4.65±0.01 |
3.47±0.03 |
–0.21 |
0.94±0.03 |
0.85±0.03 |
|
|
VIII |
24 |
3.22±0.02 |
2.47±0.05 |
–0.16 |
0.81±0.03 |
0.67±0.05 |
|
|
IX |
34 |
3.54±0.03 |
3.40±0.03 |
–0.03 |
0.85±0.03 |
0.86±0.03 |
|
|
Буй |
VI |
30 |
1.99±0.03 |
3.38±0.02 |
0.28 |
0.54±0.13 |
0.87±0.05 |
|
VIII |
86 |
4.58±0.01 |
2.15±1.26 |
–0.38 |
0.94±0.003 |
0.59±0.09 |
|
|
X |
62 |
4.49±0.01 |
3.72±0.02 |
–0.13 |
0.94±0.001 |
0.86±0.02 |
|
|
Вала |
VIII |
46 |
4.23±0.05 |
3.05±0.06 |
–0.21 |
0.89±0.05 |
0.76±0.05 |
|
Илеть |
VIII |
10 |
2.64±0.20 |
1.95±0.35 |
–0.21 |
0.80±0.09 |
0.60±0.12 |
|
Мёша |
VIII |
38 |
3.97±0.03 |
3.35±0.14 |
–0.12 |
0.90±0.03 |
0.84±0.10 |
|
Юрюзань |
VIII |
20 |
4.12±0.01 |
3.33±0.03 |
–0.18 |
0.92±0.01 |
0.79±0.03 |
Дрифт донных беспозвоночных
Видовое богатство дрифтующих донных беспозвоночных на перекате в потамали, занимаемом быстрянкой (на примере р. Чепцы), составило 109 видов и НОТ, из них в июле зарегистрировано 56, в августе – 66, в сентябре – 50 [Поздеев, Котельникова, 2014]. Сравнение состава сиртона с таковым донных сообществ по результатам одновременно собранных проб показал, что только в бентосе были зарегистрированы веснянки, бабочки, атерициды и мокрецы, только в сиртоне – пиявки, большекрылые, сетчатокрылые и мошки. Видовой состав олигохет, двустворчатых моллюсков и стрекоз в бентосе и дрифте полностью разнился. Общее видовое сходство собственно бентофауны и сиртона, оцениваемое по величине индекса Чека-новского-Серенсена, оказалось равным 0.35 и снижалось от июля (0.31) к августу (0.27) и далее – к сентябрю (0.22). Видовое богатство сиртона в разные месяцы оставалось более постоянным ( I CzS = 0.47–0.49); в донных сообществах наблюдались изменения, выраженные в увеличении коэффициента видового сходства между соседними временными выделами (июль-август – 0.52, август-сентябрь – 0.61) и снижении сходства между крайними (июль-сентябрь – 0.37).
Средние численность и биомасса донных животных, сносимых за сутки через сечение потока шириной 1 м и высотой, равной глубине участка в исследуемый период времени, составили 19.0 тыс.экз./(м²·сутки) и 14.77 г/(м²·сутки). Наибольшую долю в биомассе дрифта донных животных в среднем за период исследования создавали подёнки (44.8%), хирономиды и ручейники, наибольшую численность сиртона обеспечивали хирономиды, подёнки, мошки и олигохеты. В сиртоне частота встречаемости только трёх видов из олигохет и хирономид – Stylaria lacustris (Linnaeus), Thiene-mannimyia fusciceps и Cricotopus bicinctus Meigen – превысила 50%, их можно отнести к постоянным компонентам дрифта зообентонтов.
В суточной динамике дрифта донных беспозвоночных отмечены следующие закономерности. В утренние и дневные часы (06:00–18:00) сиртон отсутствовал или отличался минимальными численностью и биомассой. В это время в составе сиртона преобладали хирономиды, меньшее значение имели подёнки, ручейники и олигохеты, остальные группы были представлены единично или отсутствовали. Большинство донных животных дрифто-вало в сумеречные и ночные часы. Начало массового выхода в дрифт наблюдалось примерно за час до захода солнца. Наибольшие численность и биомасса дрифтующих беспозвоночных регистрировались в период с 21:00 до 02:00 часов ночи с максимумом в полночь. Окончание массового выхода зообентонтов в поток происходило за 1–2 часа до восхода солнца. Все преобладающие в сиртоне виды донных животных в той или иной степени присутствовали в речном потоке как в ночные часы, так и днём.
Сезонная динамика количества дрифтующих донных беспозвоночных изменялась следующим образом. В июле были отмечены наименьшие показатели биомассы и численности дрифта – 7.70 г/(м²·сутки) и 16.6 тыс.экз./(м²·сутки), среди них доминировала подёнка Procloeon bifidum (Bengtsson), в качестве субдоминанта выступала хирономида Thienemannimyia fusciceps. В августе общая биомасса и численность дрифтующих беспозвоночных возросли, составив 13.44 г/ (м²·сутки) и 22.9 тыс.экз./(м²·сутки), соответственно. Полностью сменился состав доминантных комплексов. Его слагали виды мошек и подёнок – Simulium ornatum Meigen (доминант), Baetis buceratus Eaton и Baetis vernus (субдоминанты). В сентябре биомасса дрифтующих донных беспозвоночных возросла почти вдвое, составив 23.18 г/(м²·сутки) при численности около 17.4 тыс.экз./(м²·сутки). Доминантные комплексы снова полностью сменились: в качестве доминан-тов выступали виды подёнок и хирономид: Ephemera lineata, Cricotopus bicinctus, в качестве субдоминантов – подёнка Baetis digitatus Bengtsson и олигохета Stylaria lacustris.
Ихтиофауна
Ихтиофауна исследованных перекатов водотоков представлена 11-ю реофильными, реолимно-фильными и эврибионтными видами трёх ихтио-фаунистических комплексов: понто-каспийского (русская быстрянка, уклейка и голавль), бореально-предгорного (речной гольян, усатый голец и европейский хариус) и бореально-равнинного (обыкновенная плотва, обыкновенный пескарь, обыкновенная щука, речной окунь, обыкновенная щиповка).
Видовое богатство ихтиофауны исследованного биотопа в разных реках варьировало от 6 до 10 видов. Относительные численность и масса рыб различались на порядок: 2.0–50.7 тыс. экз./га и 18.75–
132.26 кг/га, соответственно, в целом снижаясь от участков в ритрали к участкам в потамали малых и средних водотоков, и далее – к участкам в потама-ли крупных рек.
По характеру питания во всех водотоках присутствовали представители 5 групп: хищники (щука), бентофаги (усатый голец, пескарь, гольян, щиповка), планктофаги (уклейка), полизоофаги (окунь, голавль, хариус), поли(эври)фаги (плотва и быстрянка). По составу пищи большая часть видов – преимущественно зоофаги, меньшая часть – плотва и быстрянка – способны питаться не только животной, но также растительной пищей и детритом.
На исследованном перекате р. Буй отловлена 1 731 особь 8 видов рыб, относящихся к 4 отрядам: Cypriniformes (6 видов), Esociformes (1 вид), Salm-oniformes (1 вид), Perciformes (1 вид). В структуре рыбного сообщества по численности и биомассе преобладал речной гольян Phoxinus phoxinus (Linnaeus) – 87 и 63%, соответственно. Доли остальных видов были значительно ниже, русская быстрянка занимала второе место по численности и биомассе рыб – 6 и 13%, соответственно (табл. 2).
Таблица 2
Видовой состав, численность ( N , %) и биомасса ( B , %) рыб в ихтиофауне исследованных рек
|
Вид |
Чепца |
Буй |
Юрюзань |
Вала |
||||
|
N |
B |
N |
B |
N |
B |
N |
B |
|
|
Alburnoides rossicus |
41.4 |
38.2 |
5.5 |
12.4 |
41.6 |
31.0 |
82.7 |
35.9 |
|
Alburnus alburnus |
14.7 |
14.0 |
2.3 |
5.3 |
19.3 |
5.0 |
4.9 |
2.2 |
|
Barbatula barbatula |
– |
– |
0.7 |
0.4 |
3.1 |
1.1 |
– |
– |
|
Cobitis taenia |
– |
– |
– |
– |
– |
– |
1.2 |
1.1 |
|
Esox lucius |
0.4 |
4.8 |
0.1 |
3.1 |
2.0 |
6.6 |
1.2 |
2.2 |
|
Gobio gobio |
5.3 |
9.7 |
3.4 |
11.4 |
1.7 |
1.7 |
– |
– |
|
Perca fluviatilis |
0.5 |
3.0 |
0.4 |
0.3 |
10.2 |
27.9 |
– |
– |
|
Phoxinus phoxinus |
16.0 |
7.1 |
87.2 |
63.4 |
1.3 |
0.2 |
– |
– |
|
Rutilus rutilus |
17.0 |
15.9 |
– |
– |
1.3 |
0.4 |
7.4 |
8.7 |
|
Squalius cephalus |
4.7 |
7.3 |
– |
– |
15.3 |
8.4 |
2.5 |
50.0 |
|
Thymallus thymallus |
– |
– |
0.4 |
3.5 |
4.0 |
17.7 |
– |
– |
|
Всего |
100.0 |
100.0 |
100.0 |
100.0 |
100.0 |
100.0 |
100.0 |
100.0 |
|
Индекс Маргалефа |
0.75 |
0.65 |
1.18 |
0.58 |
||||
На исследованном перекате р. Чепцы отловлено 656 особей 8 видов рыб, относящихся к 3 отрядам: Cypriniformes (6 видов), Esociformes (1 вид), Perciformes (1 вид). В структуре рыбного сообщества переката по численности и массе преобладала русская быстрянка Alburnoides rossicus – 41 и 38%, соответственно. Доли остальных видов были значительно ниже.
На исследованном перекате р. Юрюзани отловлено 1 136 особей 10 видов рыб, относящихся к 4 отрядам: Cypriniformes (7 видов), Esociformes (1 вид), Salmoniformes (1 вид), Perciformes (1 вид). В структуре рыбного сообщества по численности и массе преобладала русская быстрянка – 42 и 31%, соответственно. Также большую роль по массе играл речной окунь Perca fluviatilis Linnaeus (28%), Доли остальных видов были значительно ниже
(табл. 2).
На исследованном перекате р. Валы отловлено 29 особей 6 видов рыб, относящихся к 2 отрядам: Cypriniformes (5 видов), Esociformes (1 вид), представители бореально-предгорного ихтиофаунисти-ческого комплекса отсутствовали. В структуре рыбного сообщества по численности преобладала русская быстрянка Alburnoides rossicus (83%), по массе – голавль Squalius cephalus (Linnaeus) (50%), также большую роль по массе играла русская быстрянка (36%), доля остальных видов была значительно ниже.
Обсуждение
Альгофлора исследованных участков рек достаточно разнообразна, большинство таксонов приходилось на диатомовые водоросли. По массе в эпилитоне также преобладали представители Ba-cillariophyta и Chlorophyta. Виды, получавшие здесь наибольшее развитие по приуроченности к местообитанию относились собственно к бентосным, меньшая часть – к планктонным или планктонно-бентосным. Большинство таксонов ниже рода характеризуются космополитическим распространением, это эвритермные виды, индифферентные по отношению к показателям pH и солёности вод, β-мезосапробы. Отдельным и важным компонентом альгофлоры биотопа, занимаемого быстрянкой, выступали нитчатые зеленые водоросли Ulothrix, реже – Cladophora, формируя обильные заросли. Среди высшей водной растительности ключевую роль по обилию и распространению играли виды Potamogeton perfoliatus и Myriophyllum spicatum.
Настоящая планктофауна на перекатах, занимаемых русской быстрянкой, отсутствует и сформирована небольшим числом видов преимущественно литорального комплекса, а их численность и биомасса были крайне низкими. Основной группой выступали виды, тяготеющие к придонным и прибрежным биотопам, а также к зарослям высшей водной растительности на вышележащих плёсовых участках изученных рек. Наряду с обитателями мелководий, в зоопланктоне отмечено несколько видов собственно планктонных и эври-бионтных коловраток ( Kellicottia longispina (Kel-licott) , Brachionus quadridentatus, Keratella quadra-ta (Muller) , Keratella cochlearis (Gosse) и кладоцер ( Bosmina longirostris ), а также представители мейобентоса (виды рода Alona ). По отношению к сапробности на исследованных участках рек преобладали олигосапробы и виды, занимающие промежуточное положение между олиго- и β -мезосапробами.
В целом, состав планктофауны перекатов, занимаемых быстрянкой, слагается широко распространенными в водных объектах европейской умеренной зоны видами и обнаруживает высокую степень сходства с зоопланктоном самых малых водотоков Камского региона: формируется на вышележащих плёсах преимущественно зарослевыми и придонными видами, а также обогащается собственно планктонными обитателями плёсов и водоёмов пойменной системы.
Донные сообщества биотопа, занимаемого быстрянкой, учитывая все изученные водотоки, характеризуются высоким видовым богатством и таксономическим разнообразием. Анализ состава массовых видов исследованных перекатов показывает преобладание эпилитических форм или псаммофилов (в случае песчаных перекатов), эврифа-гов, детритофагов и селективных альгофагов. Большую долю таксономического списка слагают виды-детритофаги, приуроченные к пескам и/или ведущие роющий образ жизни. Преимущественно это β-мезосапробы, и, даже в случае массового развития видов с низкой индикаторной значимостью, высоко чувствительные к загрязнению виды обнаруживают высокую встречаемость.
Обращает на себя внимание низкая частота встречаемости в бентосе исследованных перекатов видов хирономид и мошек Cricotopus bicinctus (8.7%) и Simulium ornatum (4.3%). Именно эти виды двукрылых составляют основу питания молоди быстрянки [Котельникова, 2016, 2017], что говорит об их преимущественном потреблении рыбами из толщи воды как компонента сиртона.
Видовое богатство сиртона в течение всего периода исследования превосходило таковое зообентоса, поскольку, на наш взгляд, слагалось из двух источников – фитофильной фауны вышележащего плёса и донных сообществ собственно переката. Особенно хорошо это демонстрируют различия состава олигохет в донных сообществах ( Tubifex newaensis, Tubifex tubifex ) и сиртоне ( Stylaria la-custris , Haemonais waldvogeli Bretscher, Ophidonais serpentina (Muller), виды рода Nais ).
Преобладающие виды в составе сиртона постоянно сменялись, что связано с прохождением ими циклов развития. Однако в дрифте постоянно присутствовал комплекс видов, преимущественно хи-рономид, с растянутым вылетом, обеспечивающий стабильность структуры сиртона в летне-осенний период. Донные сообщества переката, напротив, отличались большей стабильностью состава руководящих видов, в основном это виды с продолжительным циклом развития, в то время как состав второстепенных видов менялся от месяца к месяцу.
Величины индекса D E’ , рассчитанные по средним величинам индекса Шеннона за отдельные месяцы для разных водотоков говорят об отсутствии стресса или незначительном стрессе (отрицательные значения). Положительная величина индекса D E’ , характеризующая донные сообщества р. Буй в июне, говорит о наличии стресса. Ввиду отсутствия крупных населённых пунктов вблизи исследованного участка реки и высоких величинах D E’ в последующие месяцы, мы интерпретируем этот показатель как отражение восстановления донных сообществ после весеннего половодья.
Основу численности сиртона на исследованном перекате составляли донные животные, населяющие вышележащий плёс, что подтверждают низкие величины коэффициента видового сходства с бентосом переката. Их переход в дрифт происходил главным образом с целью вылета (подёнки, хирономиды, мошки) и, в меньшей степени, с целью миграции на нижележащие участки (олигохеты, личинки амфибиотических насекомых младших возрастов). Соотношение этих групп менялось от июля к сентябрю – доля вылетающих форм снижалась, доля мигрантов возрастала. Меньшую часть сиртона на перекате слагали собственно реофильные формы, населяющие перекат (Hydro-psychidae, Ephemeridae, отдельные виды Chirono-midae, Aphelocheirus aestivalis). Их выход в дрифт, в большинстве случаев, был вызван вылетом (июль-август). В конце вегетационного периода со снижением интенсивности лёта амфибиотических насекомых, виды, населяющие перекат, оказывались в речном потоке вынужденно – вместе с кусками населяемых ими талломов водорослей, реже – с целью вылета (виды хирономид с осенней генерацией).
В сезонной динамике состава и количества сир-тона зарегистрировано снижение видового богатства дрифтующих донных беспозвоночных от июля к сентябрю, сопровождавшееся увеличением их биомассы. Соответственно, увеличивалась средняя индивидуальная масса дрейфующих зо-обентонтов, о чём можно судить также и по изменению состава преобладающих форм среди подёнок: в июле это были мелкие представители Baeti-dae, в сентябре – крупные представители Ephemer-idae. Доля хирономид в количественных показателях сиртона от июля к сентябрю снижалась, а доля подёнок, ручейников и олигохет – возрастала.
Видовой состав ихтиофауны перекатов исследованных рек относительно невелик. В ритрали ключевую роль в рыбных сообществах играют представители бореально-предгорного пресноводного комплекса. На перекатах потамали крупных рек ключевыми выступали понто-каспийские виды, а доля бореально-предгорного комплекса в относительных численности и ихтиомассе снижалась. На перекатах потамали малых и средних рек около 90% численности и массы рыб формировали понто-каспийские виды, а представителя бореально-предгорного ихтиофаунистического комплекса отсутствовали или были крайне немногочисленны.
В большинстве исследованных водотоков русская быстрянка формирует основу численности и биомассы ихтиофауны, превосходя по этим показателям все прочие компоненты ихтиоценозов.
Заключение
Русская быстрянка обитает в реках, значительно различающихся по гидрологическим параметрам: крупные, средние и малые водотоки, с разной длиной, площадью водосбора, скоростью течения, глубиной и т.д. При этом, во всех водотоках быстрянка занимает однотипные биотопы: перекат с песчано-гравийно-галечным дном, глубинами 0.3–0.7 м и скоростями течения в межень от 0.1 м/с в рипали до 1.0 м/с в медиали. Во всех водотоках исследованные перекаты в течение вегетационного периода обильно зарастали зелёными нитчатыми водорослями и высшей водной растительностью.
Высшая водная растительность и планктофауна перекатов изученных рек очень однообразны и слагаются небольшим числом видов. Альгофлора этого биотопа, отличаясь относительно высоким видовым богатством, имела характерную черту – обилие крупных зелёных водорослей с нитчатым талломом.
Донные сообщества, несмотря на высокую вариативность качественных и количественных параметров развития, имеют характерный облик. Видами-зообентонтами, в большей степени сопутствующими русской быстрянке, выступают массовые на изученных перекатах, но не потребляемые или потребляемые ею единично, – ручейник Psy-chomyia pusilla , клоп Aphelocheirus aestivalis , виды рода Hexatoma . Необходимыми компонентами сиртона, массово потребляемыми быстрянкой, выступают виды мошек Simulium ornatum , подёнок рода Baetis , а также хирономид родов Cricotopus и Orthocladius .
Основную роль в ихтиофауне играют реофиль-ные и реолимнофильные виды понто-каспийского пресноводного комплекса. Наиболее часто в ихтиоценозах вместе с быстрянкой встречаются уклейка и щука.
Авторы благодарят руководителя Пермского филиала ФГБНУ «ВНИРО» («ПермНИРО») А.Г. Мельникову за возможность проведения работы, и заместителя руководителя Татарского филиала ФГБНУ «ВНИРО» («ТатарстанНИРО») Ф.М. Шакирову – за помощь в организации экспедиции на территориях Республик Татарстан и Марий Эл.
Список литературы Структура речных сообществ на перекатах, населяемых русской быстрянкой (Alburnoides rossicus Berg) в восточной части ареала
- Алимов А.Ф. Элементы теории функционирования водных экосистем. СПб.: Наука, 2001. 147 с.
- Богатов В.В. Экология речных сообществ российского Дальнего Востока. Владивосток: Дальнаука, 1994. 218 с.
- Богатов В.В. Основные методы изучения дрифта речного бентоса // Чтения памяти В.Я. Леванидова. Владивосток: Дальнаука, 2005. Вып. 3. С. 5-18.
- Водоросли: справочник / С.П. Вассер, Н.В. Кондратьева, Н.П. Масюк и др. Киев: Наук. думка, 1989. 608 с.
- Денисенко С.Г. и др. Результаты оценки экологического благополучия сообществ зообентоса по индексу "разности выравненностей" (DE') // Биология внутренних вод. 2013. № 1. С. 46-55.
- Котельникова В.С. Особенности питания русской быстрянки Alburnoides rossicus Berg, 1924 // Труды Зоологического института РАН. 2016. Т. 320. С. 326-335.
- Котельникова В.С. Бентофауна и ихтиофауна переката средней равнинной реки, населяемого русской быстрянки Alburnoides rossicus Berg // Вестник Пермского университета. Сер. Биология. 2017. Вып. 3. С. 288-296.
- Методика изучения биогеоценозов внутренних водоемов / под ред. Ф.Д. Мордухай-Болтовского. М.: Наука, 1975. 240 с.
- Методические рекомендации по сбору и обработке материалов при гидробиологических исследованиях на пресноводных водоемах. Фитопланктон и его продукция / ред. Г.Г. Винберг, Г.М. Лаврентьева. Л., 1984. 32 с.
- Никольский Г.В. Частная ихтиология. М.: Высш. шк., 1971. 471 с.
- Никольский Г.В. Теория динамики стада рыб. М.: Пищевая пром-сть, 1974. 447 с.
- Одум Ю. Основы экологии. М.: Мир, 1975. 740 с.
- Песенко Ю.А. Принципы и методы количественного анализа в фаунистических исследованиях. М.: Наука, 1982. 288 с.
- Поздеев И.В., Котельникова В.С. Общая характеристика зообентоса и сиртона верхнего течения р. Чепцы // Рыбохозяйственные водоёмы России: фундаментальные и прикладные исследования: Междунар. науч. конф. СПб., 2014. С. 621-633.
- Щербина Г.Х. Годовая динамика макрозообентоса открытого мелководья Волжского плёса Рыбинского водохранилища // Зооценозы водоёмов бассейна Верхней Волги в условиях антропогенного воздействия. СПб.: Гидрометеоиздат, 1993. С. 108-144.
- Hynes H.B.N. The ecology of running waters. Toronto: Univ. Toronto Press, 1970. 555 p.
- Illies J., Botosaneanu L. Problems et methodes de la classification et de la zonation ecologique des eaux courantes, considerees surtout du point de vue faunistique // Mitt. int. Verein. theor. angew. Limnol. Stuttgart, 1963. № 12. P. 1-57.
- Kottelat M., Freyhof J. Handbook of European Freshwater fishes. Berlin, Germany, 2007. 646 p.
- R Core Team (2018). R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. URL: https://www.R-project.org/.
