Структура сообщества планктона озера Кандры-Куль летом 2010 и 2012 гг.

Автор: Уманская Марина Викторовна, Быкова Светлана Викторовна, Горбунов Михаил Юрьевич, Краснова Екатерина Сергеевна, Мухортова Оксана Владимировна, Сабитова Римма Зульфировна, Тарасова Наталья Геннадьевна, Жариков Владимир Васильевич
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Общая биология
Статья в выпуске: 2-1 т.20, 2018 года.
Бесплатный доступ
Представлены результаты исследования структуры и пространственно-временного распределения компонентов планктонного сообщества (водоросли, бактерии и зоопланктон, включающий ракообразных, коловраток и инфузорий) слабо-мезотрофного озера Кандры-Куль (респ. Башкортостан) в летние периоды 2010 и 2012 гг. Выявлена значительная пространственная и временная неоднородность как абсолютного уровня развития компонентов планктона, так и их соотношения, что может указывать на происходящую в период исследования структурную перестройку сообщества. Суммарная биомасса планктонного сообщества оз. Кандры-Куль в поверхностном слое воды в исследованный период изменялась от 18 до 1085 мг С/м3. Доля микробной «петли» в суммарной биомассе могла достигать 72% (на станциях в заросшей литорали), составляя в среднем по водоему 18%. В составе планктонного сообщества озера преобладали ракообразные, формируя 72-86% биомассы в пелагиали и 51-52% в литорали. Среди автотрофов в водоеме в целом доминировали динофитовые водоросли, в литорали к ним присоединялись зеленые, диатомовые и цианобактерии. Средняя доля автотрофов в биомассе пелагического планктона в целом и его микробной части в 2012 г. увеличилась примерно в два раза, по сравнению с 2010 г. Межгодовые изменения в структуре сообщества оз. Кандры-Куль свидетельствуют о слабой тенденции нивелирования различий между пелагиалью и литоралью и незначительном увеличении продуктивности водоёма в целом.
Структура планктона, фитопланктон, бактериопланктон, зоопланктон, инфузории, пространственно-временное распределение
Короткий адрес: https://sciup.org/148205440
IDR: 148205440 | УДК: 574.5:574.3
The structure of plankton community of the lake Kandry-Kul in the summer of 2010 and 2012
The results of the study of the structure and spatio-temporal distribution of plankton community (algae, bacteria and zooplankton, including crustaceans, rotifers and ciliates) of the oligomesotrophic lake Kandry-Kul (Rep. Bashkortostan) during the summer periods of 2010 and 2012 are presented. A significant spatial and temporal heterogeneity both of the absolute level of plankton components development and their ratio may indicate a structural transformation of the community during the study period. The total biomass of the plankton community in the surface water layer of lake Kandry-Kul varied from 18 to 1085 mg C/m3 during the study period. The contribution of microbial “loop” to the total biomass could reach 72% (in the plant-filled littoral), in average on the water body 18%. In the plankton community structure crustaceans dominated, forming 72-86% of the biomass in the pelagic zone and 51-52% in the littoral. Among the autotrophs Dinophyta dominated in the whole water body, in the intertidal zone they were joined by Chlorophyta, Bacilliariophyta and Cyanobacteria.. The average contribution of autotrophs to the biomass of pelagic plankton as a whole and its microbial part in 2012 increased approximately twice as compared to 2010. Interannual changes in the structure of plankton community of lake Kandry-Kul indicate slight tendency to decrease the differences between pelagic and littoral zones and to increase the productivity of the whole water body.
Текст научной статьи Структура сообщества планктона озера Кандры-Куль летом 2010 и 2012 гг.
Состав и уровень развития планктонного сообщества водоема определяется совокупностью
экологических факторов. Его компоненты чутко реагируют на изменение параметров окружающей среды, как на уровне отдельных организмов, так и на уровне популяций и сообществ [1, 2–4]. В водоемах с различным уровнем органического загрязнения меняется не только состав планктонных организмов, но и соотношение показателей развития его отдельных компонентов. Начиная с 80-х годов ХХ в. большое внимание уделяется изучению планктонных трофических сетей, которые наряду с обычной линейной трофической цепью фитопланктон – зоопланктон – рыбы, включают в себя так называемую микробную «петлю», состоящую из бактерий и простейших [5] и при некоторых условиях определяющую скорость и направление потоков вещества и энергии в водных экосистемах. Соотношение составляющих планктонного сообщества и вклад микробной «петли» меняется вместе с изменением трофического статуса водоема [6]. По соотношению компонентов планктона возможно оценить состояние экосистемы в конкретный момент времени и предположить характер возможных изменений в ней при различных сценариях антропогенного воздействия.
Оз. Кандры-Куль (респ. Башкортостан) – крупнейший из естественных водоемов Южного Предуралья и Средней Волги, расположенный в восточной части Бугульминско-Белебеевской возвышенности. Озеро подвержено сильной ан- тропогенной нагрузке, значительную часть которой составляет рекреационная [7, 8]. Составляющие линейной трофической цепи (фито- и зоопланктон) водоема изучали с 80-х гг. ХХ в. [9–12], исследования «микробных» компонентов планктонного сообщества (бактерии и инфузории) были проведены нами впервые [13, 14].
Цель настоящей работы – изучить структуру сообщества планктона оз. Кандры-Куль с учетом организмов микробной «петли»; установить характер пространственно-временного распределения его компонентов и их взаимоотношений.
МАТЕРИАЛЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
Отбор проб воды в оз. Кандры-Куль проводили в июне-сентябре 2010 г. на 10, и в мае-сентябре 2012 г. на 17 станциях в пелагиали, открытой литорали и в зарослях высшей водной растительности ( Hippurus vulgaris L., Chara spр., Phragmites australis (Cav.) Trin., Typha angustifolia L. и др.). Всего было отобрано 122 пробы, в каждой из которых были определены гидрохимические характеристики и параметры отдельных компонентов планктонного сообщества (фито-, протозоо-, зоо- и бактериопланктона). Отбор и анализ проб осуществляли общепринятыми методами [15–20]. Видовой состав отдельных групп гидробионтов определяли по общеизвестным определителям с учетом современных ревизий. Пересчет биомассы различных компонентов планктона в массу органического углерода проводили, используя известные коэффициенты [21, 22].
Статистический анализ данных осуществляли с помощью средств MS Excel и пакета Statistica 8.0. В качестве показателя дисперсии всех величин в статье приводятся значения стандартного отклонения. Достоверность статистических гипотез оценивали при уровне значимости α=0,05. Кластерный анализ структуры планктонного сообщества озера проводили методом Варда после трансформации величин биомасс, bi = ^Ьц/ ^jbi, что эквивалентно использованию в качестве меры различия расстояния Хеллингера [23].
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Основные абиотические факторы среды. В 2010-2012 гг. химический состав воды озера, как и в 1969 г. [24] соответствовал II типу сульфатного класса, магниевой группе. Вся водная толща в период наших наблюдений была аэробной, со слабощелочной активной реакцией воды (pH); из-за различных климатических условий в 2010 и 2012 годах термический режим в водоеме несколько различался (табл. 1). По содержанию общего фосфора, прозрачности воды и концентрации хлорофилла “а” в 2010 г. озеро четко разделялось на олиготрофную пелагиаль и ме-зотрофную литораль, а в 2012 г. вся акватория была мезотрофной. Таким образом, в период исследования состояние всего озера можно оценить как слабо мезотрофное, с некоторым трендом в сторону увеличения продуктивности [25].
Основные компоненты планктонного сообщества. Общая численность фитопланктона сильно варьировала в пространстве и во времени – 6-54502 ×103 кл./л в 2010 г., 24-26332 ×103 кл./л в 2012 г.. Общая биомасса фитопланктона изменялась от 0,8 до 5831,6 мг/м3 в 2010 г. и от 6,3 до 5733,8 мг/м3 в 2012 г. Наибольшее развитие фитопланктона было зафиксировано 02.09.10 на литоральных станциях (табл. 2). В целом в 2010 г. интенсивность развития фитопланктона в литорали была намного выше, чем в пелагиали, а в 2012 г. различия между этими двумя зонами почти сгладились (табл. 2). Сезонные изменения численности и биомассы фитопланктона в пелагиали и в литорали имели похожий характер (табл. 2). По численности во всей водной массе озера в течение всего периода наблюдений доминировали цианобактерии и зеленые водоросли и только в сентябре 2012 г., -диатомовые. Основу доминирующего комплек-
Таблица 1. Средние значения абиотических показателей в поверхностном слое воды в пелагиали и литорали озера в 2010 и 2012 гг.
|
Параметры |
Пелагиаль |
Литораль |
||||||||
|
О о гл о ГЛ |
о о гл о> о гл* о |
гл О гл О гл |
гл О гл О ГЛ |
гл О гл o> 0 0 oq |
О о oq \d 0 Xq |
О О oq o> 0 oq* О |
oq О oq О oq |
oq О oq О oq |
oq О oq o> 0 0 oq |
|
|
O 2 , мг/л |
9,8 |
8,2 |
9,1 |
7,5 |
9,4 |
8,5 |
8,8 |
8,2 |
7,0 |
10,0 |
|
T, °C |
23,4 |
18,7 |
16,9 |
25 |
14,4 |
29,5 |
15,8 |
15,6 |
24,4 |
13,9 |
|
pH |
8,7 |
8,7 |
8,6 |
8,8 |
8,7 |
8,6 |
8,6 |
8,65 |
8,8 |
8,8 |
|
Eh, mV |
345 |
– |
373 |
369 |
361 |
363 |
380 |
384 |
366 |
359 |
Примечание. «–» – нет данных са по биомассе составляли динофитовые водоросли, преимущественно Ceratium hirundinella (O. F. Muller) Bergh), в различные периоды в его состав также входили представители цианобактерий, диатомовых и зеленых водорослей.
Общая численность и биомасса бактериопланктона в 2010 г. составляли 0,4-3,8 ×109 кл./л и 50,8-865,6 мг/м3, а в 2012 г. – 0,4-4,1 ×109 кл./л и 33,0-453,0 мг/м3. Большую часть бактериопланктона озера составляли свободно плавающие одиночные клетки, однако почти в половине проб был зарегистрирован агрегированный бактериопланктон, численность которого могла достигать до 13% общей численности бактерий [13]. Уровень развития бактериопланктона озе- ра в 2010 и 2012 г. был близким, тем не менее, различия между пелагическими и литоральными станциями были более выражены в 2010 г., по сравнению с 2012г. (табл. 2). Наибольшее развитие бактериопланктона было зафиксировано в литоральных станциях – в зарослях водяной сосенки (3,8 ×109 кл./л, 2.9.10) и хары (4,1 ×109 кл./л, 21.7.12). Средняя численность и биомасса бактерий в озере в 2012 г. несколько увеличились, особенно в пелагической части (табл. 2). Однако из-за большой вариабельности выявить достоверные тренды изменений не возможно – коэффициент вариации для общей численности составил 61% в 2010 г. и 46% в 2012 г., для общей биомассы – 102% и 56%, соответственно.
Таблица 2. Средние значения численности и биомассы исследованных групп планктонного сообщества в пелагиали и литорали оз. Кандры-Куль в 2010 и 2012 гг.
|
Даты |
25.06.2010 |
02.09.2010 |
27.05.2012 |
21.07.2012 |
20.09.2012 |
|
Численность |
пелагиаль |
||||
|
Бактерии, 10 9 кл./л |
0,82±0,23 |
1,28±0,27 |
1,21±0,42 |
2,15±0,62 |
1,24±0,29 |
|
Фитопланктон, 10 3 кл./л |
67,3±94,7 |
275,3±238,3 |
162,3±270,7 |
1369,4±1133 |
188,6±157,1 |
|
Инфузории, экз/л |
392±374 |
189±88 |
685±353 |
685±759 |
2275±1581 |
|
Зоопланктон, экз/л |
191±289 |
466±210 |
421±189 |
304±229 |
112±54 |
|
Биомасса, мг/м3 |
пелагиаль |
||||
|
Бактерии |
115,4±51,6 |
107,9±19,4 |
98,6±32,7 |
214,6±97,3 |
133,2±28,9 |
|
Фитопланктон |
56,7±71,5 |
338,8±179,9 |
52,7±52,9 |
919,5±796,5 |
100,4±98,4 |
|
Инфузории |
5,1±4,7 |
2,5±1,0 |
11,5±5,8 |
8,2±8,1 |
17,2±11,3 |
|
Зоопланктон |
2179±3675 |
7799±6790 |
4349±4760 |
5659±4742 |
2223±1704 |
|
Биомасса, мг С/м3 |
пелагиаль |
||||
|
Бактерии |
13,8±6,2 |
12,9±2,3 |
11,8±3,9 |
25,8±11,7 |
16±3,5 |
|
Фитопланктон |
8,0±10,0 |
47,8±25,5 |
7,5±7,4 |
128,9±111,5 |
14,1±13,8 |
|
Инфузории |
0,6±0,5 |
0,3±0,1 |
1,3±0,6 |
0,9±0,9 |
1,9±1,2 |
|
Зоопланктон |
107,5±181,6 |
389,9±339,5 |
213,5±238 |
278,1±231,2 |
109,5±84,7 |
|
Численность |
литораль |
||||
|
Бактерии, 10 9 кл./л |
1,79±1,36 |
1,45±0,31 |
1,17±0,54 |
2,42±0,82 |
1,22±0,46 |
|
Фитопланктон, 10 3 кл./л |
1026±1183 |
17881±24893 |
123,3±163,7 |
4151±7233 |
112,8±53,7 |
|
Инфузории, экз/л |
2253±2915 |
998 |
2026±821 |
1579±1644 |
3683±2919 |
|
Зоопланктон, экз/л |
699±434 |
423±97 |
351±255 |
110±70 |
108±84 |
|
Биомасса, мг/м3 |
литораль |
||||
|
Бактерии |
248,4±260,7 |
85,8±26,7 |
143,1±70,9 |
271,3±111,5 |
143,2±69,8 |
|
Фитопланктон |
212,4±201,2 |
3020±2659 |
67,4±59,5 |
1075±1438 |
66,9±94,6 |
|
Инфузории |
21,5±23,9 |
6,0 |
34,1±18,5 |
17,1±16,2 |
20,3±14,6 |
|
Зоопланктон |
5329±4313 |
3698±2643 |
3673±4852 |
947±886 |
1815±1434 |
|
Биомасса, мг С/м3 |
литораль |
||||
|
Бактерии |
29,8±31,3 |
10,3±3,2 |
17,2±8,5 |
32,6±13,4 |
17,2±8,4 |
|
Фитопланктон |
30,7±29 |
446,8±393,6 |
9,6±8,4 |
152,6±203,7 |
9,4±13,2 |
|
Инфузории |
2,4±2,6 |
0,7 |
3,8±2,03 |
1,9±1,8 |
2,2±1,6 |
|
Зоопланктон |
261,9±213,5 |
184±130,9 |
176,7±229,7 |
46,8±43,7 |
89,6±71,3 |
Примечание. Пелагиаль – интегральные величины, литораль – только поверхностный слой
Общая численность и биомасса инфузорий в 2010 г. составляли 38-8994 экз./л и 0,9-64,9 мг/ м3, а в 2012 г. – 80-7998 экз./л и 0,7-70,6 мг/м3. Максимумы численности инфузорий были зарегистрированы в прибрежных экотопах: в 2010 г. – в зарослях тростника (8994 экз./л), в 2012 г. – в зарослях водяной сосенки (7998 тыс. экз. /м3). В 2012 г., по сравнению, с 2010 г., общая численность и биомасса инфузорий увеличилась почти вдвое [14], при этом вклад миксотрофных инфузорий снизился в зоне открытой воды и в экото-пе зарослей макрофитов, соответственно с 14% и 10% в 2010 г. до 9% и 2%, в 2012 г. Практически повсеместно в планктоне озера присутствовали мелкие виды родов Balanion, Urotricha , Askenasia , Halteria, Rimostrombidium и т.д., большинство из которых является алкалифилами [26].
Общая численность и биомасса зоопланктона в 2010 г. составляли 7,5-1305,4 экз./л и 106,5-18337,0 мг/м3, а в 2012 г. – 7,0-987,5 экз./л и 70,3-20506,6 мг/м3. В 2010 г., как и в предыдущие годы [12], сообщество зоопланктона глубоководной части озера было монодоминантным с преобладанием Diaphаnosoma brachyurum Levin. В 2010 г. В открытой литорали был обнаружен новый для водоема вид D. orghidani Negrea, который в 2012 г. вошел в ранг доминант в пела-гиали озера. Низкая численность коловраток в планктоне озера в 2010 и 2012 гг. может быть обусловлена недостатком пищевых объектов [27– 29], прессом со стороны хищного зоопланктона [30, 31], а также региональными особенностями водоемов Южного Урала [32, 33]. По сравнению с периодом олиготрофии, в видовом составе зоопланктона озера возросла доля ветвистоусых ракообразных с 36% до 49% в пелагиали и с 33% до 61% в открытой литорали, в тех же биотопах доля веслоногих сократилась с 62% до 42% и с 31% до 23%, соответственно. Зарегистрированное увеличение в 1.5–3 раза численности большинства крупных кладоцер и копепод и сдвиг структуры сообщества в сторону “укрупнения” организмов, возможно, связаны с потеплением климата [34].
Как видно из представленных данных (табл. 2), численности исследованных групп гидробионтов несопоставимы: от нескольких экземпляров (зоопланкон и инфузории) до миллионов и миллиардов клеток (водоросли и бактерии) в литре воды; в то же время величины их биомасс были более сопоставимы. Следует отметить, что статистическое распределение биомассы (и численности) всех исследованных компонентов планктонного сообщества и его суммарной биомассы было очень далеким от нормального; это видно по величинам коэффициентов вариации, приближающимся или даже превышающим 100%. Намного лучше оно соответствовало логнормальному распределению. Однако, посколь- ку нашей основной целью был анализ структуры сообществ, а переход к логарифмам из-за их неаддитивности сильно затрудняет эту задачу, мы и далее будем обсуждать в этой работе не логарифмы биомасс таксономических групп, а их абсолютные величины и доли в общем составе. Среди гетеротрофных компонентов микробной «петли» в оз. Кандры-Куль основную роль играли бактерии, незначительную – инфузории. Гетеротрофные флагелляты в большей части проб практически отсутствовали и их вклад в общую биомассу планктона в дальнейшем анализе мы не учитывали. В автотрофную составляющую «микробной петли» не включали крупноклеточные динофитовые, диатомовые и эвгленовые водоросли.
Суммарная биомасса планктонного сообщества оз. Кандры-Куль в поверхностном слое воды в 2010 г. изменялась от 0,40 до 12,67 г/м3 (18,5-1008,5 мг С/м3), а в 2012 г. – от 0,45 до 18,62 г/м3 (33,2-1085 мг С/м3). В 2010 г. биомасса планктона в пелагиали была стабильно в 3,5-3,8 раза ниже, чем в литорали (рис. 1). В мае 2012 года эта тенденция сохранилась, но различие было существенно меньше – в 1,9 раза. В июле и сентябре 2012 г. биомасса планктона в пелагиали превысила таковую в литорали. Средняя биомасса план-
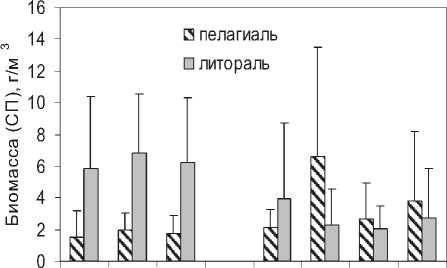
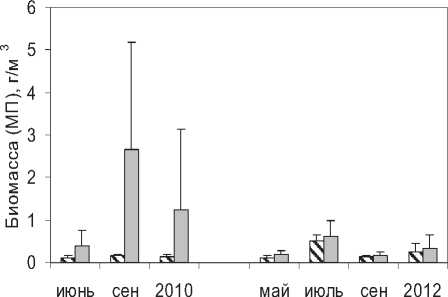
Рис. 1. Изменения суммарной биомассы планктона (СП) и ее микробных компонентов (МП) в поверхностном слое воды оз. Кандры-Куль в 2010 и 2012 гг. (показаны средние значения и стандартное отклонение)
ктона в поверхностном слое оз. Кандры-Куль несколько снизилась в 2012 г., хотя, из-за большого разброса величин по станциям, это снижение недостоверно: 2010 г. – 5,15±4,10 г/м3 с коэффициентом вариации 79,5% и в 2012 г. – 3,02±3,52 г/м3 с коэффициентом вариации 116,6%.
В среднем за период наблюдения вклад организмов микробной «петли» в суммарную биомассу планктона озера составил 18% и сильно варьировал в зависимости от времени и места расположения станции отбора (рис. 1). В целом, в поверхностном слое воды пелагических станций биомасса микробных компонентов план- ктона была относительно постоянной и мало менялась со временем (рис. 1), ее вклад в суммарную биомассу составлял 3-38% за весь период наблюдений. В литорали, где наблюдалось большее разнообразие условий (в первую очередь связанное с различным составом и степенью зарастаемости макрофитами), выявлены значительные колебания абсолютных и относительных величин биомассы микробного сообщества – 0,06+1,47 г/м3 и 1-72%, соответственно). Некоторое снижение среднего для водоема вклада организмов микробной петли в суммарную биомассу планктона с 25% (2010 г.) до 15%
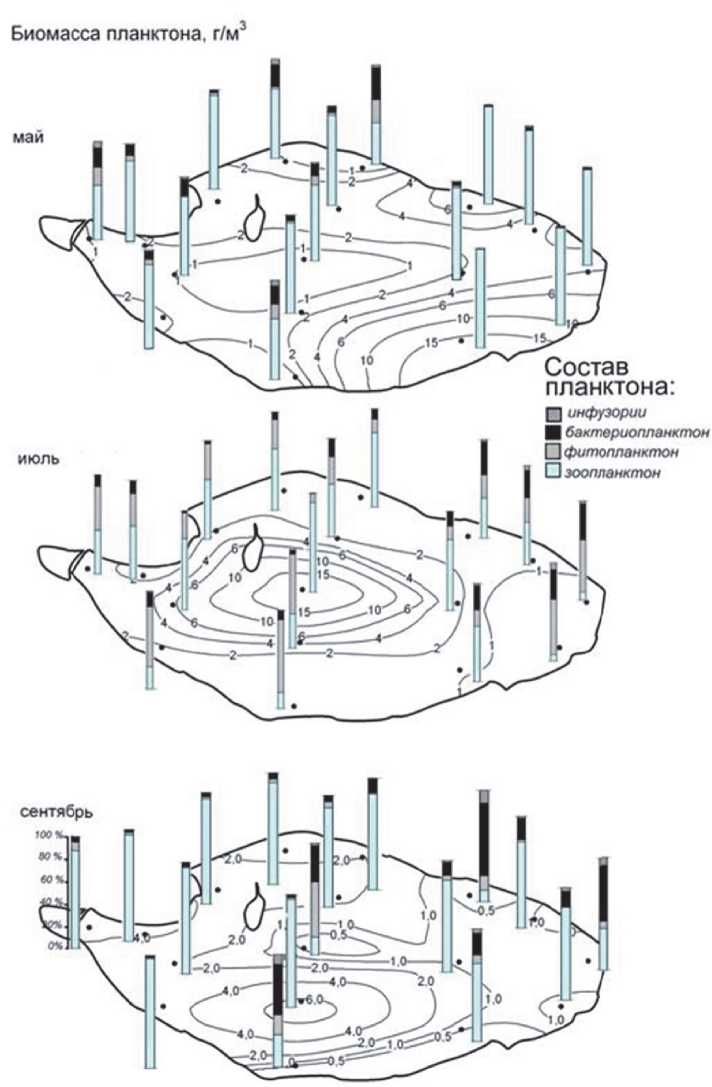
Рис. 2. Пространственно-временное распределение биомассы планктона и изменения его состава в оз. Кандры-Куль в 2012 г.
(2012 г.) может отражать происходящую эвтрофикацию, или же представляет собой артефакт из-за большого разброса величин на разных станциях.
Пространственно-временное распределение общих и структурных показателей планктонного сообщества оз. Кандры-Куль в 2012 г. (рис. 2). В мае наибольшая биомасса планктона была зарегистрирована в юго-восточной части литорали, в июле – в центральной и южной частях пелагиали, а в сентябре – только в южной части пелагиали (рис. 2).
Весной и осенью в составе планктонного сообщества явно преобладал гетеротрофный компонент сообщества, а летом вполне закономерно значительный вклад в общую биомассу практически по всей акватории озера вносил фитопланктон (рис. 2). Довольно интересен факт перераспределения микробного гетеротрофного компонента (бактерий и инфузорий) в течение сезона: весной его вклад значителен в северной и северо-западной частях озера, осенью – в восточной. Летом доли бактерий и ин- фузорий в суммарной биомассе планктона по всей акватории различались незначительно, хотя и были несколько повышены на всех литоральных станциях (рис. 2). Очевидно, эти явления отражают некую существенную пространственную неоднородность водной массы озера, однако чем именно она вызвана, пока остается неясным. В качестве возможных причин можно предложить анизотропию в строении литоральной зоны, влияние преобладающих ветров и нагонных явлений, или различное влияние и сезонные изменения характера и интенсивности антропогенной нагрузки (сельскохозяйственной и рекреационной).
Межгодовые изменения в структуре планктонного сообщества оз. Кандры-Куль в разных экотопах . Количественные показатели развития планктонного сообщества и его отдельных компонентов в пелагиали и литорали озера существенно различаются. Такие же различия, возможно, даже более выраженные, относятся и к структуре планктона, причем они проявляются как на уровне экологических груп-
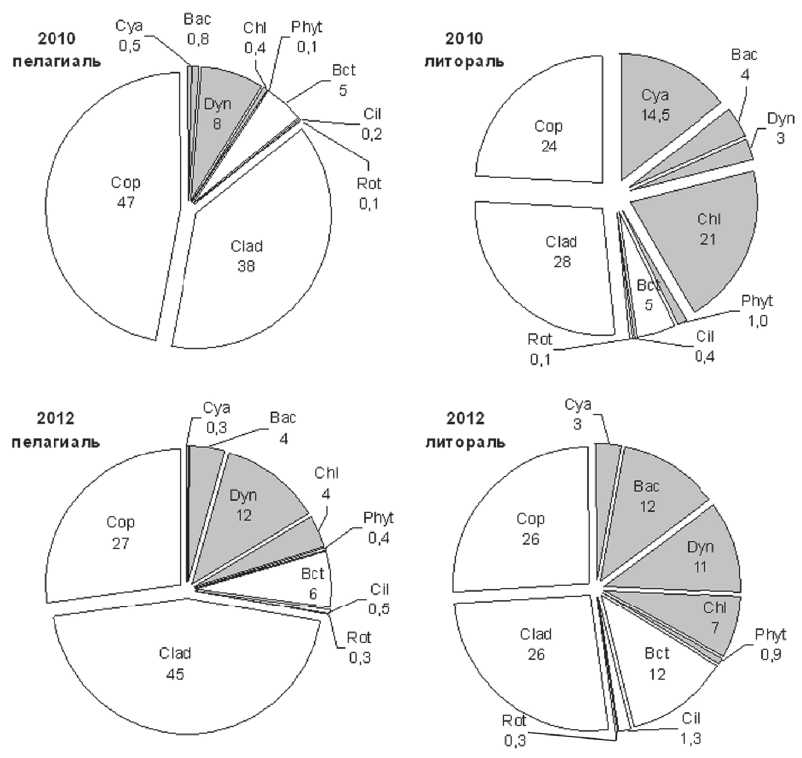
Рис. 3. Структура планктонного сообщества (в процентах от суммарной биомассы в пересчете на углерод) в разных экотопах озера в 2010 и 2012 гг.
Обозначения групп: цианобактерии–Cya, диатомовые–Bac, динофитовые–Dyn, зеленые–Chl, прочий итопланктон–Phyt; бактерии –Bct; инфузории–Cil; коловратки–Rot, кладоцеры–Clad, копеподы–Cop.
Серым цветом отмечены автотрофные компоненты планктона
пировок (фито-, зоо-, протозоо- и бактериопланктон), так и в структуре таксоценозов (крупные таксоны, напр. отделы и порядки) (табл. 2, рис. 1, 3). Необходимо отметить, что подробный анализ видовой структуры сообщества не входит в задачи данной работы; мы ограничиваемся анализом на макротаксономическом уровне.
Доля автотрофной составляющей в суммарной биомассе планктонного сообщества водоема составляла в среднем за период исследований 25,9% (27,7% в 2010 и 25,1% в 2012 г.). В 2010 г. основной вклад в биомассу пелагического планктона вносили динофитовые, а в литорали – зеленые водоросли и цианобактерии. В 2012 г. различия в структуре между пелагической и литоральной зонами несколько сгладились, на всей акватории основной вклад вносили дино-фитовые и диатомовые, а в литорали к ним добавлялись и зеленые водоросли (рис. 3). Биомасса зоохлорелл миксотрофных инфузорий играла незначительную роль в формировании общей биомассы продуцентов, в том числе зеленых водорослей.
В составе планктонного сообщества озера преобладали гетеротрофные компоненты (74%). Основной вклад в биомассу планктона в течение всего периода исследования вносил зоопланктон (66-67%), причем его доля в пелагиали была существенно больше (86% в 2010 г. и 72% в 2012 г.), чем в литорали (51-52% в оба года). В составе зоопланктона доминировали ракообразные (рис. 3). В литорали и в 2010 г., и в 2012 г. доля кладоцер и копепод оставалась практически постоянной и их соотношение составляло 1÷1. В пелагиали в 2010 г. преобладали копеподы, а в 2012 г – кладоцеры (рис. 3). В целом для водоема доля фильтраторов увеличилась с 32% в 2010 г. до 39% в 2012 г. Микрозоопланктон (инфузории и коловратки) являлся минорным компонентом планктонного сообщества. Вклад коловраток в суммарную биомассу планктона не превышал 0,2 %, а на некоторых станциях они полностью отсутствовали; вклад инфузорий – всего лишь 0,5-1,3.% (рис. 3). Тем не менее, доля последних в суммарной биомассе планктона увеличилась в 2012 г., по сравнению с 2010 г., почти вдвое (рис. 3), и, в отличие от ракообразных, это увеличение произошло синхронно и в пелагиали, и в литорали. Доля бактерий в суммарной биомассе планктонного сообщества в исследованный период составила 7,2% с увеличением вклада в 2012 г., по сравнению с 2010 г., причем оно было более выражено в литорали (рис. 3).
Средняя доля автотрофов в биомассе пелагического планктона в целом и его микробной части в 2012 г. увеличилась примерно в два раза (с 10 до 21% и с 15 до 37%, соответственно), что является одним из признаков происходящего эвтрофирования озера. Однако, в литорали вклад автотрофов снижался (рис. 3), что говорит о неоднозначности изменений, происходящих в экосистеме озера и о существенном влиянии целого ряда факторов (например, термического режима, уровня развития и видового состав макрофитов, состава фитопланктона и др.). Доля организмов микробной «петли» в суммарной биомассе планктона достоверно коррелирует с уровнем развития зоопланктона (r = -0,53) и фитопланктона (r = +0,25).
С помощью кластерного анализа структуры планктонного сообщества оз. Кандры-Куль в исследованный период было выделено 3 кластера (рис. 4). Характерно, что в состав каждого из трех кластеров вошли пробы разных лет и месяцев, и отобранные из разных биотопов. Тем не менее, в двух кластерах преобладали пробы из пелагических станций (63-67% проб), третий состоял преимущественно из литоральных проб (70%). Сезонная приуроченность проб в выделенных кластерах была выражена довольно слабо. Однако, в первом кластере собрана существенная часть проб 2010 г. и майских проб 2012 г., во втором – июльских 2012г. В третьем кластере представлены литоральные пробы практически всех дат отбора. Попытки применить в кластерном анализе не доли таксономических групп, а логарифмы их биомасс в сочетании с различными алгоритмами объединения не привели к принципиально отличающимся результатам, в т.ч. к более точному соответствию выделенных кластеров каким-либо сезонам, биотопам и т.д.
В структуре сообществ I и II («пелагических») кластеров преобладают гетеротрофные компоненты, в первую очередь ракообразные: в I кластере основными являются хищные копеподы, во II - мирные фильтраторы (кладоцеры), что обусловлено различиями в их пищевых стратегиях. Развитие кладоцер в пробах из I кластера возможно ограничено меньшим вкладом в суммарную биомассу планктона предпочитаемых кладоцерами пищевых объектов (в частности, бактерий и зеленых водорослей). В структуре сообществ III кластера преобладают автотрофные составляющие, без выраженного доминирования какой-либо одной группы. Отличительной особенностью проб этого кластера является высокий вклад цианобактерий в общую биомассу планктона. Основными чертами, резко отделяющими пробы I кластера от других, являются доминирование копепод, наименьшее развитие и разнообразие фитопланктона (практически монодоминантное сообщество динофитовых водорослей в его составе) и наименьший вклад организмов микробной «петли» (менее 7%).
В целом, в исследованный период в структуре планктонного сообщества оз. Кандры-Куль пре-
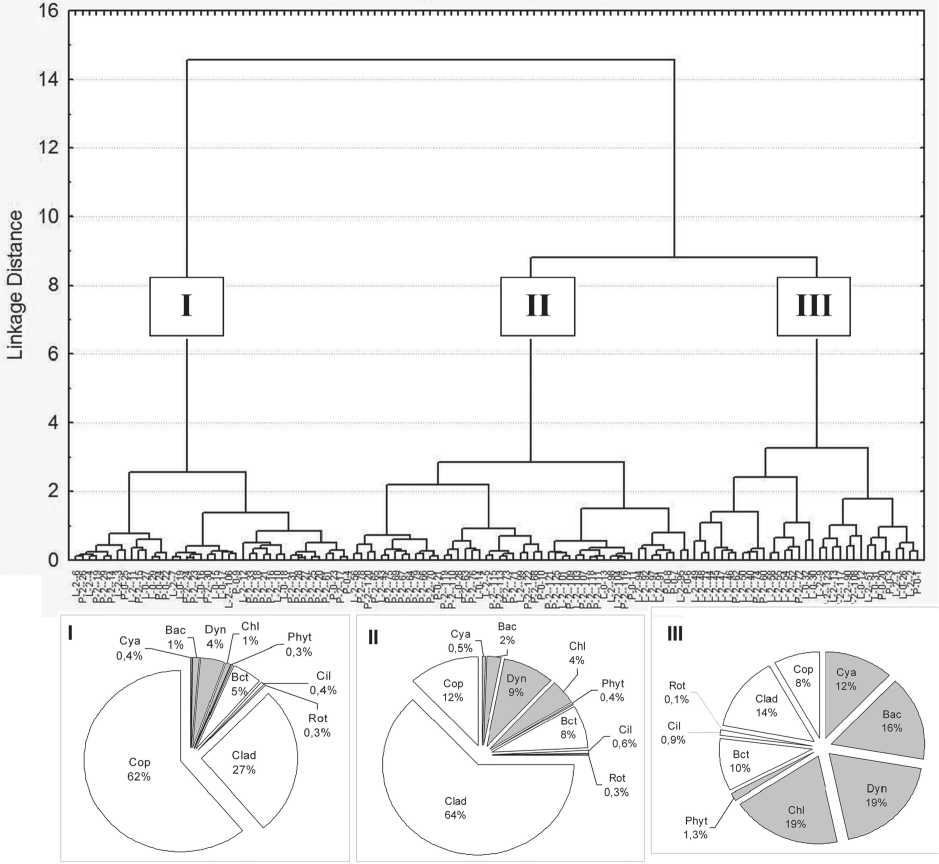
Рис. 4. Результаты кластерного анализа структуры планктонного сообщества оз. Кандры-Куль в 2010, 2012 гг.
В круговых диаграммах показана структура планктонных сообществ в разных кластерах. Обозначения – см. рис. 3.
обладают гетеротрофные компоненты и относительно невысок вклад организмов микробной «петли». Доминирование в структуре зоопланктона ракообразных указывает на эффективный контроль численности и биомассы планктонных водорослей и организмов микробной «петли» зоопланктоном [35]. Сообщество планктона оз. Кандры-Куль характеризуется значительной пространственной неоднородностью по акватории с высокой величиной варьирования между станциями: более, чем на 2 порядка для суммарной биомассы и более, чем на 3 порядка для биомассы фитопланктона. Межгодовые различия в уровне развития и распределении компонентов планктона по акватории, возможно, связаны с различными климатическими условиями в 2010 и 2012 гг., изменением степени зарастаемости литоральной зоны и состава макрофитов [36], а также с последствиями вселения в озеро азиатских карповых рыб (Hypophthalmichthys sp.) осе- нью 2010 г. Межгодовые изменения в структуре сообщества озера свидетельствуют о слабой тенденции к нивелированию различий между пелагиалью и литоралью и о незначительном увеличении продуктивности водоёма в целом.
Работа выполнена в рамках договора с природным парком «Кандры-Куль». Авторы выражают признательность коллективу парка и его директору в период исследования, С.А. Полуэктову, за интерес и техническую помощь в проведении исследования.
Список литературы Структура сообщества планктона озера Кандры-Куль летом 2010 и 2012 гг.
- Бульон В.В. Первичная продукция планктона и классификация озер//Продукционно-гидробиологические исследования пресноводных экосистем . Л.: Наука, 1987. С. 45-51.
- Трифонова И.С. Экология и сукцессия озерного фитопланктона. Л.: Наука, 1990. 184 с.
- Cairns J, McCormick P V, Niederlehner B R. A proposed framework for developing indicators of ecosystem health. Hydrobiologia, 1993. V.263, No.1. P.1-44.
- Bianchi F, Acri F, Aubry F B, Berton A, Boldrin A, Camatti E, Cassin D, Comaschi A. Can plankton communities be considered as bio-indicators of water quality in the Lagoon of Venice//Mar. Pollut. Bull. 2003. V.46, No.8. P. 964-971.
- Azam, F., Fenchel, T., Field, J. G., Gray, J. S., Meyer-Reil, L. A., Thingstad, F. The ecological role of water-column microbes in the sea//Mar. Ecol. Prog. Ser. 1983. V.10. P. 257-263.
- Auer B., Elzer U., Arndt H. Comparison of pelagic food webs in lakes along a trophic gradient and with seasonal aspects: influence of resource and predation//J. Plankton Res. 2004. V. 26, No. 6. P. 697-709.
- Государственный доклад о состоянии природных ресурсов и охране окружающей среды Республики Башкортостан в 2008 году. Уфа, 2009. 200 с.
- Государственный доклад о состоянии природных ресурсов и охране окружающей среды республики Башкортостан в 2011 году. Уфа, 2012. 367 с.
- Шкундина Ф.Б. Сезонные изменения фитопланктона озера Кандры-Куль.//Биол. науки. 1983. № 2. С. 60-64.
- Шкундина Ф. Б. Оценка степени евтрофирования оз. Кандры-Куль на основании флористического состава фитопланктона.//Гидробиол. журн. 1985. Т. 21, №2. С. 91-95.
- Шкундина Ф.Б., Гуламанова Г.А. Биологическое разнообразие автотрофного планктона озер Республики Башкортостан (Россия).//Альгология, 2011. Т. 21. № 3. C. 329-345
- Сабитова Р.З. Зоопланктон озера Кандрыкуль.//Экология водных беспозвоночных: Сб. матер. Междунар. конф. ИБВВ РАН, Борок, 30 октября-2 ноября 2010 г. Ярославль: Принтхаус, 2010. С. 267-269
- Уманская М.В., Краснова Е.С., Горбунов М.Ю. Органический пикосестон озера Кандры-Куль (респ. Башкортостан, Россия) в летний период//Изв. СНЦ РАН, 2013, Т.15,№3(7). С.2234-2239.
- Быкова С.В., Жариков В.В., Андреева В.А., Горбунов М.Ю., Уманская М.В. Инфузории озера Кандры-куль (респ. Башкортостан): Состав, пространственное распределение, сезонная динамика и экологическое состояние водоема по данным их сообщества в 2012 г.//Изв. СНЦ РАН, 2014 Т.16,№5(5). С.1748-1757.
- Унифицированные методы анализа вод/Под ред. Ю.Ю.Лурье. М.: Химия, 1973. 376 с.
- Новиков Ю.В., Ласточкина К.О., Болдина З.Н. Методы исследования качества воды водоемов. М.: Медицина, 1990. 400 с.
- Jeffrey S.W., Humfrey G.F. New spectrophotometric equations for determining chlorophylls a, b, c, and c2 in higher plants, algae and natural phytoplankton//Biochem. Physiol. Pflanz., 1975. V.167. P.191-194.
- Методика изучения биогеоценозов внутренних водоемов. М.: Наука, 1975. 240 с.
- Жариков В.В. Кадастр свободноживущих инфузорий водохранилищ Волги. Тольятти, 1996. 76 с.
- Porter K.G., Feig Y.S. The use of DAPI for identifying and counting aquatic microflora//Limnol. Oceanogr. 1980. V.25, No 5. P. 943-948.
- Копылов А.И., Косолапов Д.Б. Бактериопланктон водохранилищ Верхней и Средней Волги. М.: Изд-во СГУ, 2008. 377 с.
- Лазарева В.И. Структура и динамик зоопланктона Рыбинского водохранилища. М.: Товарищество научных изданий КМК, 2010. 183 с.
- Legendre P., Gallagher E. D. Ecologically meaningful transformations for ordination of species data//Oecologia. -2001. -Т. 129. -№. 2. -С. 271-280.
- Черняева Л.Е. Черняев А.М., Еремеева М.Н. Гидрохимия озер (Урал и Приуралье). Л.: Гидрометеоиздат, 1977. 336 с.
- Горбунов М.Ю., Уманская М.В. Современное состояние и тенденции изменения трофического статуса озера Кандры-Куль//Вода: Химия и экология, 2015. № 6. С. 3-9.
- Foissner W., Berger H., Schaumdurg J. Identification and Ecology of Limnetic Plankton Ciliates. Informationsberichte des Bayern. Hf. 3/99. Munchen: Landesamtes für Wasserwirtschaft, 1999. 793 p.
- Лазарева В.И., Жгарева Н.Н., Гусаков В.А., Иванов В.К. Структура трофической сети сообществ беспозвоночных в трех небольших озерах с различным уровнем закисления вод//Биол. внутр. вод. №1. 2003. С. 49-57.
- Lazareva V.I. Response of zooplankton communities to acidification in lakes of northern Russia//Rus. J. Aquat. Ecol. 1995. V.4, No.1. С. 41-54.
- Siegfried C.A., Bloomfild J.A., Sutherland J.W. Planktonic rotifer community structure in Adirondack, New York, U.S.A. lakes in relation to acidity, trophic status and related water quality characteristics//Hydrobiologia. 1989. V. 175, No.1. P. 33-49.
- Schindler D.W. Changes caused by acidification to the biodiversity: productivity and biogeochemical cycles of lakes//Acidification of Freshwater Ecosystems: Implication for the Future. . N.Y.: Willey & Sons, 1994. P. 153-164.
- Brakke D.F., Bohmer J., Hartmann A., Havas M., Jenkins A., Kellys S., Ormerod S.J., Paces T., Putz R., Rosseland B.O., Schindler D.W., Segner H. Physiological and ecological effects of acidification on aqutic biota//Acidification of Freshwater Ecosystems: Implication for the Future. . N.Y.: Willey & Sons, 1994. P. 275-314.
- Гареев А.М., Биккинин Р.Ф., Островская Ю.В., Сабитова Р.З., Шевченко А.М. Особенности формирования и изменчивости экологических условий в пойменных озерах (на примере оз. Шамсутдин в бирском районе Республики Башкортостан). Межведомственный сборник материалов. Уфа, 2013. С. 18-24.
- Сабитова Р.З., Мухортова О.В. Таксономическая характеристика зоопланктона озера Суртанды (Башкирское Зауралье). Экологический сборник 5: Труды молодых ученых Поволжья Междунар. конф. 2015. С. 316-318.
- Лазарева В. И., Соколова Е. А. Обеспеченность пищей планктофагов в Рыбинском Водохранилище в условиях потепления климата: динамика и продуктивность зоопланктона. Труды ИБВВ РАН, вып. 74(77), 2016. С. 77-92.
- Søndergaard, M., Jeppesen, E., Jensen J. P., Amsinck L. S. Water Framework Directive: ecological classification of Danish lakes//J. Appl. Ecol. 2005. Т. 42. №. 4. С. 616-629.
- Уманская М.В. Высшая водная растительность оз. Кандры-Куль. Самарская Лука: проблемы региональной и глобальной экологии. 2014. Т. 23, №2. С.141-145.
