Структурно-экологические признаки клубневых зонтичных (Umbelliferae) Крыма
 Крыма")
Автор: Петрова С.Е.
Журнал: Фиторазнообразие Восточной Европы @phytodiveuro
Статья в выпуске: 3 т.19, 2025 года.
Бесплатный доступ
Среди 95 изученных крымских видов зонтичных около 8% имеют гипокотильные клубни. Среди них: гипокотильноклубневые дву-, многолетние монокарпики составляют 3%, гипокотильноклубневые поликарпики – 1%, редьковидно гипокотильно-стержнекорнеклубневые дву-, многолетние монокарпики – 4% (эта группа относительно многочисленна в Крыму за счет представителей рода Trinia и Rumia crithmifolia). Метаморфизация подземных органов у гипокотильноклубневых зонтичных происходит на самых ранних этапах онтогенеза; обычно уже проростки имеют погруженный глубоко в почву клубень, образующийся за счет утолщения гипокотиля, а также длинную семядольную трубку. Группа редьковидно гипокотильно-стержнекорнеклубневых монокарпиков очень близка к гипокотильноклубневым биоморфам, однако, специализация подземных органов здесь происходит на более поздних этапах онтогенеза, хорошо развитый главный корень сохраняется до конца жизни. Виды с клубнями гипокотильного происхождения, отмеченные в Крыму, разнообразны по своей морфологии и демонстрируют разные направления структурно-экологической адаптации: у одних растений – к жизни горного лесного эфемероида (Smyrnium perfoliatum), у других – к произрастанию на открытых участках в условиях засушливого летнего периода (Bunium microcarpum, Elaeosticta lutea).
Жизненные формы, гипокотильный клубень, эфемероид, Bunium microcarpum, Chaerophyllum bulbosum, Elaeosticta lutea, Smyrnium perfoliatum, Trinia, Rumia crithmifolia
Короткий адрес: https://sciup.org/148331557
IDR: 148331557 | УДК: 581.4 | DOI: 10.24412/2072-8816-2025-19-3-303-311
Structural and ecological characteristics of tuberous Umbelliferae of Crimea
Among the 95 studied Crimean species of Umbelliferae, about 8% have hypocotyl tubers. Among them: hypocotyl-tuberous biennial and perennial monocarpics make up 3%, hypocotyl-tuberous polycarpics – 1%, radish-like hypocotyl-taproot-tuberous biennial and perennial monocarpics – 4% (this group is relatively numerous in Crimea due to representatives of the genera Trinia and Rumia). Metamorphosis of underground organs in hypocotyl-tuberous species occurs at the earliest stages of ontogenesis; usually, seedlings already have a tuber immersed deep in the soil, formed due to thickening of the hypocotyl, as well as a long cotyledon tube. The group of radish-like hypocotyl-taproot-tuberous monocarpics is very close to hypocotyl-tuberous biomorphs, however, the specialization of underground organs here occurs at later stages of ontogenesis, a well-developed main root is preserved until the end of life. Species with tubers of hypocotyl origin, noted in Crimea, are diverse in their morphology and demonstrate different directions of structural-ecological adaptation: in some plants to the life of a mountain forest ephemeroid (Smyrnium perfoliatum), in others – to grow in open areas in climate with a dry summer period (Bunium microcarpum, Elaeosticta lutea).
Текст научной статьи Структурно-экологические признаки клубневых зонтичных (Umbelliferae) Крыма
В свете изучения биоморфологического разнообразия представителей семейства Umbelliferae на территории России одним из интересных регионов представляется Крым. Зонтичные Крыма включают 95 видов и входят в десятку наиболее богатых видами семейств природной флоры республики (Yena, 2012). Крым близок к одному из основных центров разнообразия зонтичных – Средиземноморью, что важно для выявления главных направлений эволюции жизненных форм в семействе в связи с адаптивной радиацией его представителей.
Областям с прерывающейся вследствие засушливого сезона вегетацией свойственно большое разнообразие растений с запасающими органами. В данном контексте наиболее интересными структурными типами зонтичных, тесно связанными со спецификой климата Крыма, являются клубневые биоморфы, попытка изучения и классификации которых предпринята в данной статье. Органы запаса веществ позволяют растениям быть независимыми на определенных этапах развития от условий среды в течение неблагоприятного сезона, а также стремительно завершать формирование надземных органов при наступлении благоприятного времени за счет накопления в подземной сфере органических веществ. Клубневые растения часто являются эфемероидами и обладают своеобразными годичными циклами онтогенеза (Skripchinsky et al., 1968). В цикле развития монокарпических побегов у геофильных эфемероидных цветковых растений можно условно выделить три основных этапа (по Skripchinsky et al., 1968): 1) летне-осенний – клубни обычно без корней и побегов с листьями, но внутри них происходит дифференциация зачатков будущих органов очередного годичного побега, 2) осенне-зимний – сформировавшиеся в предшествующем периоде зачатки всех органов увеличиваются, выходят наружу, и многие из них вступают в непосредственный контакт с почвой, не используя, однако, сколько-нибудь активно надземной воздушной среды, 3) весенне-летний период – основные структуры монокарпического побега достигают окончательных размеров и находятся в активном контакте как с почвой, так и с воздушной средой, у поликарпиков внутри начинается дифференциация конуса нарастания побега следующего года. Все эфемероиды – геофиты, но не все геофиты – эфемероиды (Krylova, Belyanina, 1982). У зонтичных обобщенные свойства, характерные для клубневых геофитов, несколько преломляются в свете эколого-географической и филогенетической специфики самого семейства.
М АТЕРИАЛЫ И МЕТОДЫ
Изучение клубневых зонтичных, встречающихся на территории Крыма, осуществляется нами в рамках биоморфологической ревизии представителей семейства Umbelliferae. Преимущественно анализировали сборы клубневых видов из основных гербарных фондов – MW, LE, MHA, SIMF, YALT. Некоторые виды изучали в природе в Крыму: Bunium microcarpum (Boiss.) Freyn et Sint. (Крым, г. Карадаг, щебнистая осыпь на склоне), Smyrnium perfoliatum L. (Крым, г. Димерджи. Буковый лес), Trinia glauca (L.) Dumort. (Крым, г. Демерджи – Яйла, луговая степь), T. hispida Hoffm. (Крым, г. Карадаг, лугово-степные участки среди кустарниковых зарослей). В полевых и лабораторных условиях рассматривали основные морфологические особенности строения разновозрастных особей и описывали жизненные формы растений. Сравнительно-морфологический анализ проводили визуально и с помощью бинокулярного микроскопа МБС-1 в лабораторных условиях. Для изучения растений на ранних этапах развития проращивали семена, собранные автором в разных точках европейской части России ( Chaerophyllum bulbosum , Trinia hispida ) и предоставленные Ботаническим садом МГУ ( Bunium microcarpum , Ch. bulbosum ), а также анализировали проростки, найденные в природе ( Smyrnium perfoliatum ). Некоторые сведения о строении всходов Trinia , Rumia crithmifolia (Willd.) Koso-Pol. были получены из монографии Н.М. Федорончука (Fedoronchuk, 1983).
Изученные виды зонтичных распространены в Крыму в следующих местообитаниях:
Bunium microcarpum произрастает на щебнистых горных осыпях и склонах, среди скал, вдоль ущелий на полянах среди зарослей кустарников, на луговинах среди дубовых и дубовограбинниковых лесов, на щебнистых остепененных лугах, яйлах, в редких можжевеловых лесах.
Chaerophyllum bulbosum можно увидеть на сухих каменистых склонах, горных хребтах, на лесных опушках, вдоль лесных дорог, в зарослях кустарников по берегам ручьев.
Smyrnium perfoliatum обитает в горных лиственных лесах: дубовых, дубовограбинниковых, буковых, ясеневых, у ручьев среди кустарника.
Представители рода Trinia , а также близкий вид Rumia crithmifolia в целом имеют сходные местообитания. T. biebersteinii Fedoronchuk любит сухие степи и каменистые склоны, яйлы, иногда заходит в редколесье. T. hispida встречается в горах на лугово-степных участках среди кустарниковых зарослей, на каменистых склонах и в ковыльных степях, в том числе в полынно-злаковых группировках. T. glauca – на яйлах, травянистых горных склонах, в том числе степных с Stipa stenophylla , а также лугово-степных участках среди кустарниковых зарослей и лиственных лесов. Rumia crithmifolia произрастает на щебнистых склонах гор, осыпях и скалах, на выбитых петрофитных степях, в разнотравных ассоциациях, на каменистых участках в редких можжевеловых можжевелово-сосновых лесах, на лугово-степных полянах в лиственных лесах, среди кустарников по склонам хребтов
Характеристика жизненных форм клубневых растений приводится на основании понятий и методологии, предложенных нами ранее при описании биоморфологического разнообразия зонтичных Средней России (Petrova, 2016). Классификация носит комплексный характер, и разработана нами по отношению к семейству Umbelliferae в соответствии с основными положениями эколого-морфологической классификации И.Г. Серебрякова (Serebryakov, 1962), фитоценотической О.В. Смирновой с соавт. (Tsenopopulatsii, 1976), систем В.Н. Голубева (Golubev, 1984) и Е.Л. Нухимовского (Nukhimovskiy, 1997), с рядом дополнений и уточнений.
Описания сделаны по следующим признакам: 1. Тип пространственной организации и характер размещения структурных частей; 2. Характер дезинтеграции; 3. Омоложенность партикул; 4. Строение подземных побегов; 5. Наличие специализированных структур, способствующих вегетативному разрастанию, возобновлению, размножению; 6. Наличие дополнительной метаморфизации основных вегетативных органов побегов и корней, не связанной с вегетативным размножением; 7. Строение корневой системы; 8. Длительность жизни; 9. Консистенция побегов; 10. Кратность плодоношений; 11. Характер нарастания и возобновления; 12. Строение монокарпического побега; 13. Строение синфлоресценции; 14. Характер вегетации; 15. Сроки цветения; 16. Календарная продолжительность жизни.
Р ЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Среди 95 изученных крымских видов зонтичных около 8% имеют гипокотильные клубни. Среди них: гипокотильноклубневые дву-, многолетние монокарпики составляют около 3% ( Chaerophyllum bulbosum L., Elaeosticta lutea (M. Bieb. ex Hoffm.) Kljuykov, Pimenov et V.N. Tikhom., Smyrnium perfoliatum L.), гипокотильноклубневые поликарпики – около 1% ( Bunium microcarpum (Boiss.) Freyn et Sint.), редьковидно гипокотильно-стержнекорнеклубневые дву-, многолетние монокарпики – 4% (представители рода Trinia ( T. biebersteinii Fedoronchuk, T. glauca (L.) Dumort., T. hispida Hoffm. ), а также эндемичный для Крыма вид Rumia crithmifolia ).
При описании жизненных форм применяли ранее разработанный нами комплексный подход (Petrova, 2016), учитывающий до 16 признаков, представленных в «Материалах и методах».
Примеры жизненных форм изученных видов:
Bunium microcarpum – растение моноцентрическое, без дезинтеграции; глубоко гипокотильноклубневое придаточнокорневое многолетнее травянистое поликарпическое с ди-, полициклическими полурозеточными прямостоячими или при основании восходящими невысокими монокарпическими побегами, акротонно или акро-мезотонно разветвленными до 1–2 порядка по типу закрытой кисти или метелки из двойных зонтиков с открытыми зонтичками; гемиэфемероид; поздневесенне-раннелетнецветущее.
Chaerophyllum bulbosum – растение моноцентрическое, без дезинтеграции; гипокотильноклубневое стержнекорневое дву- или многолетнее травянистое монокарпическое с ди-полициклическим полурозеточным (при основании генеративного побега розетка малочленная) прямостоячим мощным монокарпическим побегом, мезотонно разветвленным до 4–5 порядка по типу метелки из двойных зонтиков с закрытыми зонтичками; способное к временному многолетнему покою в состоянии подземных клубней, в особых случаях переходящее к базисимподиальному возобновлению; виргинильный эфемероид; раннесреднелетнецветущее; возраст 2–3 года, в состоянии покоящегося клубня более 3 лет.
Smyrnium perfoliatum – растение моноцентрическое, без дезинтеграции; гипокотильноклубневое двулетнее (редко многолетнее) травянистое монокарпическое с ди-, полициклическим полурозеточным (при основании генеративного побега розетка эфемерная малочленная) прямостоячим невысоким или высоким монокарпическим побегом, акро-мезотонно разветвленным до 1–2 порядка по типу закрытой кисти или редкой метелки из двойных зонтиков с закрытыми зонтичками; гемиэфемероидное; поздневесеннецветущее.
Trinia glauca – растение моноцентрическое, без дезинтеграции; редьковидно гипокотильно-стержнекорнеклубневое дву-, многолетнее травянистое монокарпическое с полурозеточным прямостоячим невысоким, одетым при основании волокнистыми остатками отмерших листьев монокарпическим побегом, во время цветения бази-мезотонно разветвленным до 2–3 порядка по типу метелки (сжатой у мужских или шарообразной у женских особей) из несовершенных двойных зонтиков с открытыми зонтичками или из простых открытых зонтиков, собранных в 2–3 мутовки; перекати-поле; эфемероидное; поздневесенне-раннелетнецветущее. Подобной жизненной формой обладают и другие виды рода Trinia.
Метаморфизация подземных органов у гипокотильноклубневых зонтичных происходит на самых ранних этапах онтогенеза. Как правило, уже проростки имеют погруженный глубоко в почву клубень, образующийся за счет утолщения гипокотиля, а также длинную семядольную трубку (рис. A1, B1, C1). К началу генеративного периода у большинства монокарпических видов клубень находится близ поверхности субстрата. (B2, C2) У поликарпика Bunium microcarpum клубень во время репродукции довольно глубоко погружен в почву. При развитии нового монокарпического побега у B. microcarpum , такому побегу, прежде чем выйти на поверхность, какое-то время нужно расти в глубине субстрата, поэтому базальные погруженные метамеры и основания черешков листьев вегетативных розеток бывают искривленными и часто этиолированными. На многолетнем клубне прослеживаются базальные зоны в числе двух–четырех отмерших побегов предыдущих лет, нижние укороченные метамеры их входят в состав многолетнего клубня (A2). Встречаются особи, у которых одновременно развиваются и удлиненные цветоносные и вегетативный розеточный побег. У Chaerophyllum bulbosum в случае погружения клубня глубоко в субстрат при воздействии внешних условий, цветоносный побег также растет какое-то время в глубине почвы. В отдельных случаях у бутня может наблюдаться базисимподиальное возобновление (в этом случае возможен переход вида к поликарпичности). В целом, можно предположить, что размещение клубня у генеративных особей изученных видов связано с числом плодоношений: у монокарпиков клубень обычно приповерхностный, у поликарпиков – заглубленный, что вполне объяснимо с точки зрения необходимости для поликарпиков ежегодного сохранения почек возобновления в засушливый период.
Для определения возможных путей становления конкретных экологических и биоморфологических типов клубневых зонтичных важно учитывать длительность функционирования их надземных фотосинтезирующих побегов. Чаще всего клубневая биоморфа тесно связана с сокращением периода вегетации, т.е. эфемероидностью. Эфемероиды – многолетние травянистые растения, надземные побеги которых вегетируют всего 2–3 месяца, однако общая продолжительность жизни побега от заложения почки до отмирания составляет не менее 20–24 месяцев (Krylova, Belyanina, 1982). Если рассматривать эфемероиднось в соответствии с представлениями А.П. Хохрякова (Khokhryakov, 1965), что «быстрота прохождения фаз цветения и плодоношения – характерный, но недостаточный признак эфемероидности, летний перерыв вегетации – необходимый и достаточный», то нужно признать, что изученные виды зонтичных по характеру развития монокарпического побега не полностью соответствуют представлениям о настоящих эфемероидах.
Эфемероидность у растений могла возникнуть двумя путями: из ритма летнезеленого, свойственного видам мезофитных лесных и луговых сообществ, и из ритма зимне-зеленого, характерного для видов травянистых сообществ Средиземноморья (Krylova, Belyanina, 1982). В первом случае – это адаптация к ритмике мезофитных листопадных лесов, во втором – приспособление исходно зимнезеленого типа к появлению зимнего перерыва в вегетации (в то время как летний покой исходно характерен для данной группы видов) (Krylova, Belyanina, 1982).
Жизненная форма Smyrnium perfoliatum , обитающего в нижнем ярусе темных дубовых, буковых и др. лесов, по-видимому, представляет собой приспособление к жизни горного лесного эфемероида, использующего благоприятную «экологическую нишу во времени» в сообществе, где освещенность значительно меняется в ходе вегетационного сезона. У таких растений клубень, являющийся источником запасных веществ, необходим для быстрого раннего развития до распускания листьев деревьев верхнего яруса. Однако, поскольку окончательное формирование надземного побега у Smyrnium perfoliatum , а также цветение приходится на май–июнь, уместнее относить вид к гемиэфемероидам.
Остается открытым вопрос о том, следует ли рассматривать S. perfoliatum как представителя истинных лесных ранневесенних геофитов широколиственных лесов, происхождение которых многие авторы (Goryshina, 1969; Skripchinsky V., Skripchinsky Vl., 1976) считают автохтонным, связывая формирование этой группы с третичными листопадными лесами. Существуют гипотезы и об средиземноморских связях растений данного экологического типа (Goryshina, 1969). Что касается рода Smyrnium , то большинство его представителей тяготеет именно к районам Средиземноморья.
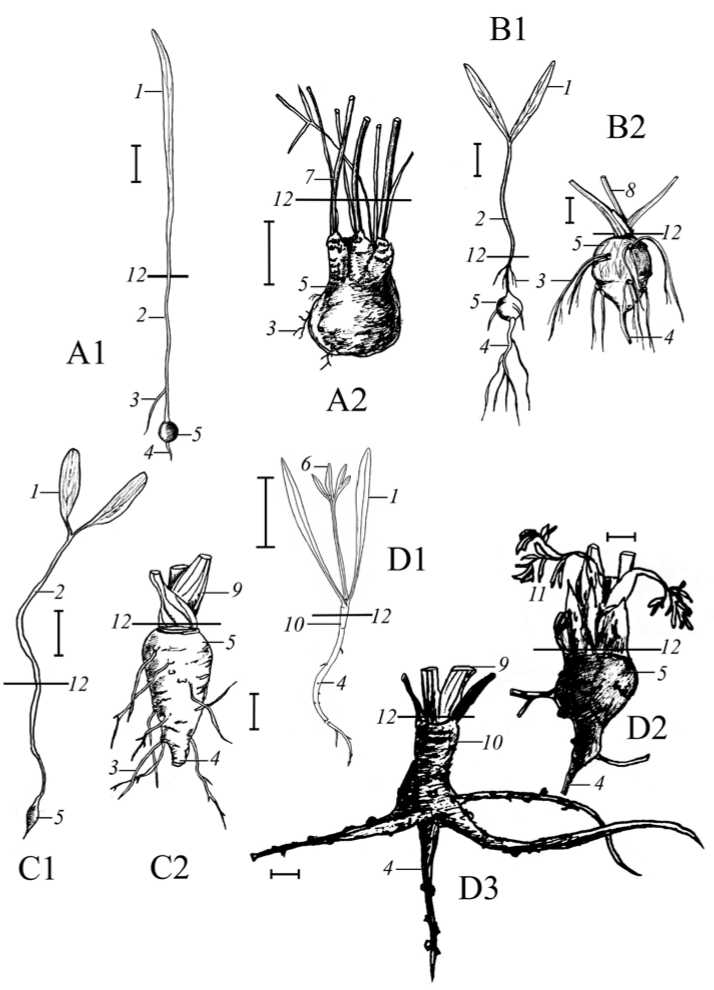
Рис. Строение клубней у гипокотильноклубневых и редьковидно гипокотильно-стержнекорнеклубневых зонтичных: A1 – проросток Bunium microcarpum , A2 – клубень генеративного растения B. microcarpum ; B1 – проросток Chaerophyllum bulbosum , B2 – клубень зрелого растения Ch. bulbosum ; C1 – проросток Smyrnium perfoliatum , C2 – клубень генеративного растения S. perfoliatum ; D1 – строение ювенильной особи в роде Trinia на примере T. multicaulis , D2 – клубень генеративного растения Trinia glauca , D3 – корневая система T. glauca , клубень не развит. 1 – семядоля, 2 – семядольная трубка, 3 – придаточный корень, 4 – главный корень, 5 – клубень, 6 – первый лист ювенильного растения, 7 – надземный стебель, 8 – черешок листа, 9 – листовое влагалище, 10 – гипокотиль, 11 – лист, 12 – уровень почвы. Мерная линейка – 1 см.
Fig. Tuber structure in hypocotyl-tuberous and radish-like hypocotyl-taproot-tuberous Umbelliferae: A1 – seedling of Bunium microcarpum , A2 – tuber of a generative plant of B. microcarpum ; B1 – seedling of Chaerophyllum bulbosum , B2 – tuber of a mature plant of Ch. bulbosum ; C1 – seedling of Smyrnium perfoliatum , C2 – tuber of a mature plant of S. perfoliatum ; D1 – structure of a juvenile plant in a genus Trinia by example of T . multicaulis , D2 – tuber of a mature plant of Trinia glauca , D3 – root system of T. glauca , tuber not developed. 1 – cotyledon, 2 – cotyledon tube, 3 – adventitious root, 4 – main root, 5 – tuber, 6 – first leaf of juvenile plant, 7 – aboveground stem, 8 – leaf petiole, 9 – leaf sheath, 10 – hypocotyl, 11 – leaf, 12 – soil level. Scale – 1 cm.
Не до конца ясно происхождение клубня у Chaerophyllum bulbosum – обитателя более светлых экотопов: лесных опушек и мезоксерофильных луговых формаций. C. bulbosum способен цвести в середине лета, и только его молодые особи обычно уходят в настоящий летний покой. Цветет C. bulbosum в июне, а плодоносит в июне–августе. В связи с этим сложно отнести C. bulbosum к истинным эфемероидам. С другой стороны, сходным образом проходит жизненный цикл, например, у некоторых видов Ferula . Так, при непродолжительной основной вегетации Ferula jaeschkeana в наиболее влажный весенне-раннелетний период фаза плодоношения приходится преимущественно на засушливый летний период, несмотря на это Ю.С. Григорьев (Grigoriev, 1980) считает это растение типичным эфемероидом. На наш взгляд, для того, чтобы внести некоторую определенность в понимание особенностей функционирования C. bulbosum , важно обратиться к изучению прегенеративного этапа развития. В виргинильном периоде надземный побег C. bulbosum функционирует весьма непродолжительное время и к концу июня часто отмирает, сохраняется лишь подземный клубень. В связи с этим растения с подобным ходом морфогенеза, на наш взгляд, уместнее называть виргинильными эфемероидами (Petrova, 2016). При изучении типа вегетации у растений, обладающих обширным ареалом, всегда необходимо иметь ввиду, что ритм развития, как и любой адаптивный признак, может быть закрепленным в различной степени (Krylova, Belyanina, 1995). Жизненная форма C. bulbosum пластична – в умеренном климате севера средней полосы России летний период покоя у вида может отсутствовать, при этом часто не формируется подземный клубень.
Bunium microcarpum и Elaeosticta lutea , распространенные на более или менее открытых сухих, часто степных или каменистых местообитаниях, развивают клубни для переживания неблагоприятного засушливого летнего периода, и, таким образом их биоморфа, как и у преобладающего большинства клубненосных зонтичных (Sdobnina, Pimenov, 1991) – это главным образом приспособление к климату с летними засухами.
Сравнение особенностей развития и вегетации у более широкого спектра таксонов данной биоморфологической группы, в частности, видов Scaligeria , Elaeosticta (Mokeeva, 1927; Pimenov, Kljuykov, 2002), позволяет сделать вывод, что группа клубневых геофильных зонтичных в настоящее время довольно разнородна по своей экологии. Наиболее правдоподобными формообразующими причинами, приведшими к возникновению у зонтичных именно клубней с преимущественным участием гипокотиля, следует, по-видимому, считать горный рельеф или открытые пространства типа степей и наличие по разным причинам перерыва в вегетации. При этих составляющих именно гипокотильный клубень становится наиболее подходящим органом (особенно у монокарпиков) для запаса веществ, сезонно приуроченного поглощения воды (за счет ежегодно обновляемых эфемерных корешков) и пережидания неблагоприятных условий в покоящемся состоянии.
Авторы В. Скрипчинский и Вл.В. Скрипчинский (Skripchinsky V., Skripchinsky Vl., 1976) отмечают, что основным фактором, определяющим эволюцию формирования эфемероидных геофитов, является очень быстрая смена кратковременного благоприятного периода года на длительный крайне неблагоприятный. Последний обычно связан с резко выраженной жарой и засухой, но может быть обусловлен и другими факторами, в частности, недостатком освещения (под пологом леса) или конкуренцией других экологически более сильных видов растений. Порядок чередования таких сезонов в разных местах может быть различен, что вызывает возникновение разных типов эфемероидных геофитов.
Группа редьковидно гипокотильно-стержнекорнеклубневых монокарпиков очень близка к предыдущей, однако, специализация подземных органов здесь происходит на более поздних этапах онтогенеза. В нее входят представители рода Trinia , а также эндемичный для Крыма вид Rumia crithmifolia . На ранних этапах развития у всех видов этой группы имеется хорошо выраженный разветвленный главный корень (рис. D1) (Fedoronchuk, 1983), утолщение гипокотиля происходит у достаточно сформированных особей (рис. D2). Лишь при переходе к репродукции главный корень перестает расти или частично редуцируется, и у многих особей (не всегда) доминирующей структурой подземной сферы становится редьковидно утолщенный гипокотиль. Иногда клубень вовсе не развивается (часто у Rumia crithmifolia ), а в подземной сфере сохраняется сильно разветвленная стержневая система, в которой однако главный корень теряется среди боковых и придаточных (рис. D3). На подземных органах отчетливо видны корнеродные выступы, к которым приурочены сезонные тонкие эфемерные корешки.
Еще В. Тролль обращал внимание на подобного рода биоморфы, относя их к реповидным геофитам (по Palamarchuk, 1948, 1956). При этом Тролль считал, что для подземных органов этих растений неприемлемо понятие стержневого корня, так как по существу они являются запасающими органами, в формировании которых принимают участие эпикотиль, гипокотиль и первичный корень, причем в различной степени. Гипокотильно-стержнекорнеклубневая жизненная форма у зонтичных – это выражение приспособленности к сухим открытым пространствам типа степей. Сами растения являются, как правило, эфемероидами или гемиэфемероидами. Побеговая структура растений этой группы организована по типу перекати-поля, то есть боковые побеги ориентированы таким образом, что создается более или менее шарообразная крона; в фазу плодоношения побег легко переламывается при основании, в результате вся надземная часть способна перемещаться ветром, парциально распространяя семена.
Если сравнивать число и процентное соотношение видов с гипокотильными клубнями, встречающихся в Крыму, с таковыми, распространенными на территории средней России (всего 3 вида, 4%, Petrova, 2016), видно, что в Крыму их больше в два раза.
З АКЛЮЧЕНИЕ
Таким образом, гипокотильноклубневые виды, произрастающие в Крыму, разнообразны по своей морфологии и демонстрируют разные направления структурной адаптации. У одних видов гипокотильные клубни могут служить приспособлением к жизни горного лесного эфемероида, когда запасающие органы необходимы для быстрого раннего развития растений до распускания листьев деревьев верхнего яруса. У других видов гипокотильные клубни формируются для переживания неблагоприятного засушливого времени года на более открытых участках в условиях климата с малом количеством осадков в летний период. Относительно многочисленна в Крыму группа редьковидно гипокотильно-стержнекорнеклубневых растений, главным образом, за счет нескольких видов рода Trinia и монотипного рода Rumia , для всех представителей данных таксонов характерно утолщение гипокотиля при развитом главном корне.
Работа выполнена по теме: Анализ структурного и хорологического разнообразия высших растений в связи с проблемами их филогении и таксономии; проблемы экологии города и устойчивого развития, № ЦИТИС: 121032500084-6.
