Структурно-продукционные характеристики водных макрофитов сбросного канала Ижевской ТЭЦ-1 (Удмуртская Республика)
")
Автор: Капитонова О.А., Тукманова С.Р.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Водные экосистемы
Статья в выпуске: 5-1 т.13, 2011 года.
Бесплатный доступ
На примере четырех характерных (модельных) видов сообществ Lemno-Typhetum angustifoliae, Typheto angustifoliae-Phragmitetum autsralis и Lemno minori-Spirodeletum показано влияние подогретых вод Ижевской ТЭЦ-1 на продукционные и анатомо-морфологические характеристики водных макрофитов. Выявлено увеличение биомассы, ускорение сезонного развития, изменение структурных параметров водных и прибрежно-водных растений.
Тепловое загрязнение, водоем-охладитель, макрофиты, гидрофиты, гелофиты, продуктивность, морфология, анатомия
Короткий адрес: https://sciup.org/148200328
IDR: 148200328 | УДК: 581.522.5
Structural and production characteristics of aquatic plants the waste channel of the Izhevsk thermal power station-1 (the Udmurt Republic)
On an example of four characteristic (modelling) species of communities Lemno-Typhetum angustifoliae, Typheto angustifoliae-Phragmitetum autsralis and Lemno minori-Spirodeletum influence of the warmed-up waters of the Izhevsk thermal power station-1 (Izhevsk, Udmurtia) on production-powered and anatomic-morphological characteristics of aquatic macrophytes are presented. The biomass increase, acceleration of seasonal development, change of structural parameters of vegetative bodies and inflorescences of hydrophytes and helophytes is revealed.
Текст научной статьи Структурно-продукционные характеристики водных макрофитов сбросного канала Ижевской ТЭЦ-1 (Удмуртская Республика)
П ресноводные водоемы-охладители тепловых электростанций вызывают интерес гидробиологов в связи поступлением дополнительного тепла в водные экосистемы, влияющего на структурные и продукционные характеристики гидробионтов и их сообществ. Для живых организмов повышение температуры среды обитания является стрессовым фактором, вызывающим интегральный неспецифический ответ, направленный на выживание биосистем за счет формирования защитных механизмов [13, 18]. Согласно H. Selye [17], в реагировании организма на стрессор – фактор, вызывающий состояние стресса – можно выделить три стадии: стадию тревоги, направленную на мобилизацию защитных сил биосистемы, стадию резистентности, означающую приспособление к возникшей ситуации, и стадию истощения, которая может привести к смерти биосистемы в случае сильного и длительного стресса. Растения, обитающие в водоемах-охладителях в условиях хронического воздействия температурного фактора относительно невысокой интенсивности, не имея возможности «уйти» от влияния стрессора, фактически вынуждены приспосабливаться к нему, мобилизуя свои внутренние защитные ресурсы.
Адаптация биосистем к воздействию экологического фактора происходит на разных уровнях организации. На субклеточном и клеточном уровне под влиянием повышенной температуры наблюдаются модификации в хлоропластах, накопление белков теплового шока, кислородсодержащих радикалов, липидов [16, 19, 20]. На уровне организма адаптация выражается в уменьшении размеров клеток и увеличении их плотности, увеличении проницаемости мембран, уменьшении испарения [20]. В отношении акваль-ных экосистем значимыми параметрами, пригодными для целей биоиндикации и экологического мониторинга, являются показатели
структурно-функциональных изменений биосистем на уровне организма, популяции и сообщества [9]. Многочисленными исследованиями [4, 7 и др.] показано увеличение биомассы сообществ гидрофитов в зоне теплового загрязнения и преимущественное распространение в ней растений определенных экологических и таксономических групп.
Цель работы – изучение влияния теплового загрязнения на характерные (модельные) виды гидрофильных сообществ – Phragmites australis (Cav.) Trin. ex Steud., Typha angustifolia L., Lemna minor L. и Spirodela polyrhiza (L.) Schleid.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Район исследований – Ижевское водохранилище, расположенное на р. Иж. Водохранилище заполнено в 1760 г. для нужд строящегося железоделательного завода, площадь водного зеркала в настоящее время составляет 24 км2, средняя глубина – 3,5 м, в приплотинной части – 12 м, полный объем водной массы 76,3 млн. м3. Водная и прибрежно-водная растительность сосредоточена в основном в верховьях водоема и на мелководных участках по периметру. Исследования проводили в течение 2003 - 2007 гг. с мая по октябрь. Опытный участок находился в зоне прямого контакта подогретых вод. Он представляет сбросной канал Ижевской ТЭЦ-1, имеет длину 120 м, ширину в месте сброса теплых вод 12 м, в средней части – 20-25 м. Площадь канала составляет 0,002 км2, глубина колеблется от 0,5 до 1,0 м. Канал отделен от основной части водохранилища земляным валом.
В качестве контроля выбран один мелководный участок водохранилища площадью 0,001 км2, расположенный у правого берега, в 1 км выше по течению от опытного участка (рис.1). С обоих участков водохранилища в 2004 г. были отобраны пробы воды для химического анализа на содержание ионов металлов (Al, Mn, Fe, Ni, Cu, Cr, Mg), выполненного в лаборатории почвенной экологии Удмуртского государственного университета по стандартизированным методикам.
Для решения поставленной задачи были выделены наиболее распространенные сообщества водных и прибрежно-водных растений Ижевского водохранилища и сбросного канала с использованием традиционных подходов и методов отечественной фитоценологии.
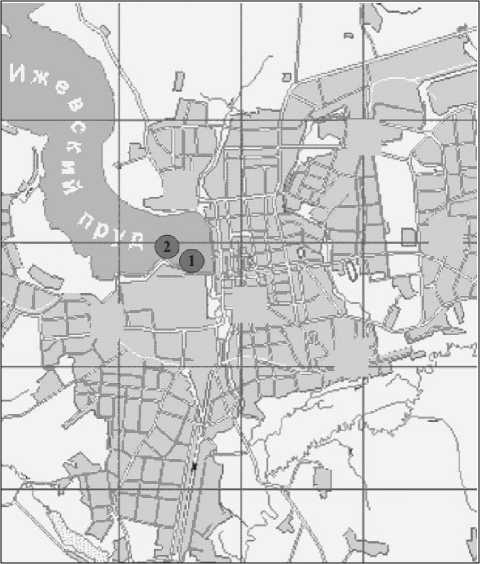
Рис. 1 . Расположение опытного (1) и контрольного (2) участков на Ижевском водохранилище
Объектами исследований послужили четыре вида макрофитов – Phragmites australis (Cav.) Trin. ex Steud., Typha angustifolia L., Lemna minor L. и Spirodela polyrhiza (L.) Schleid. Указанные виды на изученных участках водоёма являются ценозо-образователями. Из них первые два вида, согласно широко используемой отечественными гидроботаниками эколого-биоморфологической класс-сификации водных и прибрежно-водных растений [12], относятся к экологической группе высокотравных гелофитов, или воздушно-водных растений, а последние два являются представителями свободно плавающих на поверхности воды гидрофитов, относящихся к настоящим водным растениям.
Стандартные укосы для определения надземной биомассы сообществ гелофитов и гидрофитов проводили на участках с глубиной воды 0,5 м во второй половине августа, когда фитомасса достигала максимума. Для анатомо-морфологических измерений проводился сбор растительного материала. Листья свежесобранных растений рогоза узколистного, а также фронды ряски малой и многокоренника обыкновенного фиксировали в 50%-ном этиловом спирте для последующего лабораторного анализа строения эпидермальной ткани с использованием методов, описанных в литературе [2, 8, 11].
За весь период исследований было сделано 107 геоботанических описаний, в том числе 67 в опыте и 40 в контроле, произведено 72 укоса надземной биомассы сообществ гелофитов и гидрофитов, отобрано 3600 образцов для проведения морфологического и 3000 образцов для проведения анатомического анализа, выполнено 10200 измерений морфологических и 15000 подсчетов анатомических параметров. Линейные измерения и подсчет структурных элементов эпидермы проводили с помощью микроскопа МБД-1 при увеличении х 640 с применением окулярного винтового микрометра МОВ-1-16 х . По результатам всех анализов проведен стандартный статистический анализ [6] с использованием персонального компьютера IBM РС. Статистическая обработка данных проводилась в программной среде Statistica v.6.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Анализ данных по содержанию приоритетных загрязняющих веществ в воде показал отсутствие достоверных различий между опытным и контрольным участками исследованного водохранилища по уровню химического загрязнения (табл. 1), несмотря на бόльшие значения некоторых показателей на опытном участке. Это позволяет утверждать, что основным фактором, воздействующим на модельные растения, является температурный, причем за весь период наблюдений разность температур воды опытного и контрольного участков составляла в среднем 10-11 ° С. Максимальная температура в «теплых водах» зафиксирована в 2003 г. и составила +36˚С. Данные по изменению температуры воды сбросного канала в течение сезона представлены на рис. 2.
Таблица 1 . Основные химические показатели воды сбросного канала Ижевской ТЭЦ-1 (опыт) и Ижевского водохранилища (контроль) (2004 г.)
|
Показатели |
Норматив |
Опыт (M±m) |
Контроль (M±m) |
|
pH |
6-9 |
8,08±0,42 |
8,05±0,42 |
|
Al, мг/л |
0,5 |
0,046±0,031 |
0,052±0,035 |
|
Fe, мг/л |
0,3 |
0,0472±0,0097 |
0,0305±0,0061 |
|
Cu, мг/л |
1,0 |
0,0028±0,0012 |
0,0019±0,0007 |
|
Cr, мг/л |
0,05 |
0,007±0,001 |
0,001±0,0005 |
|
Mn, мг/л |
0,1 |
0,29± 0,003 |
0,012±0,001 |
|
Ni, мг/л |
0,1 |
0,005±0,001 |
0,001±0,0005 |
|
Mg, мг/л |
Не установлен |
15,1±1,2 |
16,5± 1,3 |
Исследованный канал мелководный, что обуславливает его высокую степень зарастания, достигающую 80-90%. Наиболее характерными сообществами, участвующими в зарастании канала, являются ценозы рогоза узколистного с рясками
Lemno-Typhetum angustifoliae, тростника южного с рогозом узколистным Typheto angustifoliae-Phragmitetum autsralis и рясковых Lemno minori-Spirodeletum. Они приурочены к прибрежьям и устьевой части канала. Схема зарастания сбросного канала представлена на рис. 3
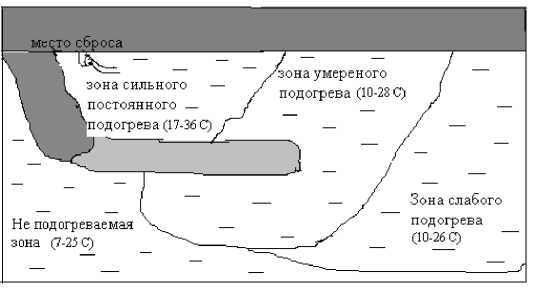
Рис. 2 . Термический режим сбросного канала
Ижевской ТЭЦ-1
Выявлено превышение надземной биомассы сообществ макрофитов в зоне теплового загрязнения по сравнению с контрольным участком (табл. 2). Выявленные различия указывают на кажущиеся благоприятные условия для роста и развития изученных растений в условиях искусственного подогрева воды. Однако в основном это достигается за счет густоты зарослей: этот показатель в «теплых водах» выше на 5-30 % побегов, что отражено и в фенологических спектрах (рис. 4). Наряду с увеличением доли вегетативных побегов в опытных популяциях, имеет место также сдвиг в развитии растений в среднем на 10-15 дней с одновременным укорачиванием фенофаз. Таким образом, перестройка популяционной структуры модельных видов, выражающаяся в изменении соотношения возрастных групп в сторону преобладания особей прегенеративных стадий развития, отражается и на продуцировании надземной биомассы формируемых ими сообществ, которая в среднем в 1,3-1,4 больше (по воздушно-сухому весу) на участке с подогретой водой.
В результате изучения особенностей морфологического строения растений модельных видов установлено, что повышение температуры водной среды действует в целом благоприятно на Phragmites australis, опытные растения которого отличались от контрольных более крупными размерами листьев и соцветий. В то же время обнаружено отрицательное влияние повышенной температуры на развитие генеративной сферы Тypha angustifolia (табл. 3).
из-за усиленного роста вегетативных
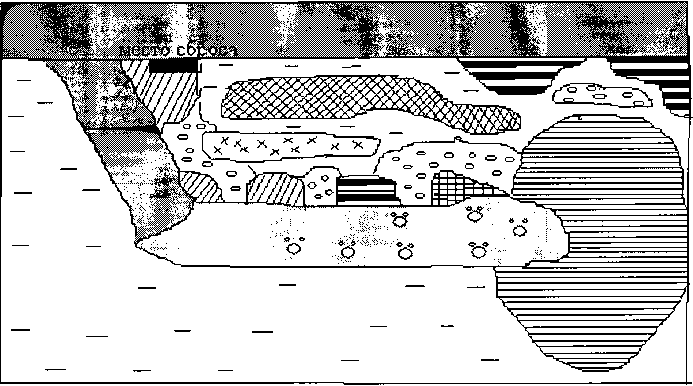
Рис. 3. Схема зарастания сбросного канала Ижевской ТЭЦ-1
Potametum nervigeri
Ceratophylletum demersi
Lemno minori-Spirodeletum
Условные обозначения:
Lemno-Typhetum angustifoliae
Heteroherboso-Typhetum latifoliae
Typhetum latifolia-angustifolia
Typheto angustifoliae-Phragm itetum autsralis земляная насыпь
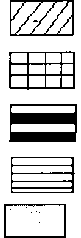

бетон
открытая вода
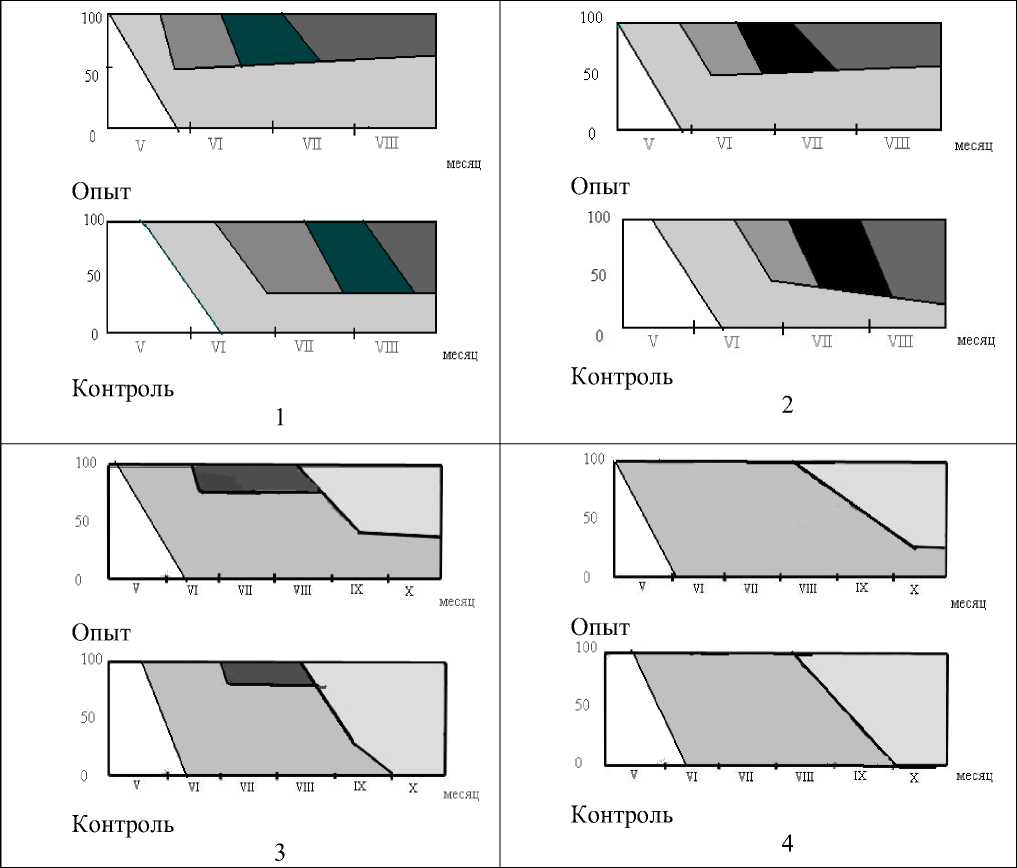
Рис. 4. Фенологические спектры T. аngustifolia (1), P. australis (2), L. minor (3) и S. polyrhiza (4) на Ижевском водохранилище (2003-2006 гг.). Условные обозначения:
вегетация
бутонизация
цветение

плодоношение
отмирание
Статистически значимые различия получены также по некоторым морфологическим параметрам Spirodela polyrhiza и Lemna minor (табл. 3). Согласно данным ряда авторов [5, 14, 15], эпидермальные клетки корня рясковых выполняют функцию поглощения, поэтому можно предположить, что выявленное увеличение длины корней у опытных растений ряски и многокоренника на фоне уменьшения площади фронда способствует поглощению дополнительного количества питательных веществ и кислорода, растворимость которого в тёплой воде существенно снижается, а потребность в нём повышается. Ранее нами было отмечено противоположное явление – укорачивание длины корня Lemna minor в условиях загрязнения водоемов тяжелыми металлами [3]. Таким образом, наблюдаемое в «теплых водах» удлинение корней рясковых отражает их реакцию на повышение температуры воды и может быть использовано в
Таблица 2 . Надземная биомасса изученных фитоценозов, г/м2
|
Ассоциация |
Участок |
N |
Сырой вес (M ± m) |
Воздушносухой вес (M ± m) |
|
Typheto angustifoliae-Phragmitetum australis |
опыт |
20 |
2860,00 ± 74,83 |
698,00 ± 64,84 |
|
контр. |
20 |
2180,00 ± 96,95 |
508,00 ± 17,72 |
|
|
Lemno minori-Spirodeletum |
опыт |
16 |
622,35 ± 103,7 3 |
33,60 ± 6,69 |
|
контр. |
16 |
520,00 ± 49,96 |
26,80 ± 3,48 |
Примечание: здесь и в последующих табл. полужирным шрифтом отмечены значения параметров, имеющие статистически значимые (р<0,05) различия в опыте и контроле в экомониторинговых исследованиях при оценке экологического состояния поверхностных вод. Удлинению корней как органа равновесия рясковых может способствовать также уменьшение вязкости воды при повышении ее температуры, что позволяет этим гидрофитам образовывать плотные ковры на поверхности воды, предотвращая переворачивание отдельных растений.
Таблица 3 . Показатели морфологического строения высокотравных гелофитов (N=300) на опытном и контрольном участках
|
Показатели |
Участок |
(M±m) |
|
Ty |
pha angustifolia |
|
|
Длина листа, см |
опыт |
172,38±2,29 |
|
контроль |
137,80±0,76 |
|
|
Ширина листа, см |
опыт |
1,28±0,02 |
|
контроль |
1,27±0,01 |
|
|
Длина пестичной части соцветия, см |
опыт |
15,04±0,18 |
|
контроль |
19,13±0,15 |
|
|
Диаметр пестичной части соцветия, см |
опыт |
1,59±0,02 |
|
контроль |
1,64±0,02 |
|
|
Длина тычиночной части соцветия, см |
опыт |
12,61±0,06 |
|
контроль |
16,88±0,16 |
|
|
Диаметр тычиночной части соцветия, см |
опыт |
1,48±0,02 |
|
контроль |
1,45±0,02 |
|
Phragmites australis
|
Длина листа, см |
Опыт |
40,07±0,11 |
|
Контроль |
37,48±0,20 |
|
|
Максимальная ширина листа, см |
Опыт |
2,40±0,05 |
|
Контроль |
2,24±0,01 |
|
|
Длина соцветия, см |
Опыт |
30,19±0,35 |
|
Контроль |
25,15±0,22 |
Таблица 4 . Показатели морфологического строения гидрофитов исследованных участков (N=600)
|
Показатели |
Участок |
Lemna minor (M±m) |
Spirodela polyrhiza (M±m) |
|
Длина |
опыт |
2,91±0,08 |
6,02±0,08 |
|
фронда, мм |
контроль |
2,77±0,08 |
6,06±0,08 |
|
Ширина |
опыт |
1,37±0,05 |
5,31±0,06 |
|
фронда, мм |
контроль |
1,39±0,06 |
5,33±0,05 |
|
Площадь |
опыт |
3,44±0,13 |
25,39±0,46 |
|
фронда, мм2 |
контроль |
3,61±0,12 |
24,84±1,04 |
|
Длина |
опыт |
36,65±1,42 |
9,11±0,12 |
|
1 корней, мм |
контроль |
32,65±1,26 |
8,62±0,15 |
Различия во внешнем строении изученных видов гидрфитов проявляются и на анатомическом уровне, причём наблюдается отчётливая тенденция к уменьшению линейных размеров структурных элементов эпидермы и, как следствие, более плотному их расположению на единице поверхности у опытных растений. Установлено статистически значимое увеличение плотности устьиц у L. minor и S. polyrhiza , а у первого вида, кроме того, уменьшение длины устьиц и ширины основных эпидермальных клеток (табл. 5).
Значимые различия выявлены и в строении эпидермы листьев T. angustifolia (табл. 6). Указанные особенности внешнего и внутреннего строения изученных растений индицируют качество среды их обитания и, вероятно, указывают на стимулируемое высокой температурой ускорение клеточных делений с одновременным торможением ростовых процессов в вегетативных органах. Мелкоклеточность эпидермы является неспецифической реакцией и, по-видимому, характерна как для водных растений в условиях изменения параметров среды их обитания, так и для наземных растений под воздействием разнообразных факторов: сильного освещения [1], пониженной температуры [10], недостаточного обеспечения элементами минерального питания [8].
|
Таблица 5. Показатели анатомической структуры гидрофитов исследованных участков (N=600) |
|||
|
Показ атели |
Участок |
Lemna minor, (M±m) |
Spirodela polyrhiza, (M±m) |
|
L верх , мкм |
опыт |
35,30±0,14 |
44,81±0,52 |
|
контр. |
35,31±0,12 |
45,70±0,43 |
|
|
D верх , мкм |
опыт |
17,02±0,34 |
23,46±0,49 |
|
контр. |
18,15±0,19 |
24,37±0,39 |
|
|
l верх , мкм |
опыт |
21,35±0,37 |
35,56±0,65 |
|
Контр. |
23,19±0,22 |
36,66±0,71 |
|
|
d верх , мкм |
опыт |
17,45±0,78 |
18,94±0,17 |
|
контр. |
17,22±0,16 |
18,88±0,17 |
|
|
N верх |
опыт |
1820,50±24,91 |
1747,65±8,41 |
|
контр. |
1795,50±14,85 |
1739,45±7,40 |
|
|
n верх |
опыт |
83,25±1,30 |
84,60±0,74 |
|
контр. |
75,02±0,97 |
76,25±0,87 |
|
Примечание: здесь и в табл. 6 приняты следующие обозначения: Lверх (Lниж) – длина основной эпидермальной клетки верхней (нижней) эпидермы, Dверх (Dниж) – ширина основной эпидермальной клетки верхней (нижней) эпидермы, lверх (lниж) – длина устьица на верхней (нижней) стороне листа, dверх (dниж) – ширина устьица на верхней (нижней) стороне листа, Nверх (Nниж) – число основных эпидермальных клеток на 1 мм2 поверхности верхней (нижней) эпидермы; nверх (nниж) – число устьиц на 1 мм2 поверхности верхней (нижней) эпидермы.
Таблица 6. Показатели анатомической структуры листа Typha angustifolia (N=300)
|
Признак |
Участок |
(M±m) |
|
L верх, мкм |
опыт |
1,14±0,32 |
|
контроль |
1,94±0,16 |
|
|
L ниж, мкм |
опыт |
0,99± 0,31 |
|
контроль |
0,92±0,35 |
|
|
D верх, мкм |
опыт |
0,67±0,14 |
|
контроль |
0,65±0,09 |
|
|
D ниж, мкм |
опыт |
0,49±0,17 |
|
контроль |
1,11±0,05 |
|
|
l верх, мкм |
опыт |
1,29±0,28 |
|
контроль |
1,69±0,21 |
|
|
l ниж, мкм |
опыт |
1,19±0,17 |
|
контроль |
1,48±0,17 |
|
|
d верх, мкм |
опыт |
1,22±0,28 |
|
контроль |
1,39±0,05 |
|
|
d ниж, мкм |
опыт |
0,66±0,23 |
|
контроль |
1,83±0,14 |
|
|
N верх |
опыт |
566,65±26,90 |
|
контроль |
180,18±14,08 |
|
|
N ниж |
опыт |
307,06±28,98 |
|
контроль |
202,70±21,91 |
|
|
n верх |
опыт |
139,64±14,91 |
|
контроль |
72,07±10,98 |
|
|
n ниж |
опыт |
130,63±13,90 |
|
контроль |
49,55±6,04 |
Список литературы Структурно-продукционные характеристики водных макрофитов сбросного канала Ижевской ТЭЦ-1 (Удмуртская Республика)
- Горышина Т.К. Растение в городе. Л.: ЛГУ, 1991. 152 с.
- Захаревич С.Ф. К методике описания эпидермиса листа//Вестн. ЛГУ. Сер. Биология. 1954. ¢ 4. С. 65-75.
- Капитонова О.А. Макрофиты в условиях промышленной среды. Ижевск, 2007. 168 с.
- Катанская В.М. Растительность водохранилищ-охладителей тепловых электростанций Советского Союза. Л.: Наука, 1979. 277 с.
- Комаров В.Л. Практический курс анатомии растений: Учеб. пособие для ун-ов, пед. и с.-х. ин-ов. М.-Л.: Изд-во АН СССР, 1941. 312 с.
- Лакин Г.Ф. Биометрия. М.: Высш. шк., 1990. 352 с.
- Лукина Л.Ф., Смирнова Н.Н. Физиология высших водных растений. Киев: Наукова думка, 1988. 188 с.
- Мирославов Е.А. Структура и функция эпидермиса листа покрытосеменных растений. Л.: Наука, 1974. 184 с.
- Моисеенко Т.И. Концепция «здоровья» экосистемы в оценке качества вод и нормирования антропогенных нагрузок//Экология. 2008. ¢ 6. С. 411-419.
- Николаевская Е.В. Изменчивость морфолого-анатомических признаков строения листа разных экотипов Trifolium repens L. в связи с вертикальной зональностью//Вестн. ЛГУ. Сер. 3 (Биология). 1990. Вып. 4 (¢24). С. 33-44.
- Николаевский В.Г. К методике количественно-анатомического изучения влияния внешней среды на структуры вегетативных органов высших растений//Бот. журн. 1964. Т. 49, ¢ 6. С. 833-838.
- Папченков В.Г. Растительный покров водоемов и водотоков Среднего Поволжья, Ярославль: ЦМП МУБиНТ, 2001. 200 с.
- Пятыгин С.С. Стресс у растений: физиологический подход//Журн. общ. биол. 2008. Т. 69, ¢ 4. С. 294-298.
- Ростовцев С.И. Биолого-морфологический очерк рясок. М., 1905. 108 с.
- Фурст Г.Г. Анатомическое строение некоторых водных растений//Бюл. Главн. ботан. сада АН СССР. 1968. Вып. 71. С. 67-74.
- Iba K. Acclimative Response to Temperature Stress in Higher Plants: Approaches of Gene Engineering for Temperature Tolerance//Plant Biol. 2002. N 53. P. 225-245.
- Selye H. A Syndrome Produced by Diverse Nocuous Agents//Nature. Vol. 138, July 4. 1936. P. 32.
- Sung D.-Y., Kaplan F., Lee Kil-ae, Guy C.L. Acquired tolerance to temperature extrems//TRENDS in Plant Science. Vol. 8. N 4, April, 2003. P. 179-187.
- Suzuki N., Mittler R. Reactive oxygen species and temperature stresses: A delicate balance between signaling and destruction//Phisiologia Plantarum. 126. 2006. P. 45-51.
- Wahid A., Gelani S., Ashrat M., Foolad M.R. Heat Tolerance in plants: An overview//Environmental and Experimental Botany. 2007. N 61. P. 199-223.
