Структурные механизмы становления симметрии у микроспориальных эмбриоидов пшеницы: данные сканирующей электронной микроскопии

Автор: Сельдимирова Оксана Александровна, Титова Галина Евгеньевна, Галин Ильшат Рафкатович, Круглова Наталья Николаевна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Биотехнология
Статья в выпуске: 3-5 т.15, 2013 года.
Бесплатный доступ
Методом сканирующей электронной микроскопии проанализированы фенотипы эмбриоидов, формирующихся в культуре in vitro пыльников яровой мягкой пшеницы. Выявлены структурные механизмы формирования эмбриоидов и рассмотрена роль синтетического ауксина 2,4-Д в индукции их образования.
Культура пыльников in vitro, эмбриоиды, полиэмбриоиды, 4-д, симметрия
Короткий адрес: https://sciup.org/148202071
IDR: 148202071 | УДК: 633.111.1:581.143.6
Structural mechanisms of symmetry establishment in microspore derived embryoids of wheat: the data of scanning electron microscopy
The phenotypes of microspore derived embryoids formed in anther culture in vitro of spring soft wheat were analyzed by the method of scanning electronic microscopy. Structural mechanisms of embryoids formation are revealed and the role of synthetic auxin 2,4-D in their induction is considered.
Текст научной статьи Структурные механизмы становления симметрии у микроспориальных эмбриоидов пшеницы: данные сканирующей электронной микроскопии
Морфогенез, как совокупность протекающих в развивающемся организме процессов дифференциации клеток с образованием специализированных тканей и органов [1], остается одной из сложнейших проблем биологии развития растений.
При морфогенезе определяющее значение принадлежит процессам становления полярности и симметрии растительного организма, так как осевая организация тела в пространстве – существенный элемент целостности растительного организма [2, 3].
Однако реальные процессы, в том числе и структурные механизмы, лежащие в формирования осевой симметрии у высших растений, изучены недостаточно [2]. Основная причина этого – недоступность зародышей растений на самых ранних этапах их развития [4].
Удобной модельной системой для исследований в области морфогенеза растений может служить соматический эмбриогенез – формирование биполярных зародышеподобных структур – эмбриоидов в строго контролируемых условиях in vitro [5], в том числе при культивировании in vitro изолированных пыльников пшеницы [6].
При анализе структурных механизмов становления полярности и симметрии эмбриоидов in vitro большую помощь оказывает метод сканирующей электронной микроскопии (СЭМ), позволяющий получить точное представление о пространственной организации изучаемого объекта и тонких деталях его строения, а также точно идентифицировать конкретный путь морфогенеза in vitro , реализующийся при культивировании изолированных пыльников.
Цель работы состояла в изучении микроспори-альных эмбриоидов пшеницы методом СЭМ и анализе структурных механизмов становления их симметрии.
Объектом исследования послужили микроспо-риальные эмбриоиды, полученные при культивировании in vitro пыльников яровой мягкой пшеницы сорта Жница.
Донорные растения выращивались в полевых условиях на экспериментальных участках научного стационара Института биологии УНЦ РАН (Уфимский р-н).
Микроспориальные эмбриоиды получали с использованием метода культуры in vitro изолированных пыльников яровой мягкой пшеницы [7]. Образцы для сканирующей электронной микроскопии готовили согласно[8], высушивали с помощью прибора «Critical point» (Hitachi, Japan), электронопроводящее покрытие создавали термическим напылением золота в вакууме с применением вакуумного универсального поста (ВУП). Объекты анализировали с помощью сканирующего электронного микроскопа JSM-6390 (Jeol, Japan; на базе БИН РАН, г. Санкт-Петербург).
В культуре in vitro изолированных пыльников при использовании низких концентраций 2,4-Д образуются эмбриоиды (рис. 1 а ), сходные по строению с зиготическим зародышем пшеницы (рис. 1 б ). Такие эмбриоиды характеризуются дорзовентраль-ной симметрией и наличием всех органов, присущих зиготическому зародышу. Разнообразные формы эмбриоидов можно объяснить их развитием в отсутствии жесткой тканевой детерминации, имеющей место у зиготического зародыша in vivo.
При повышении концентрации 2,4-Д образуются эмбриоиды, представляющие собой сложные структуры – полиэмбриоиды – с измененным типом полярности и симметрии, что выражается в образовании в их апикальной части множественных щитков и апикальных меристем побега (от 2 до 5, рис. 2), объединенных общим корневым полюсом (иногда отмечается дифференциация двух независимых или поверхностно сросшихся апикальных меристем первичного корня).
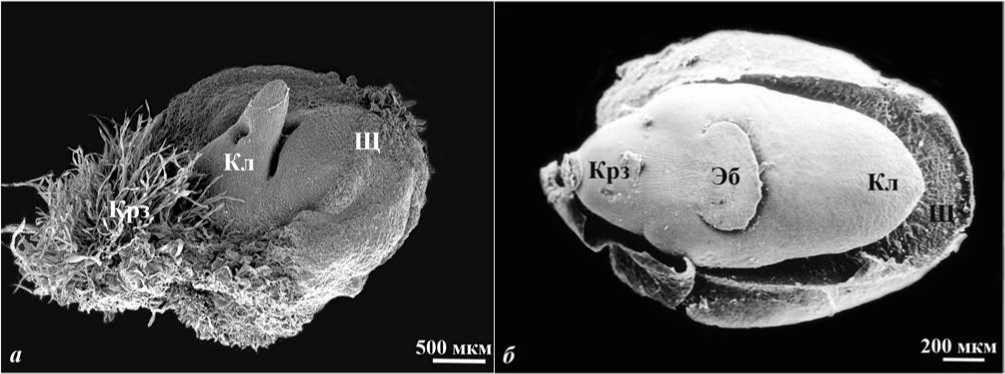
Рис. 1. Микроспориальный эмбриоид ( а ), сходный по строению с зиготическим зародышем ( б). СЭМ. Условные обозначения : Кл - колеоптиль, Крз - колеориза, Щ - щиток, Эб - эпибласт
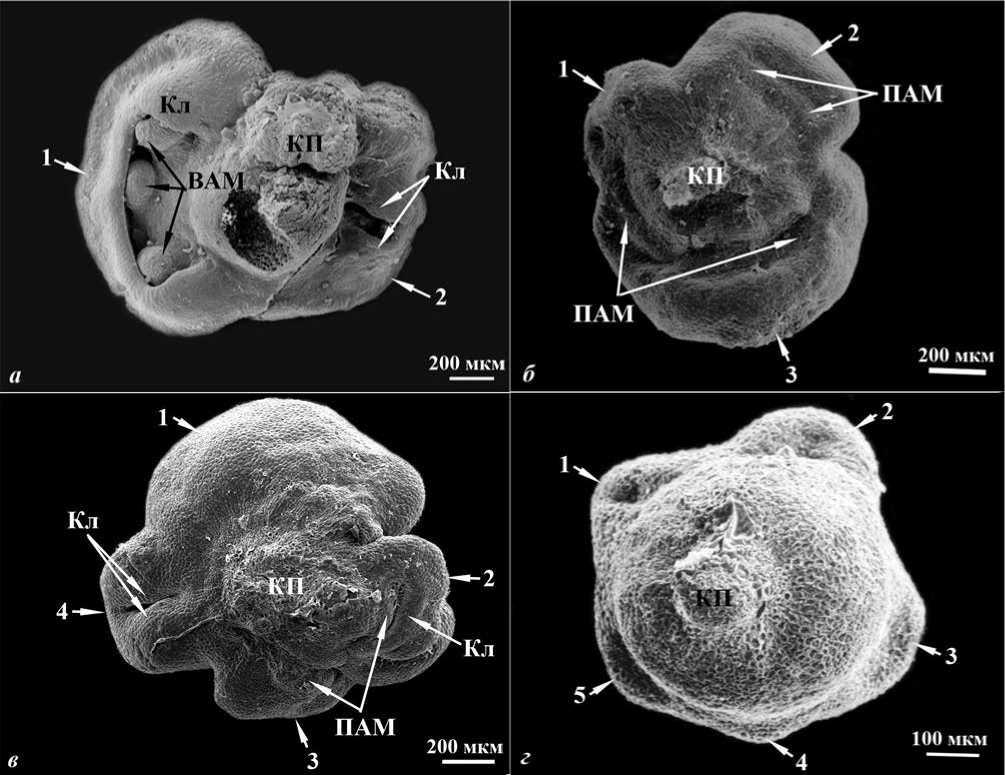
Рис. 2. Микроспориальные полиэмбриоиды с множественными щитками и апикальными меристемами побега: а - с двумя, б - тремя, в - четырьмя, г - пятью щитками, соответственно. СЭМ. Условные обозначения : ВАМ -вторичная апикальная меристема побега, Кл - колеоптиль, КП - корневой полюс, ПАМ - первичная апикальная меристема побега. Прим. Цифрами обозначены отдельные щитки
Анализ полученных результатов позволяет выделить два типа полиэмбриоидов с различным типом симметрии: 1) полиэмбриоиды с щитками, обращенными друг к другу дорзальной стороной и расположенными, в целом, терминально по отношению к соответствующим апикальным меристе мам побега (преобладающий тип) (рис. 2) и 2) полиэмбриоиды с щитками, обращенными друг к другу вентральной стороной и расположенными латерально по отношению к соответствующим апикальным меристемам побега (единичные случаи) (рис. 3).
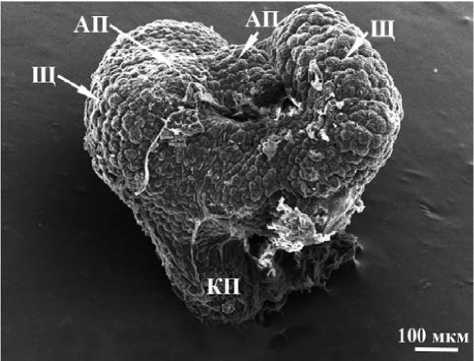
Рис. 3. Микроспориальный полиэмбриоид с щитками, обращенными друг к другу вентральной стороной. СЭМ. Условные обозначения : АП – апекс побега, КП – корневой полюс, Щ – щиток
Возможный механизм формирования таких полиэмбриоидов – кливажная полиэмбриония, состоящая в возникновении нескольких зародышей из зиготы или зародышей различного происхождения вследствие их расщепления [9].
При определенных условиях клетки зародышей и эмбриоидов могут освобождаться от организменного контроля, становиться тотипотентными, а затем детерминированными к формированию «нового» зародыша.
У голосеменных растений кливажная полиэмбриония – явление спонтанное и регулярное как в условиях in vivo , так и in vitro [11].
У покрытосеменных растений процесс кливажной полиэмбриоинии in vivo происходит довольно редко [9], но может быть индуцирован с использованием различных обработок, как правило, применением синтетических ауксинов [12–15] или ингибиторов полярного транспорта ауксинов [16] на ранних стадях развития зародышей.
Оба типа полиэмбриоидов, полученных нами в культуре in vitro пыльников пшеницы, в целом идентичны фенотипам, индуцированных воздействием in vitro ингибиторов полярного транспорта ауксинов (N-1-нафтилфталамовая кислота, кверти-цин) на изолированные зиготические зародыши Triticum aestivum L. в глобулярной стадии и раннего перехода к органогенезу [16].
В проанализированных нами литературных источниках [12-16] не прослежен генезис таких полимерных структур. Изучение генезиса полиэмбриоидов 1-го типа с использованием метода СЭМ позволило выявить структурный механизм кливажной полиэмбрионии лежащий в основе их образования.
Такой механизм заключается в следующем: за счет увеличения размеров апикальной части первичного эмбриоида (по крайней мере, с глобулярной стадии развития) на ней вместо одной возникают множественные точки роста, часто неравноценные по объему, что приводит к различиям в размерах образующихся множественных щитков и соответствующих им апикальных меристем побега (рис. 2 а – в ).
Аномальное увеличение размеров первичных апикальных меристем побега (рис. 2 б , в ) приводит к возникновению множественных вторичных апикальных меристем (рис. 2 а ), образующих в дальнейшем систему фасциированных побегов.
Известно, что главную роль в становлении симметрии зародыша растений играет негомогенное распределение ауксина (градиенты ауксина); потоки ауксина, создавая позиционную информацию, действуют как мощнейший морфогенетический фактор и определяют дифференциацию органов зародыша [2, 16].
Анализ полученных результатов позволяет сделать вывод об общности механизма гормональной регуляции образования полимерных структур на основе процесса кливажной полиэмбрионии как зиготических зародышей [16], так и микроспориаль-ных эмбриоидов пшеницы. Данный механизм состоит в нарушении полярного транспорта ауксинов в зародыше/эмбриоиде на глобулярной стадии развития, приводящей к изменению их симметрии и полярности: в зародыше – под действием ингибиторов полярного транспорта ауксинов, в эмбриоидах – вследствие создания в них избытка ауксинов при повышении концентрации 2,4-Д в индукционной среде.
C.Fischer и G.Neuhaus [17] установили, что в недифференцированном зиготическом зародыше пшеницы вокруг апикальной части существует кольцо клеток, компетентных к формированию меристемы побега. В условиях in vivo ауксин из базальной части зародыша транспортируется полярно в двух направлениях – к месту дифференциации апекса побега и к месту дифференциации щитка – и недоступен для других групп клеток зародыша. Однако при нарушении транспорта ауксин может накапливаться в клетках, в которых обычно его содержание низко. Авторы предположили, что эти клетки могут быть уже морфогенетически компетентными к дифференциации органов, либо же им для этого требуется импульс в виде потока ауксина. В результате формируются дополнительные меристемы побега и формируются полиэмбрионы.
По мнению B.Haccius [13], недифференцированный зародыш уже обладает «апикальным доминированием». Возможно, что при действии ростовыми веществами повреждается группа клеток, из которой позднее возникнет апекс побега и которая подавляет формирование других апексов. При этом действие клеток будущего апекса прекращается, а клетки, которые в норме должны давать начало семядолям, дают начало новым апексам побегав.
Дальнейшая детализация данных по генезису микроспориальных полиэмбриоидов позволит внести вклад в понимание процессов морфогенеза зародыша и кливажной полиэмбрионии, а также внести уточнения в гипотезу о механизме гормональной регуляции становления полярности и симметрии в эмбриогенезе злаков и подойти к управлению этими процессами в условиях in vitro .
-
1. Бутенко Р.Г. Биология клеток высших растений in vitro и биотехнологии на их основе. М.: ФБК-ПРЕСС, 1999. 160 с.
-
2. Медведев С.С. Механизмы формирования и физиологическая роль полярности в растениях // Физиол. раст. 2012. Т. 59. № 4. С. 543-556.
-
3. Theißen G ., Saedler H. Molecular architects of plant body plans // Progress in botany. V. 59 / eds H.-D.Behnke, K. Esser, J.W. Kadereit, U. Lüttge, M. Runge. Berlin, Heidelberg: Springer-Verlag, 1998. P. 227-256.
-
4. Raghavan V . Embryogenesis in angiosperms: a developmental and experimental study. New York: Cambridge Univer. Press, 1986. 316 p.
-
5. Zimmerman J.L . Somatic embryogenesis: a model for early development in higher plants // Plant Cell. 1993. V. 5. N 10. P. 1411-1423.
-
6. Круглова Н.Н . Микроспора злаков как модельная система для изучения путей морфогенеза: Автореф. дис. … д-ра биол. наук. СПб., 2002. 48 с.
-
7. Круглова Н.Н., Батыгина Т.Б. Методические рекомендации по использованию морфогенетического потен-
циала пыльника в биотехнологических исследованиях яровой мягкой пшеницы. Уфа, 2002. 39 с.
-
8. Миронов A.A., Комиссарчик Я.Ю., Миронов В.А . Методы электронной микроскопии в биологии и медицине. СПб.: Наука, 1994. 399 с.
-
9. Батыгина Т.Б. Воспроизведение, размножение и возобновление растений // Эмбриология цветковых растений. Терминология и концепции. Т. 3: Системы репродукции / Под ред. Т.Б. Батыгиной. СПб.: Мир и семья, 2000. С. 35-39.
-
10. Sharp W.R., Sondahl M.R., Caldas L.S., Maraffa S.B. The physiology of in vitro asexual embryogenesis // Hort. Rev. V. 2 / eds F.G. Dennis, D.N. Maynard, M.N. Rogers. Westport, Connecticut: The AVI Publishing company, 1980. P. 268-310.
-
11. Gupta P.K., Durzan D.J. Somatic polyembryogenesis from callus of mature sugar pine embryos // Nat. Biotechnol. 1986. V. 4. N 7. P. 643-645.
-
12. Яковлев М.С., Снегирев Д.П. Влияние ростовых веществ на образование многозародышевых зерновок у пшеницы // Ботан. журн. 1954. Т. 39. № 2. C. 187-194.
-
13. Haccius B. Experimentally induced twinning in plants // Nature. 1955. V. 176. N 4477. P. 355-356.
-
14. Ferguson J.D., Mc Ewan J.M., Card K.A. Hormonally induced polyembryos in wheat // Physiol. Plant. 1979. V. 45. N 4. P. 470-474.
-
15. Erdelska O., Vidovencova Z. Cleavage polyembryony in vivo and in vitro // Biol. Plant. 1994. V. 36. N 3. P. 329-334.
-
16. Fischer C ., Speth V., Fleig-Eberenz S., Neuhaus G . Induc- tion of zygotic polyembryos in wheat: influence of auxin po
lar transport // The Plant Cell. 1997. V. 9. N 10. P. 17671780.
-
17. Fischer C., Neuhaus G. Influence of auxin on the establishment of bilateral symmetry in monocots // Plant J. 1996. V. 9. N 5. P. 659-669.
STRUCTURAL MECHANISMS OF SYMMETRY ESTABLISHMENT IN MICROSPORE DERIVED EMBRYOIDS OF WHEAT: THE DATA OF SCANNING ELECTRON MICROSCOPY
Список литературы Структурные механизмы становления симметрии у микроспориальных эмбриоидов пшеницы: данные сканирующей электронной микроскопии
- Бутенко Р.Г. Биология клеток высших растений in vitro и биотехнологии на их основе. М.: ФБК-ПРЕСС, 1999. 160 с.
- Медведев С.С. Механизмы формирования и физиологическая роль полярности в растениях//Физиол. раст. 2012. Т. 59. № 4. С. 543-556.
- Theißen G., Saedler H. Molecular architects of plant body plans//Progress in botany. V. 59/eds H.-D.Behnke, K. Esser, J.W. Kadereit, U. Lüttge, M. Runge. Berlin, Heidelberg: Springer-Verlag, 1998. P. 227-256.
- Raghavan V. Embryogenesis in angiosperms: a developmental and experimental study. New York: Cambridge Univer. Press, 1986. 316 p.
- Zimmerman J.L. Somatic embryogenesis: a model for early development in higher plants//Plant Cell. 1993. V. 5. N 10. P. 1411-1423.
- Круглова Н.Н. Микроспора злаков как модельная система для изучения путей морфогенеза: Автореф. дис. … дра биол. наук. СПб., 2002. 48 с.
- Круглова Н.Н., Батыгина Т.Б. Методические рекомендации по использованию морфогенетического потенциала пыльника в биотехнологических исследованиях яровой мягкой пшеницы. Уфа, 2002. 39 с.
- Миронов A.A., Комиссарчик Я.Ю., Миронов В.А. Методы электронной микроскопии в биологии и медицине. СПб.: Наука, 1994. 399 с.
- Батыгина Т.Б. Воспроизведение, размножение и возобновление растений//Эмбриология цветковых растений. Терминология и концепции. Т. 3: Системы репродукции/Под ред. Т.Б. Батыгиной. СПб.: Мир и семья, 2000. С. 35-39.
- Sharp W.R., Sondahl M.R., Caldas L.S., Maraffa S.B. The physiology of in vitro asexual embryogenesis//Hort. Rev. V. 2/eds F.G. Dennis, D.N. Maynard, M.N. Rogers. Westport, Connecticut: The AVI Publishing company, 1980. P. 268-310.
- Gupta P.K., Durzan D.J. Somatic polyembryogenesis from callus of mature sugar pine embryos//Nat. Biotechnol. 1986. V. 4. N 7. P. 643-645.
- Яковлев М.С., Снегирев Д.П. Влияние ростовых веществ на образование многозародышевых зерновок у пшеницы//Ботан. журн. 1954. Т. 39. № 2. C. 187-194.
- Haccius B. Experimentally induced twinning in plants//Nature. 1955. V. 176. N 4477. P. 355-356.
- Ferguson J.D., Mc Ewan J.M., Card K.A. Hormonally induced polyembryos in wheat//Physiol. Plant. 1979. V. 45. N 4. P. 470-474.
- Erdelska O., Vidovencova Z. Cleavage polyembryony in vivo and in vitro//Biol. Plant. 1994. V. 36. N 3. P. 329-334.
- Fischer C., Speth V., Fleig-Eberenz S., Neuhaus G. Induction of zygotic polyembryos in wheat: influence of auxin polar transport//The Plant Cell. 1997. V. 9. N 10. P. 1767-1780.
- Fischer C., Neuhaus G. Influence of auxin on the establishment of bilateral symmetry in monocots//Plant J. 1996. V. 9. N 5. P. 659-669.
