Структурные преобразования среднего мозга крыс при экспериментальном моделировании дефицита магния

Автор: Смирнов А.В., Евсюков О.Ю., Спасов А.А., Харитонова М.В., Желтова А.А.
Журнал: Волгоградский научно-медицинский журнал @bulletin-volgmed
Рубрика: Морфология
Статья в выпуске: 1 (33), 2012 года.
Бесплатный доступ
В работе представлены результаты качественного и количественного анализа структурных изменений различных отделов среднего мозга крыс при экспериментальном моделировании алиментарного дефицита магния. Отмечены различия в характере и выраженности структурных преобразований нейронов и нейропиля различных отделов среднего мозга крыс в условиях экспериментальной гипомагнезиемии и при пищевой коррекции последней магния L-аспарагинатом.
Средний мозг, магний, дефицит
Короткий адрес: https://sciup.org/142148942
IDR: 142148942 | УДК: 616.132.2-092.9:546
The structural changes of the midbrain of rats in experimental simulation of magnesium deficiency
We present the results qualitative and quantitative analysis of structural changes in various parts of the midbrain of rats with experimental modeling of nutritional magnesium deficiency. The main differences in the nature and severity of structural changes of neurons and neuropile of various departments of the midbrain of rats in experimental hypomagnesemia and its correction with magnesium L-asparaginate.
Текст научной статьи Структурные преобразования среднего мозга крыс при экспериментальном моделировании дефицита магния
В биологических системах магний является одним из важнейших макроэлементов, являющимся кофактором более трехсот ферментов, контролирующих, прежде всего, обмен аденозинтрифосфата, его участие в ионном транспорте, передаче внутриклеточных сигналов и осуществлении активного переноса молекул через биологические мембраны, в том числе и для создания трансмембранного электрического потенциала в возбудимых тканях [1, 2, 6, 7]. Магниевая недостаточность создает неблагоприятный преморбидный фон для большого круга форм патологии нервной системы (экстрапирамидных расстройств, депрессии, мигрени и других цефалгий, сосудистых и нейродегенеративных поражений) [1, 6]. Магний важен для регуляции обмена нейромедиаторов и модулирования функции рецепторов в центральной нервной системе, в том числе ответственных за формирование депрессивно-подобного и тревожного поведения у животных [5]. В соответствии с современными представлениями о нейрохимической организации головного мозга основная масса катехол- и индоламинергических нейронов локализуется в различных отделах среднего мозга. Алиментарная коррекция магниевой недостаточности L-аспарагинатом магния способствует нормализации холинергической, норадренергической, серотонинергической нейротрансмиссии в центральной нервной системе, повышает порог коразоловых судорог [3]. Учитывая важную роль магния в патологии нейромедиаторного обмена представляется целесообразным оценить структурные изменения среднего мозга экспериментальных животных в условиях алиментарной магниевой недостаточности, а также при ее коррекции магний L-аспарагинатом.
ЦЕЛЬ РАБОТЫ
Определение влияния алиментарной недостаточности магния и ее коррекции на структурные изменения красных ядер среднего мозга крыс.
МЕТОДИКА ИССЛЕДОВАНИЯ
Исследования были выполнены на 30 половозрелых нелинейных белых крысах-самцах массой 220—240 г. В ходе эксперимента выделены группы животных: первая — контрольная интактная (10 животных), вторая — с моделированием алиментарного дефицита магния в течение 8 недель (10 животных), третья — животные, находившиеся на магний-дефицитной диете в течение 8 недель с последующим применением магния L-аспарагината в таблетированной форме в течение 4 недель (10 животных). Животные содержались в условиях вивария, согласно правилам лабораторной практики при проведении доклинических исследований в РФ (ГОСТ Р 50258-92, ГОСТ З 51000.3-96 и 51000.4-96) с соблюдением «Международных рекомендаций Европейской конвенции по защите позвоночных животных, используемых при экспериментальных исследованиях», (1997). Эксперименты были одобрены Комитетом по этической экспертизе исследований Волгоградского государственного медицинского университета (протокол № 35-2006 от 27.03.2006). Интактные животные (1-я группа) получали полноценную магний-сбалансированную диету, содержащую 0,84 г MgO (0,5 г элементарного магния) на 1 кг диеты и отстоянную воду (содержание магния 20 мг/л). Животные, у которых моделировали гипомагнезиемию, получали специальную магний-дефицитную диету фирмы «ICN Biomedicals Inc.» (Aurora, Ohio, США), которая включала 20 % казеина, 70 % крахмала, 0,3 % DL-метионина, 0,2 % холина битартрата, 5 % кукурузного масла, 1 % поливитаминной смеси, 3,5 % диеты составляла полиминеральная смесь AIN-76, не содержащая магния. Для питья использовалась дистиллированная вода. Скорость и глубину развития гипомагнезиемии контролировали, определяя концентрацию магния в плазме и эритроцитах крови спектрофотометрическим методом по цветной реакции с титановым желтым (Sigma, США) с измерением на спектрофотометре «СФ-26» (ЛОМО, Россия) в кювете с длиной оптического пути 1 см при длине волны 550 нм. При снижении концентрации магния ниже 1,4 ммоль/л в эритроцитах и ниже 0,7 ммоль/л в плазме считалось, что у животных развилась гипомагнезиемия средней тяжести, свидетельствующая об адекватности используемой экспериментальной модели. Животным, находящимся на магний-дефицитной диете, в течение 8 недель перорально через зонд вводили исследуемую соль магния. Доза магния L-аспарагината рассчитывалась исходя из 50 мг элементарного магния на 1 кг веса животного. Усредненное время компенсации дефицита магния составляло около 4 недель. Забой животных проводился согласно требованиям, изложенным в «Международных рекомендациях по проведению медико-биологических исследований с использованием животных» (1989). Образцы головного мозга фиксировались в течение 24 часов в нейтральном забуференном 10%-м формалине с дальнейшим обезвоживанием в батарее спиртов и изготовлением парафиновых срезов толщиной 4—6 мкм, которые окрашивали по стандартным методикам гематоксилином и эозином, тионином по Нисслю. При морфологическом исследовании оценивали такие морфометрические параметры, как средняя площадь перикарионов нейронов, средняя площадь ядра нейрона, средняя площадь цитоплазмы перикарионов нейронов, ядерно-цитоплазматическое отношение, удельное количество нейронов с «темной» цитоплазмой. Полученные данные обрабатывались методами базисного статистического анализа в программе STATISTICA/6.0 фирмы StatSoft Inc. (США) для Windows.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ И ИХ ОБСУЖДЕНИЕ
В группе животных с дефицитом магния (в течение 8 недель) наблюдается уменьшение массы тела животных по сравнению с контролем на 18 % (p < 0,05), уменьшение массы головного мозга по сравнению с контролем — на 4 % (p < 0,05), уменьшение относительной массы головного мозга на 2 % (p < 0,05). При микроскопическом исследовании различных отделов среднего мозга наиболее выраженные изменения отмечаются в красных ядрах, центральном сером веществе, черной субстанции, в верхнем центральном и дорсальном ядрах шва. Наибольший интерес представляют изменения со стороны красных ядер, обеспечивающих интеграцию эфферентных сигналов экстрапирамидной системы [8], а также ядер шва, составляющие морфологическую основу серотонин-опосредованных депрессивно-подобных и тревожных поведенческих расстройств у изучаемых животных [1]. Отмеченные регионы среднего мозга представлены преимущественно крупными мультиполярными нейронами с треугольной и веретеновидной формой перикарионов, также встречаются мелкие и средние нейроны с округлой и полигональной формой перикарионов. Ядра многих крупных и мелких нейронов при дефиците магния сморщены, резко ба-зофильны, с мозаичной конденсацией гетерохроматина. В кариоплазме отдельных нейронов становятся трудно различимыми эктопированные ядрышки, отмечается появление глыбчатых пик-нотически измененных гомогенных ядерных фрагментов, напоминающих апоптозные тельца, отме- чается очаговая вакуолизация ядер и цитоплазмы перикарионов нейронов. В большинстве нейронов обнаруживается мозаичный гиперхроматоз цитоплазмы, в некоторых — просветление перинуклеарной области цитоплазмы перикариона. В нейропиле ядер центрального серого вещества, ядер шва при дефиците магния выявлялись набухшие глиоциты, явления периваскулярной ней-рофагии. В указанных регионах выявлены выраженное полнокровие сосудов микроциркулятор-ного русла, стаз эритроцитов в капиллярах, отек и набухание эндотелиоцитов капилляров и мелких артериол, ядра эндотелиоцитов артериол иногда располагаются перпендикулярно базальной мембране, что является одним из признаков вазоспазма.
В красных ядрах среднего мозга магний-де-фицитных животных при морфометрическом исследовании наблюдается увеличение средней площади перикарионов нейронов — на 2,5 %, а при алиментарной коррекции магниевой недостаточности — на 1,62 % по сравнению с контролем (табл.). Отмечается увеличение средней площади цитоплазмы перикарионов нейронов у магний-дефицитных крыс — на 6,4 %, а при коррекции магния L-аспарагинатом — на 4,2 % по сравнению с контролем. Средняя площадь ядер нейронов при дефиците магния уменьшается — на 4,5 %, а при пищевой коррекции — на 2,6 % по сравнению с контролем.
В группе животных с дефицитом магния отмечается достоверное (при р < 0,05) увеличение удельного количества темных нейронов на 23,4 % по сравнению с контрольной группой (рис. 1).
Таким образом, достоверное увеличение количества темных нейронов красных ядер среднего мозга (рис. 2) в группе с моделируемым в течение 8 недель магниевым дефицитом может свидетельствовать как о роли изучаемого макроэлемента в регуляции функций экстрапирамидной системы, так и о магний-индуцированных механизмах повреждения высокоспециализированных клеток центральной нервной системы [6, 8]. При этом диссоциированные изменения ядер и цито-плазмы перикарионов нейронов среднего мозга могут отражать различные механизмы морфологических преобразований при нарушении магниевого гомеостаза.
Изменение некоторых морфометрических параметров нейронов красных ядер среднего мозга крыс при алиментарном дефиците магния ( М ± m ) и его коррекции
|
Показатель |
Группы |
||
|
контрольная |
дефицит магния (8 недель) |
солевая коррекция магниевой недостаточности (магния L-аспарагинат) |
|
|
Масса тела, г |
225,78 ± 11,41 |
185,16 ± 6,91* |
193,43 ± 7,88* |
|
Масса головного мозга, г |
1,90 ± 0,05 |
1,83 ± 0,04 |
1,86 ± 0,07 |
|
Средняя площадь перикарионов нейронов, мкм2 |
202,16 ± 6,20 |
207,15 ± 3,26 |
205,45 ± 4,14 |
|
Средняя площадь ядра нейрона, мкм2 |
67,12 ± 5,08 |
63,34 ± 4,11 |
64,61 ± 3,15 |
|
Средняя площадь цитоплазмы перикарионов нейронов, мкм2 |
135,04 ± 5,12 |
143,67 ± 4,39 |
140,80 ± 4,12 |
|
Ядерно-цитоплазматическое отношение, число |
0,49 ± 0,03 |
0,44 ± 0,02 |
0,46 ± 0,03 |
|
Удельное количество нейронов с «темной» цитоплазмой, % |
9,12 ± 1,02 |
12,78 ± 1,06* |
11,56 ± 1,84 |
* p < 0,05 при сравнении с контролем.

Рис. 1. Увеличение количества темных нейронов в красном ядре среднего мозга при дефиците магния.
Окр. тионином по Нисслю, ув. х 100
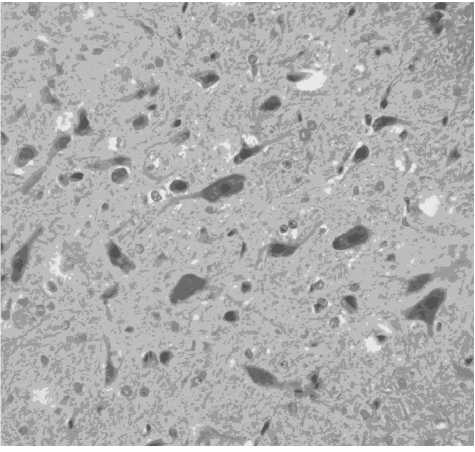
Рис. 2. Красное ядро среднего мозга при моделировании алиментарного дефицита магния. Окр. гематоксилином и эозином, ув. х 400
ЗАКЛЮЧЕНИЕ
Проведенное морфологическое исследование выявило ряд структурных изменений в нейронах и нейропиле различных отделов среднего мозга (красных ядрах, центральном сером веще- стве, черной субстанции, ядрах шва) крыс при экспериментальном моделировании алиментарного дефицита магния, что согласуется с литературными данными [8] о роли магниевой недостаточности в развитии экстрапирамидных расстройств, а также нейромедиаторного дисбаланса в центральной нервной системе [5]. Обнаруженные морфологические изменения в головном мозге крыс с алиментарной коррекцией дефицита магния L-аспарагинатом свидетельствуют о модулирующем влиянии этого вещества на состояние мик-роциркуляторного русла, нейронов и нейропиля ядер среднего мозга.
