Структурные типы местообитаний и стабильность пространственной организации индивидуальных территорий певчего и черного дроздов

Автор: Пискунов В.В., Давиденко Т.Н.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Наземные экосистемы
Статья в выпуске: 1-1 т.11, 2009 года.
Бесплатный доступ
В статье представлены данные по спектру используемых дроздами местообитаний в лесах южной части Приволжской возвышенности, проведено количественное описание структурных параметров местообитаний и индивидуальных территорий. Выделены структурные типы растительных сообществ, которые являются экологически наиболее подходящими для каждого вида, установлено постоянство структурной организации растительных сообществ в пределах индивидуальных территорий с абсолютным преобладанием одного-двух профильных типов фитоценозов. В пространственном расположении индивидуальных территорий наибольшая межгодовая стабильность их границ выявлена в пределах переходных типов местообитаний.
Типы местообитаний, индивидуальные территории, птицы
Короткий адрес: https://sciup.org/148198264
IDR: 148198264 | УДК: 598.288.5:591.5+581.524
Habitat structure types and stability of the spatial organization of song thrush and blackbird individual territories
In this article data on a spectrum of habitats used by Turdus philomelos and Turdus merula in woods of southern part of Privolzhskay sublimity are presented, the quantitative description of structural parameters of habitats and individual territories is lead. Structural types of vegetative communities which are ecologically the most suitable for each kind are allocated, the constancy of the structural organization of vegetative communities within the limits of individual territories with absolute prevalence of one-two phitocenoses profile types is established. In a spatial arrangement of individual territories the greatest interannual stability of territories borders is revealed within the limits of transitive biotopes.
Текст научной статьи Структурные типы местообитаний и стабильность пространственной организации индивидуальных территорий певчего и черного дроздов
Многие авторы сходятся во мнении, что критерием пригодности территории для гнездования многих видов птиц служат такие внешние признаки биотопа, которые указывают на обилие доступных ресурсов [3, 5, 10]. Например, для воробьиных птиц таким признаком может служить характер структурной организации растительных сообществ, который определяет целый комплекс условий — кормовых, защитных и микроклиматических. В зависимости от структурной сложности растительные сообщества обладают неодинаковой экологической емкостью, которая проявляется в способности обеспечивать животных всеми необходимыми ресурсами [1, 4]. Вариации в качественных и количественных характеристиках биотопов приводят к тому, что в разных местообитаниях наблюдаются отличия в плотности гнездования отдельных видов птиц. Для большинства видов отмечается четкая биотопическая приуроченность, однако в пределах выбранного биотопа возможна значительная вариация видовых и структурных компонентов местообитаний [6].
На территории лесных сообществ южной части Приволжской возвышенности проведено изучение закономерностей пространственного размещения и временной стабильности индивидуальных территорий певчего ( Turdus
Владимир Валерьевич Пискунов, доцент кафедры ботаники и экологии; Татьяна Николаевна Давиденко, доцент той же кафедры.
philomelos C.L. Brehm) и черного ( Turdus merula Linnaeus) дроздов с целью выявления наиболее благоприятных местообитаний и определения типа структурной организации растительных сообществ, который является наиболее предпочитаемым изучаемыми видами птиц для гнездования.
Опираясь на данные по толерантности видов к изменению структурной неоднородности растительных сообществ, мы проанализировали спектр используемых дроздами местообитаний и провели количественные описания структурных параметров в пределах их индивидуальных территорий. Это позволило определить разнообразие используемых тем или иным видом местообитаний и выделить среди множества структурных типов растительных сообществ те, которые являются экологически наиболее подходящими для каждого вида и определяют возможность достижения высокого уровня плотности гнездования.
МАТЕРИАЛ И МЕТОДЫ
В ходе исследования сравнивались плотность гнездования певчего и черного дроздов, стабильность контуров индивидуальных территорий и признаки местообитаний на различных учетных площадках в 20 га, где в течение 5 лет в гнездовой период проводилось ежегодное картирование индивидуальных территорий.
Для учета дроздов в различных растительных сообществах использовался метод картографирования [11]. Все встреченные особи регистрировались на карте посещений при помощи специальных символов, на основе полученных регистраций выделялись кластеры, каждый из которых соответствует одной паре [2, 7]. В дальнейшем путем наблюдения за перемещением самцов между точками вокальной активности очерчивался контур индивидуальной территории. Для каждого вида были составлены отдельные карты расположения индивидуальных территорий в пределах каждого изученного растительного сообщества. Гнездовая плотность получена через усреднение данных за 5 лет (2003-2007 гг.).
Определение стабильности пространственного размещения индивидуальных участков по годам проводилось путем кодирования каждого квадрата (25х25 м) учетных площадок по принципу «плюс-минус» по критерию попадания в него индивидуальной территории. В дальнейшем проводился расчет дисперсии, которая служила критерием стабильности или вариации границ индивидуальных территорий по годам.
Описание структурной организации растительных сообществ проводилось с использованием специализированных методик [8, 9]. При определении структурного типа местообитаний в пределах выделенных растительных сообществ закладывались трансекты средней протяженностью 1 км, на которых в 30 точках измерялось проективное покрытие листвы на 11 различных высотах.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Для проведения исследования был выбран участок леса, на котором отчетливо прослеживается изменение структурных особенностей местообитаний при продвижении от центра к периферии массива: происходит постепенное уменьшение доли участия древесных пород в сложении сообществ, увеличение густоты кустарникового яруса, происходит ксерофитизация травяного покрова.
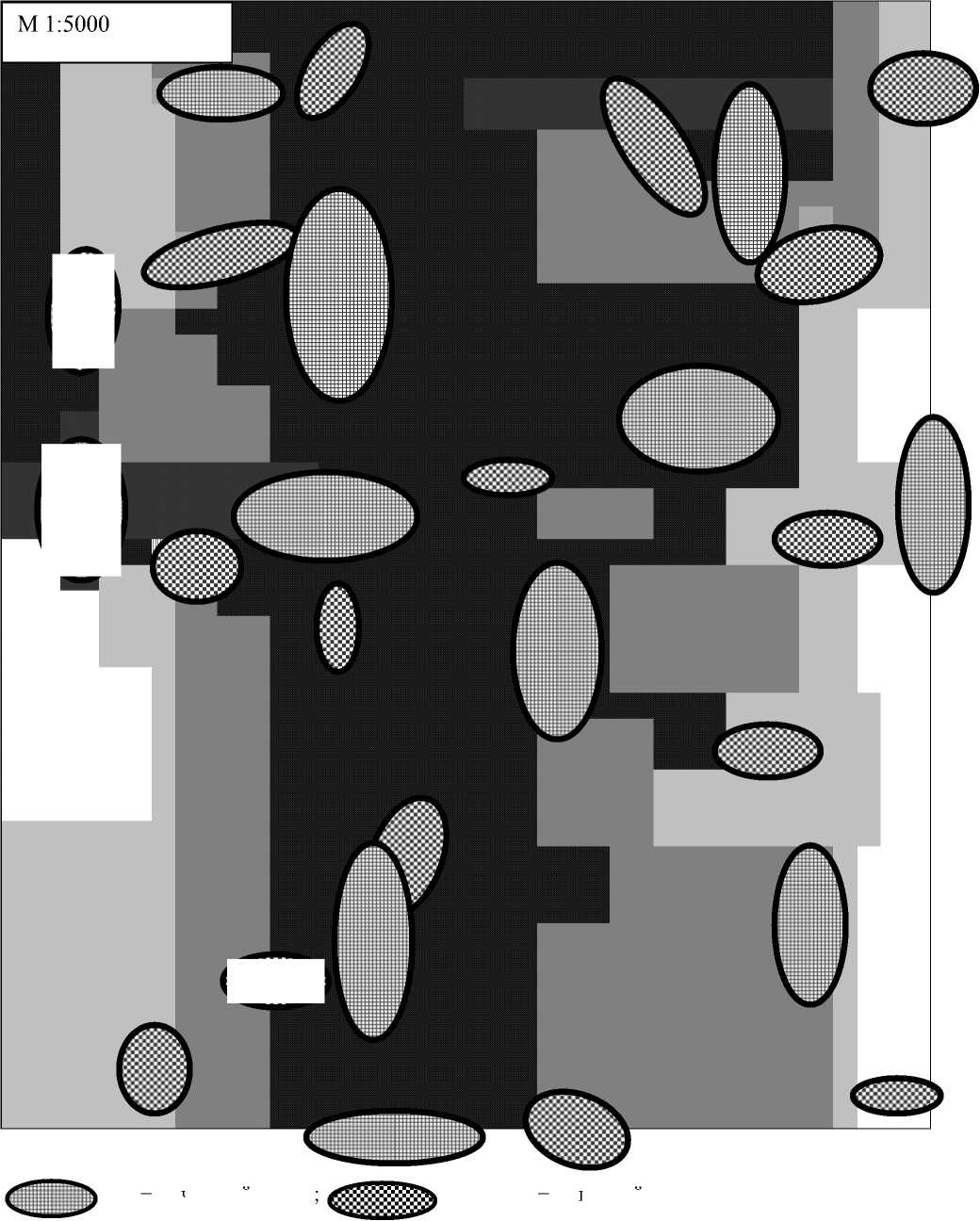
Исходя из особенностей структурной организации растительных сообществ, было выделено три основных типа местообитаний дроздов: 1) типичный лесной участок (густота древостоя 1000-1500 экз./га); 2) разреженный участок леса с хорошо развитым кустарниковым ярусом (густота древостоя 500-800 экз./га); 3) кустарниковые заросли на границе местообитаний лес-степь (густота древостоя 50-100 экз./га). Для первого типа местообитаний характерна очень высокая сте пень вертикальной структурированности при среднем уровне горизонтальной. Наибольшая горизонтальная неоднородность отмечается в пределах второго типа местообитаний дроздов, высока она также в кустарниковых зарослях за счет мозаичности подлеска. В соответствии с биотопическим подразделением было выделено три основных типа локализации индивидуальных территорий дроздов: 1) целиком в пределах лесного биотопа, 2) частично в пределах лесного типа местообитаний, частично в переходной кустарниковой зоне, 3) целиком в пределах кустарниковых зарослей (рис. 1).
Анализ распределения индивидуальных территорий изучаемых видов показал, что для певчего дрозда не наблюдается значимого изменения плотности гнездования при переходе от густого участка леса к разреженному, но резко снижается на границе со степью. Индивидуальные территории этого вида в переделах каждого типа местообитаний распределены равномерно, однако размеры территорий, расположенных на границе леса с кустарниковыми зарослями в среднем в 2,2 раза больше, чем прочих. Наименьшую суммарную площадь имеют территорий, целиком находящихся в пределах кустарникового биотопа.
Для черного дрозда характерна меньшая суммарная плотность гнездования, больший размер индивидуальных участков. Преобладающая часть территорий этого вида приурочена к типично лесному биотопу, и лишь некоторые территории расположены в пределах экотонной зоны лес-степь. При этом отмечена тенденция снижения размеров индивидуального участка в кустарниковых сообществах и увеличение ее в лесных биотопах при продвижении в центр лесного массива (табл.).
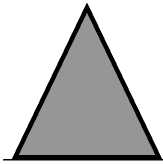
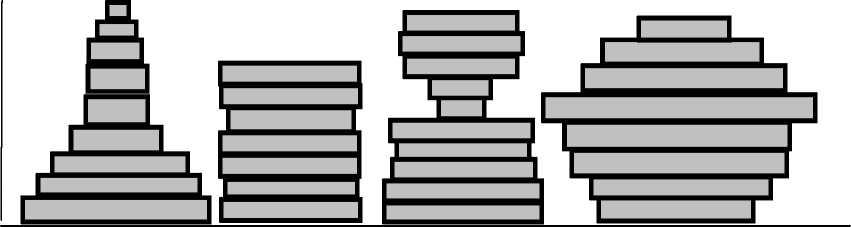
Несмотря на значительную разнородность растительных сообществ, в районе исследования было установлено, что в пределах индивидуальных территорий дроздов выделяется четыре основных структурных типа растительных сообществ (рис. 2).
Было установлено, что у изученных видов дроздов имеются предпочтения по характеру профильной организации растительных сообществ на индивидуальных участках: на территориях певчего дрозда наиболее часто встречается первый и второй тип распределения листвы по профилю сообщества, для черного дрозда более характерны третий и четвертый типы.
черный дрозд;
певчий дрозд
Рис. 1. Схема пространственного расположения индивидуальных территорий черного и певчего дроздов в различных типах биотопов: 1 — лесной биотоп, 2 — разреженный участок леса с хорошо развитым кустарниковым ярусом, 3 — кустарниковые заросли на границе местообитаний лес-степь, 4 — степь



Для певчего дрозда в границах индивиду- фитоценозов с ярко выраженной тенденцией альных территорий характерно преобладание убывания густоты ярусов и сомкнутости ли- ствы при продвижении снизу вверх по профилю. Так, на 70% территорий этого вида, несмотря на значительные отличия в абсолютных значениях, показатели густоты и сомкнутости крон нижних ярусов превышают таковые верхних ярусов. Для 20% территорий певчего дрозда отмечается практически равномерное распределение листвы по всему профилю сообщества (в основном это территории в пределах кустарниковых зарослей без четко выраженного древесного яруса).
Растительные сообщества в границах индивидуальных территорий черного дрозда, напротив, характеризуются наличием максимальных значений сомкнутости листвы не только в нижней, но и в верхней или средней частях профиля. Для большинства территорий данного вида (80%) типично наличие очень густого подлеска и травостоя, разреженного подроста и четко выраженного древесного яруса с высокими значениями сомкнутости крон (третий профильный тип). Другие 20% территорий характеризуются совершенно иной структурной организацией. Здесь отмечено наличие четко выраженных ярусов подроста, подлеска, древостой разреженный, наибольшая густота и сомкнутость характерна для подроста и высокого подлеска. За счет этих особенностей максимальные значения сомкнутости листвы достигаются в средней части структурного профиля (четвертый профильный тип).
Таблица
Обилие и особенности расположения гнездовых территорий певчего и черного дроздов
|
Местообитания |
Плотность гнездования, пар/100 га |
Доля территорий в пределах биотопа, % |
||
|
Черный дрозд |
Певчий дрозд |
Черный дрозд |
Певчий дрозд |
|
|
Типичный лесной участок |
18,6 |
22,7 |
64 |
23 |
|
Разреженный участок леса с развитым подлеском |
10,7 |
26,2 |
30 |
65 |
|
Кустарниковые заросли на границе местообитаний лес-степь |
5,4 |
8,3 |
6 |
12 |
|
Всего: |
34,7 |
57,2 |
100 |
100 |
А

2 3
Б
> 1
8 18,
0 12,
0 9,0 6,0 5,0 3,0 1,5 1,0
территории певчего дроз- территории черного дрозда
Рис. 2. Основные структурные типы растительных сообществ (по характеру распределения листвы снизу-вверх по профилю) в лесах южной части Приволжской возвышенности ( А ): 1 — «убывающий», 2 — «стабильный», 3 — «убывающе-возрастающий», 4 — «возрастающе-убывающий»; ( Б ) профильная организация сообществ, выявленная на индивидуальных территориях дроздов
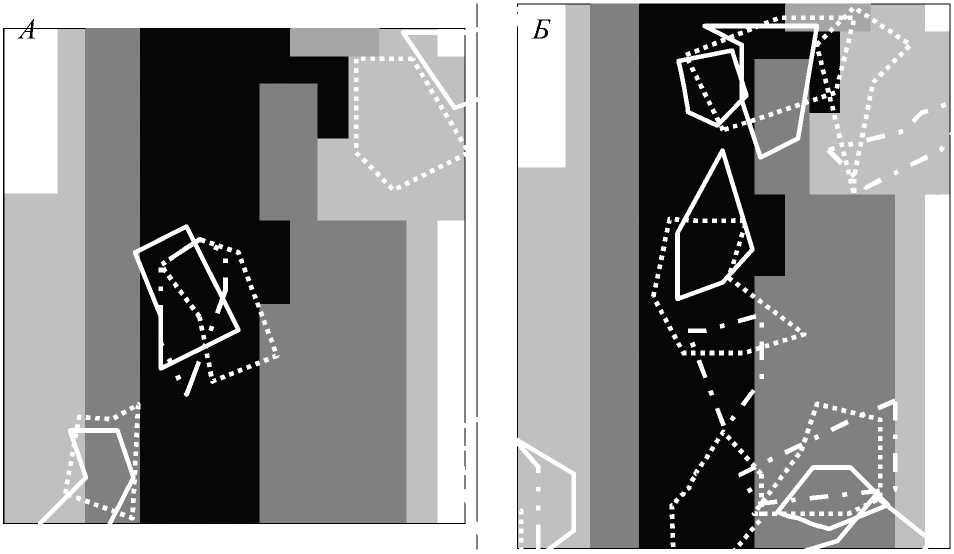
Для изучения стабильности пространственной организации индивидуальных территорий певчего и черного дроздов был проведен анализ распределения гнездовых территорий по годам. Он показал, что в целом для этих видов характерно значительное террито риальное постоянство расположения индивидуальных участков. Однако самый продолжительный срок занимания территории (с совпадением границ в разные годы) составляет не более 3 лет.
z М 1:5000
Рис. 3. Границы индивидуальных территорий певчего ( А ) и черного ( Л ) дроздов по юда.
2004; 2005; 2006
На уровне выделенных типов местообитаний из года в год сохранялось соотношение территорий, расположенных в пределах различных местообитаний. При этом для певчего дрозда наибольшее изменение пространственного расположения территорий отмечено в пределах лесного типа местообитаний. Напротив, на границе с кустарниковыми зарослями контуры индивидуальных территорий изменялись незначительно из года в год (межгодовое совпадение площади индивидуального участка более 50%), несмотря на изменение мест расположения гнезд (рис. 3). Для черного дрозда, большинство территорий которого располагались в лесном биотопе, отмечено значительное непостоянство границ индивидуальных территорий именно в этом типе местообитаний. Установлено, что за весь исследованный период только три территории остались более или менее постоянными из года в год. В одном случае отмечено совпадение только точки основной вокальной активности самца при полном изменении пространственного размещения участка. На границе местообитаний лес-степь, напротив, за весь период локализация индивидуальных территорий была относительно постоянной.
При сравнении особенностей расположения гнездовых территорий в период вторых кладок для певчего дрозда особых изменений выявлено не было. Как и в случае первой кладки, большинство территорий были расположены на границе леса с кустарниковыми зарослями. Для черного дрозда, напротив, при вторых кладках изменялись не только расположение гнезд, но и предпочитаемый тип местообитания. Так, в 40% случаев в конце июня — июле территории этого вида были локализованы не в лесном биотопе, а в обоих пограничных местообитаниях. Характерно также преобладание на этих территориях растительных сообществ, структурированных по четвертому профильному типу, который менее характерен для индивидуальных территорий в начале сезона размножения.
ЗАКЛЮЧЕНИЕ
Таким образом, для певчего и черного дроздов на изученной территории характерно использование для гнездования трех основных биотопов, которые значительно отличаются по структурным характеристикам растительных сообществ. Для певчего дрозда пограничные местообитания являются более подходящими, чем для черного, поскольку 77% всех его индивидуальных территорий располагалось именно в экотонной зоне. Черный дрозд предпочитает типично лесные биотопы. И для певчего, и для черного дроздов установлено постоянство структурной организации растительных сообществ в пределах индивидуальных территорий с абсолютным преобладанием одного-двух профильных типов фитоценозов. В пространственном расположении индивидуальных территорий выявлено определенное постоянство: наибольшей стабильностью и у черного, и у певчего дроздов отличаются территории в пределах переходных зон между лесом и кустарниковыми зарослями и между кустарниковыми зарослями и степью.
Список литературы Структурные типы местообитаний и стабильность пространственной организации индивидуальных территорий певчего и черного дроздов
- Владышевский Д.В. Оценка качества местообитаний лесных птиц//Экологическая оценка местообитаний лесных животных. Новосибирск: Наука, 1987.
- Гудина А.Н. Методы учета гнездящихся птиц: Картирование территорий. Запорожье: Дикое поле, 1999.
- Кулешова Л.В. Анализ структуры птичьего населения в связи с ярусностью леса//Орнитология. М., 1968. Вып. 9.
- Пискунов В.В., Давиденко Т.Н. Изменение экологической емкости лесных местообитаний под воздействием рекреации//Изв. Сарат. ун-та. 2007. Т. 7. Сер. Химия. Биология. Экология. Вып. 1.
- Преображенская Е.С. Экология воробьиных птиц Приветлужья. М.: КМК Scientific Press Ltd., 1998.
- Черенков С.Е., Губенко И.Ю. Факторы, определяющие пространственную структуру поселения дроздов в условиях высокой мозаичности леса//Зоол. журн. 1995. Т. 74, № 2.
- Bibbу C.J., Burgess N.Q., Hill D.A. Birds census technigues. London: Academic press, 1992.
- Blondel J., Сurvillier R. Une methode simple et rapide pour decrier les habitats d'oiseaux: le stratiscope//Oikos. 1977. № 29.
- Erdelen M. Birds communities and vegetation structure: Correlation and comparison of simple and diversity indices//Oecologia. 1984. V. 61.
- Pearson D.L. The relation of foliage complexity to ecological diversity of three Amazonian bird community//Condor. 1975. V. 77.
- Tomialojc L. The combined version of the mapping method//Bird census work and nature conservation. Guttingen: Dachverband Deutscher Avifaunisten. 1980.
