Study of chlorophyll a and b in etiolated and androgenic plants of barley

Author: Asakaviciute R., Jacquard С., Clement С.
Journal: Журнал стресс-физиологии и биохимии @jspb
Section: Original article
Article in issue: 1 т.2, 2006.
Free access
In order to characterize the occurrence of albino plantlets during the androgenetic process in barley (Hordeum vulgare L.), chlorophyllous content of microspore derived structures were analyzed in various cultivar. The dynamics of quantities of a and b chlorophylls and their relations within etiolated and androgenic plants of the contrasting cultivars of Igri and Cork have been compared. Research on chlorophyll content dynamics showed that the androgenic plants of cv. Igri accumulated a higher content of chlorophyll compared to the regenerants of cv. Cork. This deficiency of chlorophyll in regenerants may be directly related to a high number of albinos in the anther culture of cv. Cork
Albinism, barley, etiolated and androgenic plants, dynamics, chlorophylls
Short address: https://sciup.org/14323954
IDR: 14323954
Text of the review article Study of chlorophyll a and b in etiolated and androgenic plants of barley
Tel.: (+370) 687 22926, fax: (+370) 5 2645430, e-mail: , web: t
Tel.: (+370) 687 22926, fax: (+370) 5 2645430, e-mail: , web: t
One goal of recent plant biotechnology is to obtain doubled haploid plants. Doubled haploids can be used by plant breeders to produce homozygous plants and to develop agronomically improved varieties. Barley ( Hordeum vulgare L.) is one of the most commonly used species for doubled haploid production, with haploids mostly derived from androgenesis following microspore or anther culture (Caredda et al. 2000; Jacquard et al. 2003). However, the androgenic process induces formation of albino plantlets in varying proportions, depending on the cultivar. This phenommenon occurs most commonly in Poaceae (Caredda and Clement 1999) and greatly limits the potential yield of androgenesis in the most sensitive species such as barley (Jahne and Lorz 1995). Actually, several cultivars used in breeding programmes because of their agronomic importance can not be used following androgenesis because they produce exlusively albino plantlets (Andersen et al. 1987).
The process of androgenesis in barley has three stages: 1) induction – the usual development of gametophyte is blocked and an alternative sporophyte programme is induced; 2) cultivation – the microspores produce callus or embryoid structures; 3) regeneration – haploid plants are regenerated from androgenic embryoids or callus.
A great obstacle in barley anther culture is a distinct manifestation of albinism. The chloroplasts of microspores lose their inner membrane, are filled with lipids and globulins, and chlorophyll a is not synthesized from protochlorophyllid a (Mannienen 1997). The DNA of microspore chloroplasts is damaged at the early stage of microspores development. The efficiency of the anther culture method is largely dependent on the plant genotype and on cultivation conditions (Cistue et al. 1995; Sarrafi and Ghaemi 1997; Jacquard et al. 2003). Andersen has found that the genetic nature of donor plant affects the formation of embryoids by 20-40% and formation of green regenerates by 50-80% in the wheat anther culture (Andersen et al. 1987).
In some barley cultivars, green and albino plantlets are produced by an intermediate callus phase (Dunwell et al. 1987) or by direct embryogenesis (Jacquard et al. 2003). Using indirect embryogenesis, it has been shown that pt-genome deletions/rearrangements occur during early regeneration causing the regeneration of albino plantlets (Sunderland and Huang 1985; Leon et al. 1998). The transcripts from the pt-genes rbc L, psb D-psb C, as well as the 16S and 23S rRNAs are present at reduced levels in albino plantlets when compared to those found in green plantlets. Actually, transcripts of nuclear genes rbc S and Cab, which respectively encoded the small subunit of Rubisco and chlorophyll a linked proteins are abnormally low in albino plants (Dunford and Walden 1991). Using direct embryogenesis no information is avaible, particularly in microspore derived embryos at the biochemical level.
In order to further characterize the albino phenotype following anther culture, we examined the dynamics of chlorophylls a and b in etiolated and androgenic plants.
MATERIALS AND METHODS
The study was carried out at the Laboratory of Plant Biology and Physiology of Champagne Ardenne University in Reims, France in 2002. The experimental material: spring barley Cork was used as a genotype producing albino plants in anther culture at a high percentage, and winter barley Igri producing green regenerants at a high percentage (Wojnarowiez et al. 2002). The androgenic plants were obtained according to Caredda’s anther culture method (Caredda et al. 2000). The etiolated plants were grown at 20±2°C temperature in the dark for 8 days (Kouril et al. 1999). The test samples were taken after 8 days etiolation.
Plant material , the barley (Hordeum vulgare L.) winter cultivar ‘Igri’ and the spring cultivar ‘Cork’ were germinated and grown in the greenhouse at 25oC for a week. For the cv. Igri, seedlings were then vernalized for eight weeks at 4oC with 16 h photoperiod and grown in a controlled environment cabinet at 12oC with the same photoperiod in Florimond Desprez Ets.
Anther culture, spikes were collected at the vacuolated microspore stage. Anthers were dissected and pretreated at 4oC in the dark during 3-4 days in 3% (w/v) mannitol.
After pretreatment, anthers were cultured on the Caredda (1999) medium composed of microelemnts including KNO3 (1.9 g l-1), NH4NO3 (0.166 g l-1), KH 2 PO 4 (0.170 g l-1), CaCl 2 · 2H 2 O (0.029 g l-1), MgSO4 · 7H2O (0.374 g l-1); microelements including KI (0,830 mg l-1), MnSO 4 · H 2 O (16,3 mg l-1), Na 2 MoO 4 · 2H 2 O (0,250 mg l-1), H 3 BO 3 (6,2 mg l-1), ZnSO 4 · 7H 2 O (10,6 mg l-1), CuSO 4 · 5H 2 O (0,025 mg l-1), Fe-Na-EDTA (40 mg l-1); and aminoacids including glutamine (752 mg l-1). This medium was supplemented with 32 g l-1 mannitol as an osmoticum and 60 g l-1 of maltose as a carbohydrate source. The pH was adjusted to 5.6 before the addition of agarose (6 g l-1). After autoclaving, filter-sterilized vitamins, including myo-inositol (0.1 mg l-1), thiamine-HCl (0.4 mg l-1), and growth substances, including AIA (2 mg l-1) and BAP (1 mg l-1) were added to the medium.
Thirty anthers were placed per 5 cm Petri dishes. Dishes were sealed with a double layer of Parafilm and maintained at 26oC, with 85% relative humidity, in the dark for 21 days. After 21 days of culture, embryo-like structures (ELS) larger than 1 mm were transferred to the regeneration medium according to Caredda (1999) containing 3% (w/v) sucrose as a unique carbohydrate source and AIA/BAP (0.4/0.4 mg l-1) as phytohormones. The petri dishes were maintained at 26oC, 85% relative humidity, and 60 µE m-2 s-1 for 14 days with a 16/8 photoperiod. Albino and green androgenic plantlets were counted after 2 weeks on the regeration medium.
Preparation of Pigments, the content of chlorophylls a and b was extracted with acetone and tested in (1) androgenic and (2) etiolated plants (Lichtenthaler 1987). The content of pigments in the etiolated plants was tested every 24 hours up to the 10th day. In the androgenic plants (green (1) and albino (2) regenerants) the content of pigments was measured after 18 days regeneration, taking samples every two days up to 30th day of regeneration (3 replicates). The measurements were carried out by a spectrophotometer ‘Novaspec® II’ at 663 nm, 647 nm and 470 nm of the wavelength.
The content of chlorophylls was calculated:
The concentration of chlorophyll a (mg l-1):
C a = 12.25 x D 663 nm – 2.79 x D 647 nm
The concentration of chlorophyll b (mg l-1):
C b = 21.5 x D 647 nm – 5.1 x D 663 nm
The concentration of chlorophyll a and b (µg g-1): C a + C b
The ratio of chlorophyll a to chlorophyll b : C a / C b
Statistical data analysis, theoretical lines of regression give the results of chlorophylls. The regression determines how much on average the value of X changes when changing the value of Y, and vice versa. Linear regression equations y (x) = a + bx and determination factors (R2) have been calculated using the software “STAT”.
RESULTS
Changes in the content of chlorophylls a and b could be related with albino phenomenon in the anther culture. Two standard cultivars of barley were used in this experiment: cv. Cork was used as a genotype producing a high number of albino plants in anther culture and winter barley cv. Igri as a genotype producing high number of green plants (Wojnarowiez et al. 2002). The content of chlorophylls a and b and their ratios in etiolated and androgenic plants were determined.
Results on control plants (plants were not etiolated prior to experiment) grown under standard light/dark regime have shown that the content of chlorophylls in dynamics increases in Igri more intensively than in Cork during ten days (Fig. 1A). In Igri the content of chlorophylls increased by 0.926 µg g-1 per day, and in Cork the content of chlorophylls increased by 0.531 µg g-1 per day.
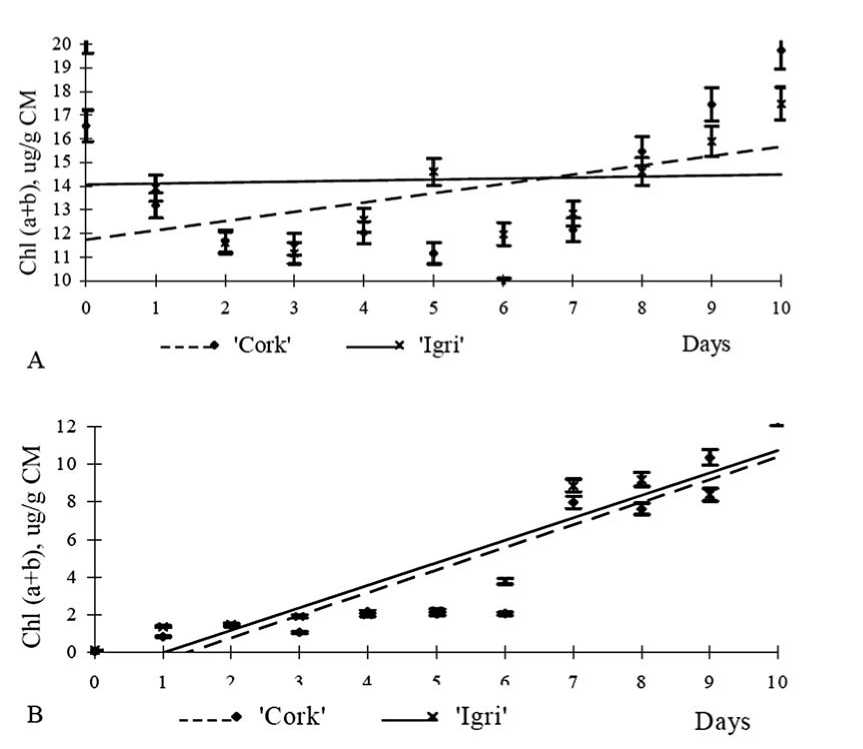
Fig. 1 . Changes of the content of the chlorophylls a and b in ‘Cork’ and ‘Igri’ every twenty-four hours until the tenth day: (A) control seedlings grown under standard light/dark regime (Y (Ic) =0.926x+13.01, R2=0.78, Y (Cc) =0.531x+12.92, R2=0.32) and (B) etiolated seedlings transferred to standard light/dark regime (Y (Ie) =1.384x+0.48, R2=0.87, Y (Ce) =1.221x+0.16, R2=0.83); The content of Chl a and b was measured by spectrophotometric (Lichtenthaler 1987; Kouril et al. 1999) method in 80% acetone at room temperature; CM – crude matter.
It was identified in dynamics for etiolated plants after transferring them for 10 days to the light that the content of chlorophylls in plants of Igri had increased by 1.384 µg g-1 per day, and in the etiolated plants of Cork – by 1.221 µg g-1 per day (Fig. 1B). Chlorophyll a was detected in one minute after the etiolated plants were transferred to the light. This suggests that plants are likely to synthesize protochlorophyllid a in the dark which further, at light, is transformed into chlorophyll a . Results showed that in the etiolated plants after transferring them for 10 days to the light the rate of recovery of photosynthesis in Igri was more intensive than in Cork plants.
The content of chlorophyll a in green plants is twice as that as of chlorophyll b (He et al. 1999). In our experiment we estimated ratios of chlorophylls a and b in dynamics within ten days. In non-etiolated plants the ratio of chlorophylls decreased by 0.112 times in Igri and by 0.060 times in Cork (Fig. 2A).
Theoretically, the optimal a / b chlorophyll ratio is 3/1 (Lichtenthaler 1987). The non-etiolated ‘Cork’ plants reached the optimal 3/1 ratio in shorter time than Igri plants.
The etiolated plants grown in the dark had no chlorophyll and were white and yellow. These plants appear to be very weak, their leaves and tillers did not develop properly. The recovery of a and b chlorophylls started in one minute after transferring them to the light.
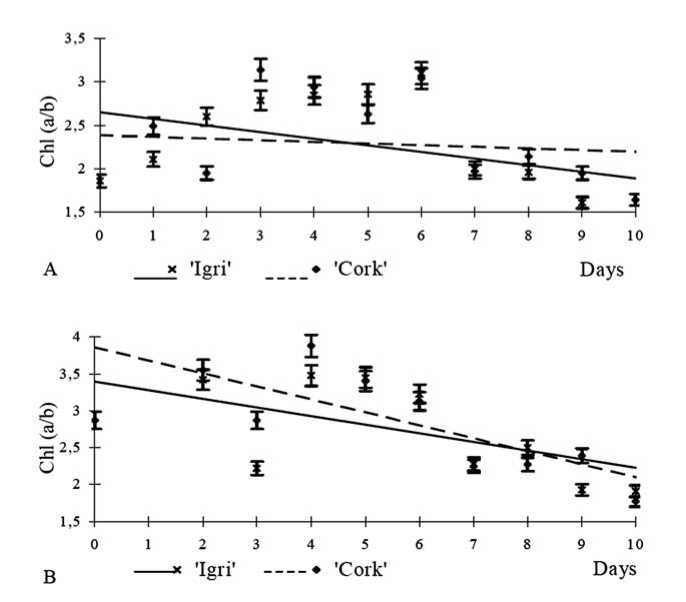
Fig. 2. Changes of chlorophyll a/b ratio in ‘Cork’ and ‘Igri’ every 24 hours up to the 10 days: (A) control seedlings grown under standard light/dark regime (Y ( Ic ) =-0.112x+2.37, R2=0.62, Y ( Cc ) =-0.060x+2.34, R2=0.14) and (B) etiolated seedlings transferred to standard light/dark regime (Y (Ie) =-0.054x+2.65, R2=0.12, Y (Ce) =-0.105x+3.20, R2=0.25). The Chl a / b ratios were measured by spectrophotometric (Lichtenthaler 1987; Kouril et al. 1999) method in 80% acetone are room temperature.
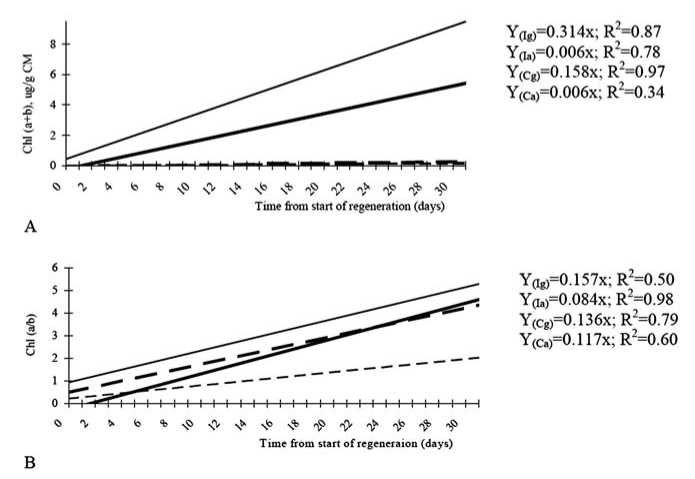
Fig. 3. Changes in the chlorophylls a and b in the regenerants of ‘Cork’ and ‘Igri’ (theoretical lines of regression): (A) the content of chlorophyll a and b ; and (B) the ratio of chlorophylls a / b ( ‘Igri’ – green regenerants, ‘Igri’ – albino regenerants, ‘Cork’ – green regenerants, ‘Cork’ – albino regenerants). The Chl a + b and Chl a / b ratios were measured by spectrophotometric (Lichtenthaler, 1987) method 80% acetone at room temperature; CM, crude matter
The content of chlorophylls a and b in the green regenerants increased in dynamics of 30 days (Fig. 3A). The increase of chlorophyll in green Igri regenerants was determined by 87% by the time from the start of regeneration and only by 13% by other uncontrolled factors. The dynamics of chlorophyll increase in green regenerants of Cork was affected in similar way, i.e. the content of chlorophylls was determined by the time by 97%, and by other uncontrolled factors only by 3%.
Green Igri regenerants had higher content of chlorophyll than green Cork regenerants after 30 days regeneration and growth in the light. This suggests that the efficiency of photosynthesis in the regenerants of Igri is more efficient by 50% than in the green regenerants of Cork.
The albino regenerants do not synthesise chlorophylls because of genetic disorders. Protochlorophylls a and b as were found in the albino regenerants, but not chlorophylls a and b . Changes in the ratio in dynamics showed increase by 0.084 times for regenerants of Igri for every two days during 30 days, and by 0.117 regenerants of Cork.
The genotype appears to be the most important factor in androgenetic albinism, event though the improvement of the anther pretreatment and the culture conditions largely contributed to reduce the amount of albino regenerated plantlets in Hordeum vulgare and Triticum aestivum (Caredda and Clement 1999).
In conclusion, the content of chlorophylls in the etiolated and not etiolated plants during ten days increased at a higher rate in Igri than Cork. This suggests that photosynthesis recovers more efficiently in the etiolated Igri plants than in Cork plants.
The content of chlorophylls a and b in the green regenerants of Igri increased significantly higher than in the green regenerants of Cork. This could be the reason of higher efficiency of photosynthesis in the green regenerants of Igri than of Cork. Photosynthesis was found to be blocked in all albino regenerants both of Igri and of Cork.
References Study of chlorophyll a and b in etiolated and androgenic plants of barley
- www,moksliuke. tai.It
- Andersen, S.B., Due, I.K, Olsen, A, (1987) The response of anther culture in a genetically wide material of winter wheat (Triticum aestivum L.). Plant Breeding, 99, 181-186.
- Caredda, S., Clement, С. (1999) Androgenesis and albinism in Poaceae: influence of genotype and carbohydrates. In: Anther and Pollen: from biology to biotechnology, pp. 211-228.
- Caredda, S., Doncoeur, C., Devaux, P., Sangwan, R.S., Clement, С. (2000) Plastid differentiation during androgenesis in albino and non-albino producing cultivars of barley {Hordeum vulgare L.). Sex Plant Reprod., 13, 95-104.
- Cistue, L., Ziauddin, A., Simion, E., Kasha, K.J. (1995) Effects of culture conditions on isolated microspore response of barley cultivar Igri. Plant Cell Tiss Org Cult., 42, 163-169.
- Dunford, R.P., Walden, R.M. (1991) Plastid genome structure and plastid-related transcript levels in albino barley plants derived from anther culture. Curr Genet., 20, 339-347.
- Dunwell, J.M., Francis, R.J., Powell, W. (1987) Anther culture of Hordeum vulgare L. A genetic study of microspore callus production and differentiation. Theor Appl Genet., 74, 60-64.
- He, Q., Schlich, Т., Paulsen, H., Vermaas, W. (1999) Expression of a higher plant light-harvesting chlorophyll a/b -binding protein in Synechocystis sp. PCC 6803. Eur. J. Biochem., 263, 561-570.
- Jacquard, C., Wojnarowiez, G., Clement, С. (2003) Anther culture in barely. In: Doubled haploid production in crop plants. Spinger, Berlin, Heidelberg, Tokyo, pp. 21-28.
- Jahne, A., Lorz, H. (1995) Cereal microspore culture. Plant Sci., 109, 1-12.
- Kouril, R., Ilik, P., Naus, J., Schoefs, B. (1999) On the limits of applicability of spectrophotometric and spectrofluorimetric methods for the determination of chlorophyll a/b ratio. Photosynthesis Research., 62, 107-116.
- Leon, P., Arroyo, A., Mackenzie, S. (1998) Nuclear control of plastid and mitochondrial development in higher plants. Annu Rev Physiol PlantMol Biol., 49, 453-480.
- Lichtenthaler, H.K. (1987) Chlorphylls and carotenoids: pigments of photosynthetic biomembranes. Methods nzymol., 148, 350-382.
- Mannienen, O. (1997) Optimizing anther culture for barley breeding. Agricultural&Food Science in Finland., Vol.6. N.5/6, 389-398.
- Sarrafi, A., Ghaemi, M. (1997) L'androgenese et son utilisation en genetique et amelioration des plantes.Biotechnologiesvegetates haplodiploidisation, France, pp. 45-88.
- Sunderland, N., Huang, B. (1985) Barley anther culture: the switch of programme and albinism. Hereditas Suppl., 3, 27-40.
- Wojnarowiez, G., Jacquard, C., Devaux, P., Sangwan, R.S., Clement, С. (2002) Influence of copper sulfate on anther culture in barley (Hordeum vulgare L.). Plant Science, 162, 843-847
