Субстраты энергетического обеспечения скелетных мышц при оперативном удлинении костей голени

Автор: Стогов Максим Валерьевич, Еманов Андрей Александрович, Тушина Наталья Владимировна, Смирнов Александр Викторович
Журнал: Гений ортопедии @geniy-ortopedii
Рубрика: Оригинальные статьи
Статья в выпуске: 1, 2012 года.
Бесплатный доступ
Изучен обмен основных субстратов энергообеспечения скелетных мышц при удлинении костей голени по методу Г. А. Илизарова. Показано, что в передней большеберцовой мышце удлиняемого сегмента конечности происходило интен- сивное потребление гликогена, икроножная мышца в качестве энергетических субстратов использовала глюкозу крови и внутриклеточные запасы липидов. Аналогичные мышцы контралатерального сегмента конечности использовали внемы- шечные энергетические субстраты (глюкоза и липиды крови).
Удлинение конечностей, биохимия скелетных мышц, энергетический обмен
Короткий адрес: https://sciup.org/142121506
IDR: 142121506
Substrates of energy supply of skeletal muscles for surgical lengthening of leg bones
The metabolism of the main energy supply substrates of skeletal muscles has been studied for leg bone lengthening according to the Ilizarov method. The intensive glycogen consumption has been demonstrated to occur in the anterior tibial muscle of the limb segment being lengthened, the gastrocnemius muscle has been shown to use blood glucose and intracellular reserves of lipids as energy substrates. It has been determined that the similar muscles of contralateral limb segment use extra-muscular energy substrates (glucose and blood lipids).
Текст научной статьи Субстраты энергетического обеспечения скелетных мышц при оперативном удлинении костей голени
Сократительная функция скелетных мышц обусловлена возможностью превращения энергии биохимических процессов в механическую работу. Метаболизм в скелетных мышцах специализирован таким образом, чтобы достаточно эффективно обеспечивать синтез АТФ для работы сократительной системы, образуя тем самым единый структурнометаболический комплекс, обеспечивающий функциональную активность органа. Для пополнения запаса АТФ в мышцах используются следующие энергетические источники: 1. Анаэробное расщепление углеводов (гликолиз, гликогенолиз) и креатинфосфата; 2. Аэробное расщепление углеводов; 3. Аэробное расщепление липидов (β-окисление) [2, 3]. Вклад каждого из этих источников в энергети- ческое обеспечение мышц при внешних нагрузках зависит как от доступности субстратов окисления в крови [7, 8], так и от запасов внутритканевых источников энергии [4, 9, 10]. Ранее нами было показано, что при удлинении конечностей одним из ключевых процессов в энергообмене скелетных мышц удлиняемого сегмента конечности становится гликолиз [5]. Однако вопрос о субстратах энергообмена (в том числе и гликолиза), а также роль внутритканевых и внетканевых источников энергетического обеспечения мышц при удлинении остается открытым.
Цель исследования — изучить особенности субстратного обеспечения скелетных мышц в условиях оперативного удлинения костей голени по методу Илизарова.
МАТЕРИАЛЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
Исследование проведено на 36 взрослых беспородных собаках, которым в течение 28 дней осуществляли удлинение костей голени по Илизарову. Режим удлинения составлял 1 мм/сутки за 4 приема. Животных выводили из опыта путем внутривенного введения летальных доз барбитуратов через 14, 28 суток дистракции, 15, 30 суток фиксации и 30, 180 суток после снятия аппарата. На проведение экспериментальных и клинических исследований получено разрешение комитета по этике при ФГБУ «РНЦ «ВТО» им. акад. Г. А. Илизарова Минздравсоцразвития РФ».
Содержание животных, оперативные вмешательства и эвтаназию осуществляли в соответствии с требованиями Европейской конвенции по защите экспериментальных животных, «Правилами проведения работ с использованием экспериментальных животных» и требованиями инструкции № 12/313 Министерства здравоохранения РСФСР «Санитарные правила по устройству, оборудованию и содержанию экспериментальных биологических клиник» от 06.01.73 г.
Активность АсАТ, АлАТ, концентрацию лактата в ткани, а также концентрацию глюкозы, общего холестерина, триглицеридов в сыворотке крови определяли на фотометре Stat Fax® 1904 Plus (США), используя наборы реагентов фирмы Vital Diagnostic (РФ). В депротеинизированном саркоплазматическом и сывороточном растворе находили содержание пирувата по методу Umbright в модификации Бабаскина [1]. Уровень гликогена в мышцах определяли непрямым антроновым методом, содержание общих липидов в мышцах находили гравиметрическим методом, после их экстракции хлороформ/метаноловой смесью (2:1) [6]. Общие липиды сыворотки крови определяли с по -мощью наборов реактивов фирмы La Chema (Чехия), общий белок — по Лоури [6].
Достоверность различий между выборками оценивали с помощью W-критерия Вилкоксона для независимых выборок. Корреляционную зависимость оценивали по критерию Спирмена.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Результаты исследования показали, что в ПББМ и ИКМ удлиняемого и контралатерального сегмента конечности в качестве энергетических источников использовались различные субстраты. Так, значительное снижение запасов гликогена в ходе эксперимента мы отмечали только в ПББМ удлиняемого сегмента (табл. 1). При этом уровень гликогена в ПББМ контралатеральной конечности на этапе дистракции и фиксации превышал его содержание в мышцах здоровых животных более чем вдвое. В ИКМ оперированной и контралатеральной конечности содержание гликогена статистически значимо от нормы на всех сроках наблюдения не отличалось.
Одними из энергетических источников в ИКМ на этапах удлинения и фиксации могли являться внутритканевые запасы липидов, снижение уровня которых в ИКМ удлиняемой и контралатеральной конечности мы наблюдали на этапе фиксации и через месяц после снятия аппарата (табл. 2). Уменьшение запасов липидов в мышцах обеспечивало «сохранность» углеводных энергетических резервов, подтверждение чему — обнаруженная нами на этапе дистракции обратная зависимость между уровнем гликогена и общих липидов в ИКМ как оперированной, так и контралатеральной конечности: r = –0,69 (р = 0,05) и r = –0,67 (р = 0,05) соответственно.
Внемышечные источники липидов, присутствующие в крови, либо вообще не использовались мышцами, либо их использование компенсировалось их потреблением, т. к. достоверных изменений концентрации общих липидов, триглицеридов и общего холестерина на сроках наблюдения не обнаруживалось, хотя для общих липидов и триглицеридов наблюдалась тенденция к снижению концентрации в сыворотке крови с конца этапа дистракции до конца фиксации.
Проведенные результаты показывают, что в удлиняемой ПББМ основным субстратом энергообмена являлся гликоген ткани, здесь активировался про- цесс гликогенолиза, в контралатеральной ПББМ и в ИКМ обеих конечностей — гликолиз. Однако, вопреки ожиданию, роста суммарного содержания продуктов гликолиза в мышцах удлиняемого сегмента конечности в ходе эксперимента не наблюдалось, мало того, происходило снижение их концентрации относительно значений здоровых животных (рис. 1).
Наблюдаемое снижение содержания лактата и пирувата в мышцах на фоне активации в них гликолиза связано с: 1) увеличением реакций пере-аминирования в ткани, способствующих утилизации из мышц пирувата и аминного азота в составе аланина в печень (аланиновый цикл); 2) с активацией цикла Кори, осуществляющего перенос лактата взамен глюкозы между мышцами и печенью. В пользу обоих предположений свидетельствовал рост концентрации глюкозы и лактата в сыворотке крови на фоне снижения содержания в ней пирувата (рис. 2). Дополнительное подтверждение этого — обнаруженная нами обратная зависимость между уровнем лактата в сыворотке крови и его содержанием в мышцах удлиняемого сегмента: для ПББМ r (кровь/ПББМ) = (–0,67) (р = 0,05), для ИКМ r (кровь/ИКМ) = (–0,69) (р = 0,05).
В свою очередь, об активации аланинового цикла говорило обнаруженное нами в мышцах увеличение на 28-е сутки дистракции ферментативной активности трансаминаз, ферментов, катализирующих первую реакцию аланинового цикла — реакцию трансаминирования (табл. 3). При этом если в ПББМ удлиняемого сегмента конечности и в ИКМ обеих конечностей увеличивалась активность как АлАТ, так и АсАТ, то в ПББМ контралатеральной конечности возрастала активность только АсАТ. Представленное наблюдение подтверждает высказанное выше предположение об участии аминотрансфераз в утилизации пирувата, образующегося в ткани в результате активации гликолитических процессов.
Таблица 1
Содержание гликогена (мг/100 мг ткани) в скелетных мышцах собак при оперативном удлинении костей голени (Медиана; 25-й÷75-й процентили)
|
Этап эксперимента |
Дистракция |
Фиксация |
После снятия аппарата |
||||
|
мышца |
Здоровые животные (n = 6) |
14-е cутки (n = 5) |
28-е сутки (n = 5) |
15-е сутки (n = 5) |
30-е сутки (n = 5) |
1-й месяц (n = 5) |
6-й месяц (n = 5) |
|
ОПБ |
2,66 |
0,99 0,01 0,53÷2,70 |
1,04 0,05 0,32÷2,71 |
1,33 0,05 0,76÷1,86 |
0,74 0,05 0,49÷1,77 |
4,19 3,35÷5,20 |
4,47 3,51÷4,75 |
|
КПБ |
2,37÷3,46 |
5,58 0,01 4,43÷7,19 |
5,63 0,05 3,62÷6,65 |
4,41 0,05 4,24÷4,77 |
5,06 3,35÷6,02 |
3,92 3,30÷4,52 |
3,98 2,26÷5,19 |
|
ОИМ |
3,53 |
2,78 1,49÷3,88 |
2,48 1,70÷2,87 |
2,61 2,17÷3,29 |
3,57 2,62÷4,57 |
2,43 1,75÷4,17 |
2,83 2,05÷3,50 |
|
КИМ |
2,91÷4,83 |
3,952,÷267,01 |
4,11 2,74÷6,90 |
3,41 2,88÷4,08 |
3,440,÷252,19 |
3,45 2,16÷5,10 |
4,53 2,98÷5,21 |
Примечание. ОПБ и КПБ — передняя большеберцовая мышца удлиняемого и контралатерального сегмента конечности соответственно; ОИМ и КИМ — икроножная мышца удлиняемого и контралатерального сегмента конечности соответственно. Верхний индекс — уровень значимости различий (р) по сравнению со здоровыми животными.
Таблица 2
Содержание общих липидов (% от массы сырой ткани) в скелетных мышцах собак при оперативном удлинении костей голени (Медиана; 25-й÷75-й процентили)
|
Этап эксперимента |
Дистракция |
Фиксация |
После снятия аппарата |
||||
|
мышца |
Здоровые животные (n = 6) |
14-е cутки (n = 5) |
28-е сутки (n = 5) |
15-е сутки (n = 5) |
30-е сутки (n = 5) |
1-й месяц (n = 5) |
6-й месяц (n = 5) |
|
ОПБ |
1,92 |
1,57 1,30÷1,84 |
1,92 1,60÷2,80 |
2,97 2,09÷3,99 |
2,83 2,01÷4,19 |
2,37 1,84÷2,76 |
1,74 1,39÷2,09 |
|
КПБ |
1,38÷2,10 |
2,08 1,69÷2,47 |
1,95 1,92÷2,54 |
2,94 2,55÷3,88 |
2,82 0,05 2,64÷2,93 |
1,96 1,92÷2,46 |
1,99 1,78÷2,15 |
|
ОИМ |
3,02 |
4,15 3,98÷4,34 |
3,28 2,32÷3,33 |
2,40 0,05 1,53÷3,02 |
2,19 0,05 1,81÷2,40 |
1,87 0,01 1,63÷2,11 |
3,34 3,07÷4,16 |
|
КИМ |
2,47÷3,78 |
2,74 2, 56÷2,93 |
2,43 1,92÷3,42 |
3,00 2,44÷2,96 |
2,07 0,03 1,80÷2,25 |
3,30 2,71÷3,44 |
2,80 2,44÷3,11 |
Примечание. ОПБ и КПБ — передняя большеберцовая мышца удлиняемого и контралатерального сегмента конечности соответственно; ОИМ и КИМ — икроножная мышца удлиняемого и контралатерального сегмента конечности соответственно. Верхний индекс — уровень значимости различий (р) по сравнению со здоровыми животными.
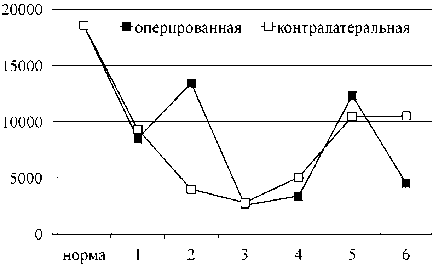
передняя большеберцовая мышца икроножная мышца
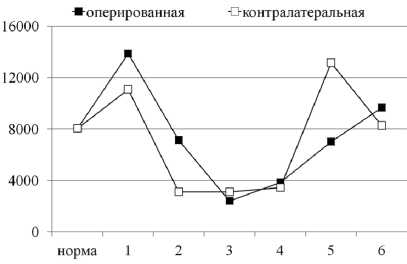
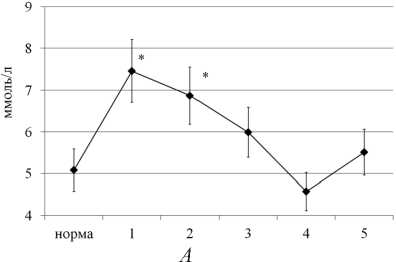
Рис. 2. Концентрация глюкозы (А, ммоль/л), лактата и пирувата (Б, ммоль/л) в сыворотке крови собак в динамике оперативного удлинения костей голени. Примечание: по оси ОХ – сроки эксперимента: 1 — 14-е сутки дистракции; 2 — 28-е сутки дистракции; 3 — 15-е сутки фиксации; 4 — конец фиксации (30-е сутки); 5 — 1 месяц после снятия аппарата. * — статистическая значимость различий по сравнению с нормой при р < 0,05
Рис. 1. Произведение лактат × пируват в скелетных мышцах удлиняемого и контралатерального сегмента конечности собак в условиях оперативного удлинения костей голени. Примечание: по оси ОХ — сроки эксперимента: 1 — 14-е сутки дистракции; 2 — 28-е сутки дистракции; 3 — 15-е сутки фиксации; 4 — конец фиксации (30-е сутки); 5 — 1 месяц после снятия аппарата; 6 — 6-й месяц после снятия аппарата
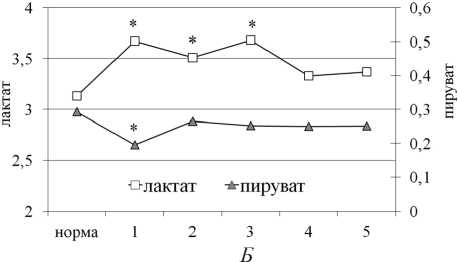
Таблица 3
Активность аспарагиновой (АсАТ) и аланиновой (АлАТ) аминотрансферазы в скелетных мышцах собак при оперативном удлинении костей голени (Медиана; 25-й÷75-й процентили)
|
Этап эксперимента |
АсАТ, мккат/г белка |
АлАТ, мккат/г белка |
|||
|
ПББМ |
ИКМ |
ПББМ |
ИКМ |
||
|
Здоровые животные (n = 6) |
0,108 0,099÷0,130 |
0,084 0,080÷0,107 |
0,276 0,264÷0,277 |
0,237 0,207÷0,250 |
|
|
14-е сутки дистракции (n = 5) |
О |
0,123 0,105÷0,197 |
0,145 0,093÷0,177 |
0,247 0,231÷0,271 |
0,206 0,199÷0,263 |
|
К |
0,099 0,080÷0,123 |
0,092 0,079÷0,112 |
0,243 0,224÷0,290 |
0,248 0,210÷0,293 |
|
|
28-е сутки дистракции (n = 5) |
О |
0,1790,05 0,174÷0,183 |
0,1950,05 0,192÷0,199 |
0,334 0,05 0,307÷0,360 |
0,2940,05 0,274÷0,314 |
|
К |
0,1560,05 0,146÷0,166 |
0,1680,05 0,158÷0,177 |
0,212 0,165÷0,268 |
0,3480,05 0,315÷0,381 |
|
Примечание. ПББМ — передняя большеберцовая мышца, ИКМ — икроножная мышца; О — оперированная (удлиняемая) конечность; К — контралатеральная конечность. Верхний индекс — уровень значимости различий (р) по сравнению со здоровыми животными. * — статистическая значимость различий по сравнению с контралатеральной конечностью при р < 0,05.
ЗАКЛЮЧЕНИЕ
Таким образом, на этапах оперативного удлинения костей голени у собак в скелетных мышцах удлиняемого и контралатерального сегмента в качестве энергетических источников использовались различные субстраты. В ПББМ удлиняемого сегмента конечности основным субстратом энергетического обмена являлся внутриклеточный источник — гликоген. В ПББМ контралатеральной конечности — внемышечные источники энергии, в основном глюкоза крови. В ИКМ оперированного и контралатерального сегмента конечности на этапе дистрак-
ции использовались внетканевые энергетические субстраты (глюкоза и липиды крови), на этапе фиксации — смешанные источники: внутриклеточные липиды, глюкоза крови и, возможно, триглицериды крови. Помимо этого, в ходе оперативного удлинения конечности между мышцами и печенью осуществлялся интенсивный обмен недоокисленных субстратов, обеспечивая их утилизацию из ткани и способствуя поддержанию в ней высокой интенсивности процессов энергетического обмена.
