Суточная динамика CO2-газообмена в системе «болото - атмосфера» в период отсутствия активной вегетации растений

Автор: Михайлов О.А., Загирова С.В., Гончарова Н.Н., Мигловец М.Н.
Журнал: Известия Коми научного центра УрО РАН @izvestia-komisc
Рубрика: Биологические науки
Статья в выпуске: 3 (15), 2013 года.
Бесплатный доступ
Статья посвящена изучению на мезо-олиготрофном болоте средней тайги суточной динамики вертикальных потоков СО2 в приземном слое атмосферы. Впервые с использованием метода микровихревых пульсаций для территории европейского Северо-Востока России установлено, что в холодный период года суточный баланс вертикальных потоков СО2 не превышает 1.1 г/м-2. Величины общего экосистемного обмена СО2 и гросс-фотосинтеза зависят от интенсивности ФАР, а дыхание экосистемы – от температуры поверхности почвы.Весной более низкие значения показателей СО2 -газообмена в системе «болото–атмосфера» могут быть связаны с обводнением значительной части болота после таяния снега.
Средняя тайга, верховое болото, вертикальные потоки со2, метод микровихревых пульсаций, суточная динамика
Короткий адрес: https://sciup.org/14992620
IDR: 14992620 | УДК: 581.55:551.583.1:581.526.35
The diurnal dynamics of CO2 gas exchange in the “peatland - atmosphere” system during the period of active plant vegetation absence
The paper studies the diurnal dynamics of CO2 fluxes in the surface layer at the meso-oligotrophic peatland in middle taiga using the eddy covariance technique. It was found that in cold season the daily balance of CO2 fluxes does not exceed g/m-2. NEE and Pgross values depend on PAR and Reco – on the temperatureof soil surface. Lower values of CO2 gas exchange in spring may be related toflooding much of the peatland after the snow melts.
Текст научной статьи Суточная динамика CO2-газообмена в системе «болото - атмосфера» в период отсутствия активной вегетации растений
Торфяные болота играют ключевую роль в углеродном цикле планеты. За несколько тысячелетий в болотах высоких широт Северного полушария накоплено 400 – 500 Гт углерода, который в условиях потепления климата может стать источником активного поступление парниковых газов в атмосферу [1,2]. Согласно имеющимся данным, направление и величина вертикальных потоков СО 2 в системе «болото – атмосфера» зависят от температуры почвы [3], состояния растительного покрова [4], интенсивности ФАР [5]. Для Европейского Севера данные о величине стока и эмиссии углеродсодержащих парниковых газов в болотных экосистемах немногочисленны. Установлено, что тундровые сообщества с высоким уровнем грунтовых вод в летние месяцы активно поглощают диоксид углерода из атмосферы [6,7]. В болотных экосистемах бореальной зоны в летний период величина стока/эмиссии СО 2 зависит от характера микрорельефа и видового состава растений [8].
Наиболее интенсивно CO2-газообмен на границе «болото – атмосфера» осуществляется в период активной вегетации растений, который в усло- виях средней тайги на территории Республики Коми, по данным многолетних наблюдений, начинается 3 мая и продолжается в среднем 158 дней [9]. На территории Сыктывдинского района Республики Коми, где проводились исследования, окончание периода активной вегетации растений в 2010 г. отмечено 27–28 сентября, когда среднесуточная температура воздуха установилась ниже +5 °С, а его возобновление в 2011 г. – 1–2 мая. Согласно исследованиям, проведенным на севере Финляндии [10], в осенне-зимний период эмиссионный поток диоксида углерода с поверхности болота в атмосферу не прекращается и может достигать 10 – 40% эмиссии, наблюдаемой в конце июня – июле. Однако данные о величине потоков СО2 в системе «болото–атмосфера» на европейском Северо-Востоке в период отсутствия активной вегетации растений не обнаружены. В связи с этим нами была поставлена цель – выявить закономерности суточной динамики вертикальных потоков диоксида углерода в приземном слое атмосферы на мезо-олиготрофном болоте средней тайги в осенне – весенний период.
Объект и методы исследования
Исследования проводили на мезо-олиготрофном болоте Медла-Пэв-Нюр, расположенном в Сыктывдинском районе Республики Коми, в 40 км на северо-запад от г. Сыктывкара (51° 56´ с.ш. 56° 13´ в.д.). Площадь его составляет 2790 га, средняя мощность торфа – 1,4 м, максимальная – 3,4 м. Болото располагается на второй надпойменной террасе, водоприемниками служат р. Пожег и р. Пычим. Торфяная залежь верхового, смешанного, переходного и низинного типа с зольностью 23 – 35% [11].
Геоботанические описания выполнены на четырех площадках 10х10 м, расположенных вдоль трансекты протяженностью 1 км, по общепринятой методике [12]. При классификации типов растительности на исследуемом участке болота использовали аэрофотоснимки. Растительность района исследований представлена следующими сообществами:
-
1) олиготрофные сосново-кустарничково-пуши-циево-сфагновые сообщества, где на кочках доминируют сосна ( Pinus sylvestris L.) , андромеда (Andromeda polifolia L.) , мирт болотный ( Chamaedaphne calyculata (L.) Moench) , клюква ( Oxycoccus microcarpus Turcz. ex Rupr.) , морошка ( Rubus chamaemorus L.) , пушица ( Eriophorum vaginatum L.) , сфагновые мхи ( Sphagnum angusti-folium (C. Jens. ex Russ.) , Sphagnum fuscum (Schimp.)) , Polytrichum strictum Brid., а в мочажинах – андромеда , мирт болотный , клюква ( Oxycoccus palus-tris Pers.) , пушица , сфагновые мхи ( Sphagnum an-gustifolium, Sphagnum ma-gellanicum Brid.);
-
2) мезотрофные кус-тарничково-травяно-сфаг-новые сообщества, с доминированием на кочках андромеды, мирта болотного, багульника ( Ledum palustre L.) , клюквы, пушицы, осоки ( Carex pau-ciflora Lightf.) , сфагновых мхов ( Sphagnum angusti-folium, Sphagnum fuscum, Sphagnum magellanicum ), в мочажинах – осок ( Carex lasiocarpa Ehrh. , Carex limosa L. , Carex rostrata Stokes) , вахты трехлистной ( Menyanthes trifoliata L.) , шейхцерии ( Scheuchzeria palustris L.), мирта болотного, клюквы, пушицы, андромеды, мхов Sphagnum fallax (Klinggr.) , Sphagnum magellanicum, Warnstorfia exannulata (B.S.G.);
-
3) мезоевтрофная травяно-моховая проточная топь с кочковато-топяным микрорельефом, с преобладанием на кочках таких видов, как андромеда, мирт болотный, клюква ( Oxycoccus palustris ) , хвощ ( Equisetum fluviatile L.) , осока ( Carex lasiocarpa ) , мхи р. Sphagnum ( Sphagnum magellanicum, Sphagnum fuscum ), а на топяном ковре – клюква ( Oxycoccus palustris ) , осока ( Carex lasiocarpa ) , пушица ( Eriophorum gracile Koch) , вахта трехлистная, мытник болотный ( Pedicularis palustris L.) , пузырчатка ( Utricu-laria intermedia Hayne) , сфагновые мхи ( Sphagnum fallax, Sphagnum magellanicum ) , Warnstorfia exannu-lata , мхи р. Calliergon.
Для измерений вертикальных потоков СО 2 -газообмена в системе «болото–атмосфера» использовали метод микровихревых пульсаций, детально описанный в работах [13, 14]. Измерительная система закрытого типа, которая включала ультразвуковой анемометр, определяющий направление и скорость ветра в трех проекциях и акустическую температуру воздуха (Solent R3, Gill Instruments Ltd., Великобритания), а также инфракрасный газоанализатор (Li-7000, Li-Cor Inc., США). Отбор проб воздуха проводили на высоте 4,23 м от поверхности болота (без учета снежного покрова) (рис. 1). Данные о скорости ветра и изменении концентрации диоксида углерода регистрировали с
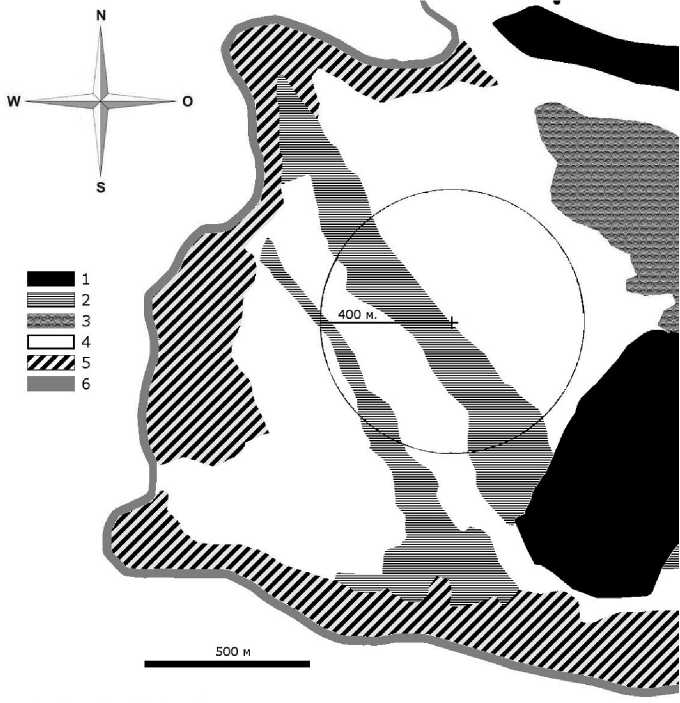
Рис. 1. Схема расположения измерительной системы на участке болота Мед-ла-Пэв-Нюр. 1 – минеральный лесной остров; 2 – мезоевтрофная травяномоховая проточная топь; 3 – олиготрофные сосново-кустарничково-пушициево-сфагновые сообщества; 4 – мезотрофные кустарничково-травяносфагновые сообщества; 5 – окраинная топь; 6 – граница болота.
частотой 20 Гц. Параметры микроклимата (температуру и влажность воздуха, интенсивность ФАР, падающей и отраженной солнечной радиации) регистрировали автоматической метеостанцией (Campbell Scientific, США). Среднюю величину чистого экосистемного обмена диоксида углерода (NEE) , представляющего разницу между величинами валовой первичной продукции экосистемы ( P gross ) и суммарного дыхания ее компонентов ( R eco ), рассчитывали за 30-минутные интервалы с использованием программного обеспечения EdiRe (Robert Clement, Университет Эдинбурга, Великобритания). Контроль качества полученных данных включал: 1) контроль показателей сигнала датчиков, в том числе диапазон разброса и всплески; 2) статистические тесты, выявление резких изменений во временных рядах данных; 3) резкие изменения температуры воздуха, влажности, атмосферного давления за период усреднения данных (30 мин) [15].
Не отвечающие параметрам проверки значения потоков CO 2 не учитывали при дальнейшей обработке данных. Результирующий поток диоксида углерода ( NEE ) разделяли на дыхание экосистемы ( R eco ) и гросс-фотосинтез ( P gross ) по общепринятой методике [3, 16, 17]. Для расчета интенсивности дыхания экосистемы использовали уравнение:
R
eco
= R 10 * exp
308.6*
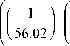
т , surf
+46.02
где R eco – дыхание экосистемы (мкг м-2с-1), R 10 – дыхание экосистемы при температуре воздуха 10 °С (мкг м-2с-1), T surf – температура поверхности почвы (°С). Коэффициент R 10 рассчитывали отдельно для каждого периода измерений.
Значение гросс-фотосинтеза вычисляли, используя выражение:
P = NEE - R gross eco
.
Баланс вертикальных потоков диоксида углерода за сутки рассчитывали по формуле:
NEE.. , -0.0036*((NEE , *N,, ) + diurnal . av.daytime daytime
+( NEE av . nighttime
* N nighttime
)),
где NEE diurnal – суточный экосистемный обмен вертикальных потоков диоксида углерода (г/м-2сут-1), NEE av.daytime – средний экосистемный обмен вертикальных потоков диоксида углерода за период суток, при котором значения ФАР превышали 20 мкмоль м-2с-1 (день), NEE av.nighttime – средний экосистемный обмен вертикальных потоков диоксида углерода за период суток, при котором значения ФАР были меньше 20 мкмоль м-2с-1 (ночь), N daytime – продолжительность дня (час), N nighttime – продолжительность ночи (час).
Измерения СО2-газообмена на болоте проводили с 30 сентября 2010 г. по 28 апреля 2011 г. Для выявления межсезонных различий в скорости потоков СО2 сравнивали данные нескольких суток в осенний (3–4 октября 2010 г.), зимний (22–23 ноября 2010 г.) и весенний (18–19 апреля 2011 г.) периоды года. Краткая характеристика микроклиматических условий в эти дни приведена в табл. 1.
Таблица 1
Характеристики микроклиматических условий на болоте Медла-Пэв-Нюр в дни проведения наблюдений (средние значения)
|
Дни наблюдений |
Т возд , °С |
Т пов , °С |
ФАР*, мкмоль м-2с-1 |
|
3 – 4 октября |
2.6 |
3.6 |
197.4 |
|
22 – 23 ноября |
-9.6 |
-9.2 |
56.4 |
|
18 – 19 апреля |
4.1 |
4.1 |
196.4 |
Примечание. *В расчетах учтены значения ФАР ≥ 20 мкмоль м-2с-1.
Результаты и обсуждение
В исследованиях зарубежных авторов было показано, что переход от стока к эмиссии в болотных экосистемах осенью связан с завершением вегетации растений. В Канаде этот процесс начинается с середины августа, в центре Европейской России и Центральной Сибири – с середины сентября [4, 5].
В среднетаежной подзоне на территории Республики Коми осенний период начинается в конце августа – начале сентября и продолжается до ноября, зима отмечается с ноября по март, а весна наступает в конце марта – начале апреля [9]. Согласно нашим наблюдениям, в октябре в течение суток, несмотря на невысокие значения потоков диоксида углерода, четко выделяются периоды дневного поглощения и ночной эмиссии СО 2 в атмосферу с поверхности болота (рис. 2). Сток углекислого газа из атмосферы в болотные сообщества происходил с 8 до 17 ч и составил в среднем -16,2 мкг/м-2с-1. В остальное время суток отмечена эмиссия СО 2 с поверхности болота со средней скоростью 30,6 мкг/м-2с-1. В результате за сутки суммарная величина NEE составила 1,08 г/м-2. Полученные нами средние величины скорости нетто-обмена диоксида углерода между атмосферой и болотом в октябре сопоставимы с результатами измерений на олиготрофном болоте в Центральной Сибири [5]. Однако они в два раза ниже по сравнению с данными, приведенными для мезотрофного болота на севере Финляндии в осенний период [18]. На исследованном нами участке болота величина NEE за сутки близка к результатам, полученным на верховом болоте в Центральной Сибири [5], и соответствует данным, приведенным для олиготрофного болота Канады [19]. Скорость вертикальных потоков диоксида углерода ( NEE ) в течение дня в этот период года составила 4–10%, а ночью – 38–60% от значений, приводимых другими авторами для летних месяцев [10, 18, 19, 20].
С формированием снежного покрова в начале зимы эмиссионный поток СО 2 с поверхности болота не прекращался: его средняя скорость составила 12,7 мкг/м-2с-1, а суммарное значение за сутки – 1,1 г/м-2.

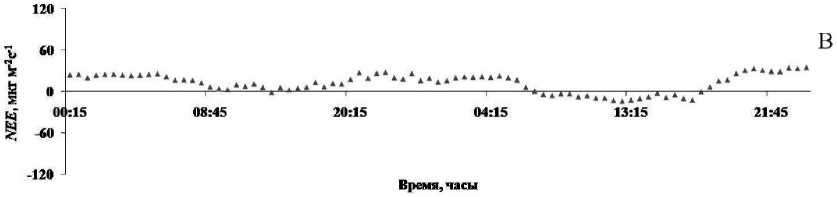
Рис. 2. Суточная динамика NEE с поверхности верхового болота осенью (А), зимой (Б) и весной (В). Примечание: Здесь и далее отрицательные значения соответствуют поглощению, а положительные – выделению СО 2 .
В этот период по причине отсутствия фотосинтеза растений величина NEE равнялась R eco . Полученные нами значения СО 2 газообмена более чем в два раза выше результатов, полученных ранее для мезотрофных болот на северо-востоке Гренландии [20] и на севере Финляндии [10]. Однако они близки к данным, приведенным для олиготрофных болот на юге Швеции [21] и юго-востоке Канады [19].
Ранней весной, с началом таяния снега и оттаиванием верхних горизонтов почвы, когда активизируются процессы дыхания и фотосинтеза у растений, в дневное время суток на болоте нами отмечена слабая эмиссия (8,6 мкг/м-2с-1) или слабый сток СО 2 (-3,7 мкг/м-2с-1). В целом за сутки преобладало выделение СО 2 в атмосферу. Ночью, несмотря на низкую температуру воздуха и почвы, скорость эмиссии диоксида углерода с поверхности болота возрастала и составила в среднем 23,5 мкг/м-2с-1. В этот период величина NEE за сутки оставалась по-прежнему низкой (1 г/м-2). Сходные значения NEE для весенних месяцев приведены в литературе для верховых болот центральной части Европейской России и Центральной Сибири [5].
В экосистемах Субарктики резкое увеличение эмиссии СО2 сразу после схода снега на болоте некоторые авторы связывают с усилением почвенного дыхания на фоне низкого фотосинтеза растений [18] и физической диффузией накопленного за зиму диоксида углерода в торфяной залежи [20]. Переключение эмиссии СО2 на сток в болотной экосистеме происходит очень быстро в течение нескольких дней с началом вегетации травянистых растений и кустарничков [5].
С помощью корреляционного анализа нами сделана попытка установить зависимость показателей СО 2 -газообмена в течение суток от основных микроклиматических факторов в дни проведения наблюдений на болоте (табл. 2).
Таблица 2
Корреляционные зависимости (r)* показателей СО 2 -газообмена от температуры поверхности почвы и интенсивности ФАР за сутки весной и осенью (значимы при р ≤ 0.05)
|
Показатели СО 2 -газообмена |
Осень |
Весна |
||
|
T пов |
ФАР |
T пов |
ФАР |
|
|
NEE (в днев- |
r=0.60, |
r=0.86, |
r=0.10, |
r=0.45, |
|
ное время) |
r=0.000 |
p=0.000 |
p=0.52 |
p=0.002 |
|
R eco (в ночное |
r=0.77, |
r=0.82, |
||
|
время) |
p=0.000 |
p=0.000 |
||
|
P gross (в днев- |
r=0.63, |
r=0.87, |
r=0.56, |
r=0.64, |
|
ное время) |
p=0.000 |
p=0.000 |
p=0.000 |
p=0.000 |
Примечание. * r – коэффициент корреляции пирсола.
Величина NEE в болотной экосистеме в дневное время суток зависела от интенсивности ФАР. Однако весной она менее выражена. Осенью, когда некоторые кустарнички и мхи еще сохраняли способность фотосинтезировать, переход от выделения к поглощению СО 2 на болоте наблюдали при ФАР> 90 мкмоль м-2с-1, а весной – при ФАР >160 мкмоль м-2с-1 (рис. 3). Эти различия могли быть связаны с разрушением части пигментов в листьях кус-
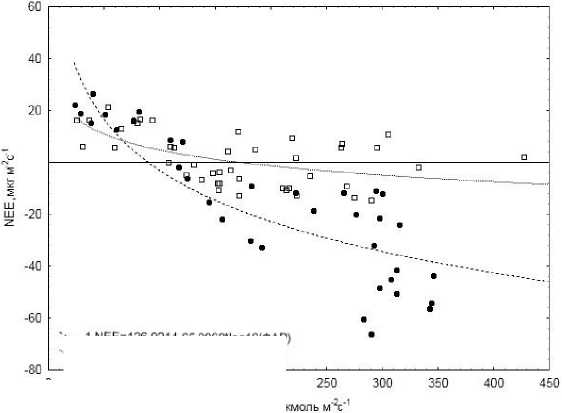
X J N Е Е= 126.9214-65.0968* log 10(Ф АР) \2 NEE=45.4201-20.3156'log 1О(ФАР)
О 50 100 150 200
ФАР м
Рис. 3. Зависимость нетто-обмена СО 2 ( NEE) от интенсивности ФАР: 3–4 октября 2010 г. (1); 18–19 апреля 2011 г. (2).
Заключение
В результате проведенных исследований установлено, что в холодный период года с прекращением вегетации растений в болотной экосистеме среднетаежной подзоны эмиссия СО 2 превышает его сток. Величина нетто-обмена СО 2 ( NEE) за сутки составляет 1,04–1,1 гм-2сут-1. NEE и P gross в дневное время зависят, прежде всего, от интенсивности ФАР, а скорость экосистемного дыхания тесно связана с температурой поверхности почвы. Более низкие показатели СО 2 -газообмена в экосистеме весной, по сравнению с осенью, могут быть обусловлены состоянием фотосинтетического аппарата растений, а также высокой обводненностью болота после таяния снега.
тарничков и мхов зимой, что ограничивало их фотосинтетическую способность в области низкой солнечной радиации в весенний период.
С функциональным состоянием пигментного комплекса в листьях могут быть связаны также различия в скорости фотосинтеза ( P gross ) растений в разные периоды года (рис. 4). Установлено, что при сходных значениях ФАР фотосинтез в сообществах растений болота осенью идет интенсивнее в 3–3,5 раза, чем весной. В осенний период коэффициент корреляции P gross с температурой поверхности почвы также был выше, чем весной (табл. 2). Согласно данным других авторов [22], у кустарничков ранней весной под действием повторяющихся заморозков нарушается транспирационный поток воды в растениях, что приводит к снижению фотосинтеза.
Снижение экосистемного дыхания в ночное время было сопряжено со снижением температуры поверхности почвы, а зависимость R eco от температуры в течение суток описывалась экспоненциальной функцией (рис. 5). Увеличение Т пов от 0 до +4 °С в октябре приводит к возрастанию R eco более, чем в три раза, весной – всего лишь в два раза. Возможно, в апреле гетеротрофное дыхание ограничивается сильной обводненностью болота в результате таяния снега, что в свою очередь снижает дыхание всей экосистемы. В зимний период влияния микроклиматических факторов на вертикальные потоки СО 2 нами не выявлено.
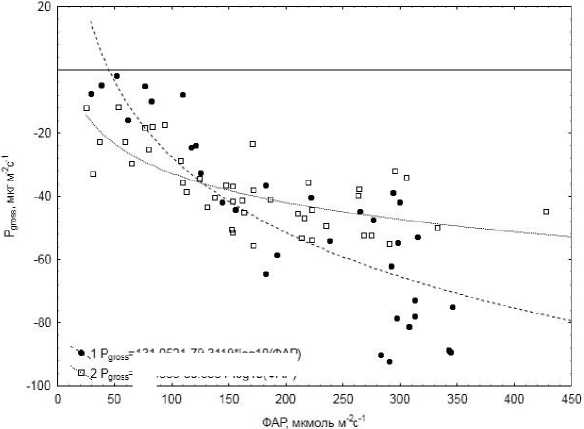
Рис. 4. Зависимость гросс – фотосинтеза ( Pgross) от интенсивности ФАР: 3–4 октября 2010 г. (1); 18–19 апреля 2011 г. (2).
=131.0521 -79.3119'1 од 10(ФАР) -28.5888-30.6534 'log 10 (ФАР)
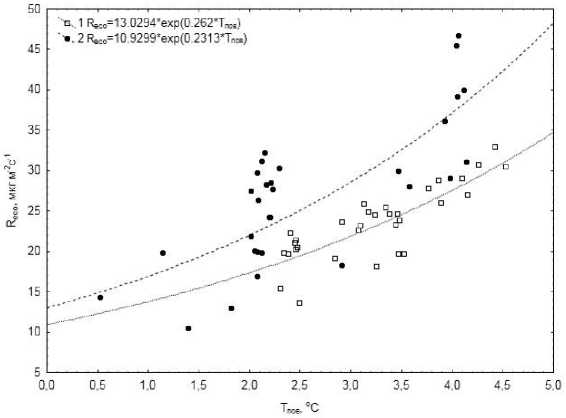
Рис. 5. Зависимость экосистемного дыхания ( Reco) от температуры поверхности почвы; 3–4 октября 2010 г. (1), 18–19 апреля 2011 г. (2).
Работа выполнена при финансовой поддержке проекта ПРООН/ГЭФ 00059042 «Укрепление системы особо охраняемых природных территорий Республики Коми в целях сохранения биоразнообразия первичных лесов в районе верховьев реки Печора» в 2011 г. и научного проекта молодых ученых и аспирантов УрО РАН № 11-4-НП-408 «Эмиссия парниковых газов в экосистеме мезо-олиготрофного болота средней тайги».
Список литературы Суточная динамика CO2-газообмена в системе «болото - атмосфера» в период отсутствия активной вегетации растений
- Gorham E. Northern peatlands: role in the carbon cycle and probable responses to climatic warming//Ecol. Аpplic. 1991. Vol. 1. No 2. P. 182-195.
- Davidson E.A., Janssens I.A. Temperature sensitivity of soil carbon decomposition and feedbacks to climate change//Nature. 2006. Vol. 440. P. 165-173.
- Lloyd J., Taylor J.F. On the temperature dependence of soil respiration//Functional Еcology. 1994. Vol. 8. №3. P. 315-323.
- Lafleur P.M., McCaughey J.H., Joiner D.W., Bartlett P.A. Seasonal trends in energy, water, and carbon dioxide fluxes at a northern boreal wetland//J. Geophys. Res. 1997. Vol. 102. No. D24. Р. 29009 -29020.
- Arneth A., Kurbatova J., Kolle O., et al. Comparative ecosystem-atmosphere exchange of energy and mass in a European Russian and a central Siberian bog II. Interseasonal and interannual variability of CO2 fluxes//Tellus. 2002. No. 54B. P. 514-530.
- Замолодчиков Д.Г., Карелин Д.В., Иващенко А.И. Пороговая температура углеродного баланса южных тундр//Доклады Академии наук. 1998. Т. 358. №5. С. 708-709.
- Heikkinen J.E. P., Virtanen T., Huttunen J. T. et al. Carbon balance in East European tundra//Global Вiogeochem. Сycles. 2004. Vol. 18. GB1023 DOI: 10.1029/2003GB002054
- Михайлов О.А., Загирова С.В., Мигловец М.Н. и др. Оценка потоков диоксида углерода в растительных сообществах мезо-олиготрофного болота средней тайги//Теоретическая и прикладная экология. 2011. №2. С. 44-51.
- Леса Республики Коми/Под ред. Г.М. Козубова и А.И. Таскаева. М.: Дизайн. Информация. Картография, 1999. 332 с.
- Laurila T., Soegaard H., Lloyd C. R. et al. Seasonal variations of net CO2 exchange in European Arctic ecosystems//Theor. and Аppl. Сlimatology. 2001. No. 70. Р. 183-201.
- Торфяные ресурсы Республики Коми/Сост.: Б.Н.Ерцев, Н.В.Мальцева, О.А.Баулина и др. Сыктывкар: Полиграф-сервис, 2000. 613 c.
- Шенников А.П. Введение в геоботанику. Л.: Изд-во ЛГУ, 1964. 447 с.
- Moncrieff J.B., Massheder J.M., de Bruin H. et al. A system to measure surface fluxes of momentum, sensible heat, water vapour and carbon dioxide//J. Нidrology. 1997. No. 188-189. Р. 589-611.
- Baldocchi D.D. Hincks B.B., Meyers T.P. Measuring biosphere-atmosphere exchanges of biologically related gases with micrometeorological methods//Ecology. 1988. Vol. 69. №5. P. 1331-1340.
- Foken Th., Gockede M., Mauder M. et al. Postfield data quality control//Handbook of Мicrometeorology. Springer, Netherlands, 2005. P. 181-208.
- Reichstein M., Falgez E., Baldocchi D. et al. On the separation of net ecosystem exchange into assimilation and ecosystem respiration: review and improved algorithm//Global Сhange Вiology. 2005. No. 11. P. 1424-1439.
- Lasslop G., Reichstein M., Papale D., Richardson A. D. et al. Separation of net ecosystem exchange into assimilation and respiration using a light response curve approach: critical issues and global evaluation//Global Сhange Вiology. 2010. No. 16. P. 187-208.
- Aurela M., Laurila T., Tuovinen J.-P. Seasonal CO2 balances of a subarctic mire//J. Geophys. Res. 2001. Vol. 106. No. D2. Р. 1623-1637.
- Lafleur P.M., Roulet N.T., Admiral S.W. Annual cycle of CO2 exchange at a bog peatland//J. Geophys. Res. 2001. Vol. 106. Р. 3071-3081.
- Nordstroem C., Soegaard H., Christensen T. R., Friborg T., Hansen B. U. Seasonal carbon dioxide balance and respiration of a high-arctic fen ecosystem in NE-Greenland//Theor. and Аppl. Сlimatology. 2001. No. 70. P.149-166.
- Lund M., Lindroth A., Christensen T.R., Ström L. Annual CO2 balance of a temperate bog//Tellus. 2007. No. 59B. P. 804-811.
- Maanavilja L., Riutta T., Aurela M. et al. Spatial variation in CO2 exchange at a northern aapa mire//Biogeochemistry. 2011. No. 104. P. 325-345.
