Сверхэкспрессия гена альтернативной оксидазы способствует поддержанию роста и развития растений арабидопсиса при действии ультрафиолетовой радиации

Автор: Гармаш Елена Владимировна, Епова Арина Александровна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Экология
Статья в выпуске: 3-4 т.15, 2013 года.
Бесплатный доступ
Проведено изучение влияния УФ-А и УФ-Б радиации в естественном соотношении доз на рост, развитие и семенную продуктивность растений Arabidopsis thaliana с генетически модифицированным уровнем экспрессии гена альтернативной оксидазы AOX1а. 3-недельные растения экспонировали в течение 10 дней к УФ облучению в дозе 38 и 0.5 кДж/м 2 в сутки соответственно. УФ-облученные растения мутантной линии XX-2 со сверхэкспрессией AOX1а имели более высокую площадь листьев и массу семян в расчете на растение по сравнению с растениями дикого типа (Col-0) и антисенсовой линии AS-12. Учитывая значение альтернативной оксидазы в предотвращении образования активных форм кислорода, был сделан вывод о том, что усиление экспрессии AOX1a способствовало формированию устойчивости растений к УФ облучению и поддержанию роста в неблагоприятных условиях.
Альтернативная оксидаза, уф радиация, рост, площадь листьев, семенная продуктивность
Короткий адрес: https://sciup.org/148201970
IDR: 148201970 | УДК: 581.1:551.521.13
The overexpression of an alternative oxidase gene contributes to growth and development of arabidopsis plants under uv radiation impact
To study the role of alternative respiratory pathway in plant growth reactions to chronic ultraviolet (UV) radiation, transformed lines of Arabidopsis thaliana were used: AOX1a overexpressors (XX-2), AOX1a anti-sense plants (AS-12). Mutants were compared with the untransformed wild type Columbia (Col-0). Three-week-old Arabidopsis plants were exposed to supplemental UV radiation. The calculated daily doses of UV-A and UV-B were 38 и 0.5 kJ/м 2 respectively. After 10 days of UV-treatment leaf area in the AOX-overexpressing line decreased to a lesser extent compared to the wild type Col-0 and the line AS-12. As a result of the leaf area decrease the plants of the mutant line XX- 2 had a larger seed yield per plant than those of the anti-sense line and the wild type. Since in these transformed plants, AOX protein level affects ROS formation when the cytochrome pathway is restricted, a higher AOX engagement may lead to less ROS production, and hence may maintain more optimal conditions for growth.
Текст научной статьи Сверхэкспрессия гена альтернативной оксидазы способствует поддержанию роста и развития растений арабидопсиса при действии ультрафиолетовой радиации
УФ радиация является одним из сильных стрессовых факторов окружающей среды. УФ излучение, достигающее поверхности Земли, состоит из УФ-А (315-400 нм) и небольшой части УФ-В волн (280-315 нм). УФ ингибирует рост всех органов растения [1] и стимулирует образование активных форм кислорода (АФК), которые в избыточном количестве вызывают окислительный стресс [2, 3]. Важную роль в устойчивости растений к неблагоприятным факторам играют механизмы репарации и/или переключения метаболических путей.
В электрон-транспортной цепи растительных митохондрий (ЭТЦ) помимо основного цитохромного пути (ЦП) транспорт электронов возможен по альтернативному цианидустойчивому пути (АП) через альтернативную оксидазу (АОХ) [4]. В отличие от ЦП электронный транспорт по АП не связан с двумя пунктами генерации мембранного потенциала, поэтому энергетически неэффективен. Считается, что вовлечение АОХ препятствует образованию АФК, защищая клетку от окислительного стресса 5, 6]. Однако информация о роли АОХ в адаптации растений к УФ радиации носит эпизодический характер [7]. Показано, что образуемая под действием УФ-В перекись стимулирует синтез белка АОХ и активирует фермент.
Цель работы состояла в изучении продолжительного влияния УФ-А и УФ-Б радиации в естественном соотношении доз на рост, развитие и семенную продуктивность растений Arabidopsis tha-liana с разным уровнем экспрессии гена AOX1а .
МАТЕРИАЛ И МЕТОДЫ
Объектом исследования послужили растения Arabidopsis thaliana (L.) Heynh. (сем. Brassicaceae)
дикого экотипа и мутантных линий, имеющих разный уровень экспрессии гена АОХ1а . В работе использовали семена арабидопсиса линии Col-0 (растения дикого типа, экотип Columbia-0), линии ХХ-2 со сверхэкспрессией АОХ1а (растения, трансформированные конструкцией, экспрессирующей АОХ1а в сенсовой ориентации), линии AS-12 (растения, трансформированные конструкцией, экспрессирующей АОХ1а в антисенсовой ориентации под контролем промотора CaMV35S). Семена получены из Nottingham Arabidopsis Stock Centre (NASC, Великобритания) и любезно предоставлены В.И. Тарасенко (СИФИБР СО РАН, г. Иркутск) после скрининга на наличие генетической конструкции методом ПЦР [8].
Перед посевом семена выдерживали на влажной фильтровальной бумаге в темноте при +4°С в течение 4 сут. Выращивание осуществляли в почвенной культуре на универсальном грунте в пластиковых стаканчиках объемом 0.2 л. После стратификации растения переносили в условия климатической камеры (Binder, Германия) (Т +22°С, влажность 70%, фотопериод 16 ч, освещенность 65-80 мкмоль/м2 с). Для освещения использовали встроенные в камеру фитолампы OSRAM Lumilux 36 W (Германия).
Часть 3-недельных растений экспонировали к УФ облучению. Использовали лампы УФ Camelion LH26-3U E27 Blacklight 26W (Китай). Доза диапазона УФ-А и УФ-Б составляла 38 и 0.5 кДж/м2 в сутки, что соответствовало уровню УФ облучения в ясный солнечный день.
Для оценки ростовых характеристик использовали выборку как минимум из 20 растений. Площадь листьев определяли при анализе фотоснимков с помощью программы Image Tool for Windows Ver.3.00. В конце репродуктивного периода подсчитывали количество стручков, производили сбор семян, определяли их массу в расчете на растение.
Данные проанализированы с помощью ANOVA с использованием критерия Дункана при уровне значимости P ^ 0.05 (Statistica 6.1 software, StatSoft. Inc., Tulsa, OK, USA).
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
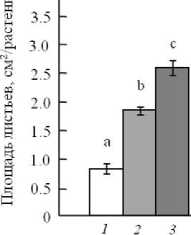
К моменту экспозиции к УФ радиации все линии растений A. thaliana находились в активной фазе вегетативного роста и имели 6-8 розеточных листьев (фаза роста 1.06-1.08, по [9]). Однако в процессе прохождения этапов вегетативного развития растения мутантной линии XX-2 сформировали меньшую площадь листьев по сравнению с диким типом и линией AS-12 (рис. 1). Это указывает на участие АП в энергетическом балансе клетки и влиянии соотношения дыхательных путей на рост растений. Известно, что в оптимальных для роста условиях растительные клетки избегают вовлечения АП, функционирование которого приводит к потере энергии дыхания [10].
УФ-облученные растения проходили быстрее фазы развития (табл.). Под действием УФ фазы роста 6.90 и 8.00, соответствующие завершению цветения и началу образованию плодов (стручков) соответственно, начинались на несколько дней раньше, чем у контрольных растений. Ускорение этапов развития, по-видимому, связано со стратегией растений избежать хроническое негативное влияние УФ на репродуктивный потенциал.
«> 4.0
Col-0
XX-2 AS-12
Рис. 1 . Развитие ассимилирующей поверхности разных мутантных линий Arabidopsis thaliana . Представлены средние и их ошибки (n = 20). Обозначения: 1 – 4 розеточных листа, 2 – 4-8 розеточных листьев, 3 – 6-8 розеточных листьев. Разные надстрочные символы обозначают достоверность изменений параметра (ANOVA, тест Дункана, p ^ 0.05)
Таблица . Влияние УФ радиации на этапы развития растений Arabidopsis thaliana , экотип Col-0 (в соответствии с The Soil-Based Phenotypic Analysis Platform [9])
|
Фаза развития |
Описание |
Дни от посадки |
|
|
Контроль |
УФ |
||
|
1.08 |
8 розеточных листьев |
23-25 |
23-25 |
|
1.10 |
10 розеточных листьев |
25-27 |
24-26 |
|
1.12 |
12 розеточных листьев |
27-29 |
25-27 |
|
1.14 |
14 розеточных листьев |
29-31 |
27-29 |
|
3.20-3.90 |
Рост розетки |
27-33 |
25-30 |
|
6.00 |
Начало цветения |
30-34 |
27-30 |
|
6.90 |
Окончание цветения |
39-45 |
32-36 |
|
8.00 |
Начало плодоношения |
41-47 |
33-38 |
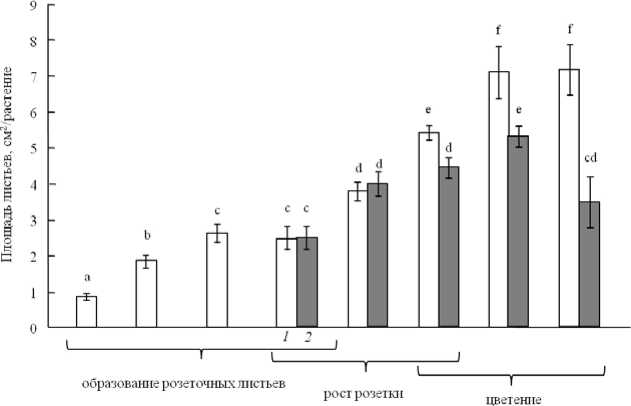
Влияние УФ облучения на параметры ассимилирующей поверхности обнаружено через 10 дней от начала воздействия, когда появление новых листьев прекратилось. Количество розеточных листьев на растении было максимальным и составляло 12-14. Начиная с этого периода, растения были способны развивать ассимилирующую поверхность только за счет увеличения площади листа [9]. Контрольные 5-недельные растения экотипа Col-0 имели площадь листьев 7 см2/раст., что в 1.3 раза выше, чем у УФ-облучаемых особей (рис. 2). Резкое сокращение площади (до 3.5 см2/раст.) под действием УФ обнаружено в конце 6 недели роста. Этот период соответствует середине фазы цветения (фа- за роста 6.30), когда розетка листьев перестает увеличиваться в диаметре.
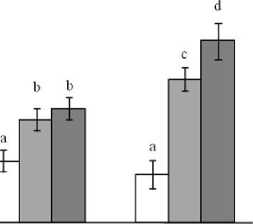
Есть сведения о том, что задержка роста листа при действии УФ-Б радиации, главным образом, связана с ингибированием процессов растяжения клеток адаксиальной поверхности [1]. Как и особи дикого экотипа, растения обеих мутантных линий характеризовались снижением площади листьев под воздействием УФ (рис. 3). Однако площадь ассимилирующей поверхности у линии XX-2 уменьшалась в меньшей степени и составляла после 10 дней УФ облучения 4.5 см2/раст., что в среднем в 1.3 раза больше, чем у растений дикого экотипа и линии AS-12 соответственно. Известно, что в оп- тимальных условиях роста разный уровень экспрессии АОХ в данных мутантных линиях не влиял на про-/антиоксидантное состояние тканей листа [11]. Усиление экспрессии АОХ происходило только при ограничении пропускной способности ЦП специфическими ингибиторами. Можно полагать, что при действии УФ усиленная экспрессия АОХ препятствовала развитию окислительного стресса, создавая более оптимальные условиях для роста.
Сокращение площади листьев при действии УФ повлияло на репродуктивный потенциал растений. Масса семян опытных растений дикого экотипа и растений линии AS-12 составляла в среднем 20 мг/раст., что вдвое ниже, чем в контрольном варианте (рис. 4). Растения линии XX-2 сформировали большее количество семян с массой около 30 мг/раст., что только в 1.4 раза меньше, чем у растений, не подвергшихся воздействию УФ.
Фазы развития
Рис. 2 . Влияние УФ облучения на формирование площади листьев растений дикого типа (Col-0). Представлены средние и их ошибки (n = 20). Обозначения: 1 – контроль, 2 – УФ. Разные надстрочные символы обозначают достоверность изменений параметра (ANOVA, тест Дункана, p ^ 0.05)
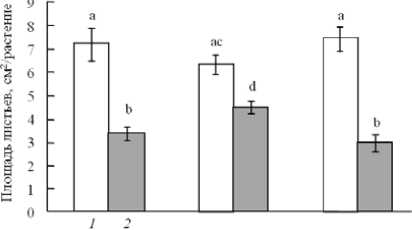
Рис. 3 . Влияние УФ облучения на формирование площади листьев растений дикого типа (Col-0). Представлены средние и их ошибки (n = 20). Обозначения: 1 – контроль, 2 – УФ. Разные надстрочные символы обозначают достоверность изменений параметра (ANOVA, тест Дункана, p ^ 0.05)
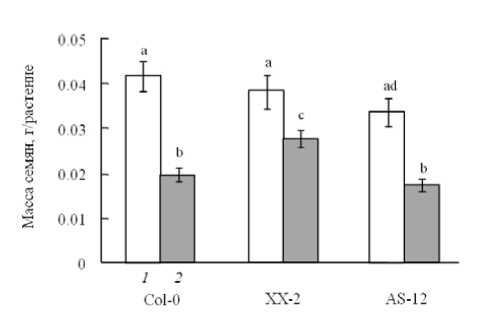
Рис. 4 . Влияние УФ облучения на семенную продуктивность растений разных мутантных линий арабидопсиса. Обозначения как на рис. 2
Таким образом, уровень экспрессии гена AOX1a определял формирование защитных механизмов в процессе адаптации проростков арабидопсиса к УФ облучению. Повышенная активность АОХ, характерная для растений линии XX-2, способствовала под действием УФ развитию ассимилирующей поверхности и поддержанию семенной продуктивности. Результаты согласуются с представлениями о том, что активация АОХ в неблагоприятных для роста условиях обеспечивает баланс энергетического и пластического обмена, защищает клетку от избыточного образования АФК.
Авторы благодарят к.б.н. Р.В. Малышева (ИБ Коми НЦ УрО РАН) за техническое содействие при проведении эксперимента.
Работа поддержана грантами Уральского отделения РАН № 12-У-4-1008, № 12-С-4-1015.
Список литературы Сверхэкспрессия гена альтернативной оксидазы способствует поддержанию роста и развития растений арабидопсиса при действии ультрафиолетовой радиации
- Jansen M. A. K. Ultraviolet-B Radiation Effects on Plants: Induction of Morphogenic Responses//Physiol. Plant. 2002. V. 116. P. 423-429.
- Galatro A., Simontacchi M., Puntarulo S. Free Radical Generation and Antioxidant Content in Chloroplasts from Soybean Leaves Exposed to Ultraviolet-B//Physiol. Plant. 2001. V. 113. P. 564-570.
- Yannarelli G.G., Gallego S.M., Tomaro M.L. Effect of UV-B Radiation on the Activity and Isoforms of Enzymes with Peroxidase Activity in Sunflower Cotyledons//Environ. and Exp. Bot. 2006. V. 56. P. 174-181.
- Vanlerberghe G.C., McIntosh L. Alternative Oxidase: From Gene to Function//Annu. Rev. Plant Physiol. Plant Mol. Biol. 1997. V. 48. P. 703-734.
- Maxwell D.P., Wang Y., McIntosh L. The Alternative Oxidase Lowers Mitochondrial Reactive Oxygen Production in Plant Cells//Proc. Natl. Acad. Sci. USA. 1999. V. 96. P. 8271-8276.
- Rasmusson A.G., Fernie A.R., van Dongen J.T. Alternative Oxidase: a Defence against Metabolic Fluctuations?//Physiol. Plant. 2009. V. 137. P. 371-382.
- Zhao M.-G., Liu Y.-G., Zhang L.-X., Zheng L., Bi Y.-R. Effects of Enhanced UV-B Radiation on the Activity and Expression of Alternative Oxidase in Red Kidney bean Leaves//J. Integr. Plant Biol. 2007. V. 49. P. 1320-1326.
- Тарасенко В.И., Гарник Е.Ю., Константинов Ю.М. Экспрессия гена gdh2 арабидопсиса зависит от активности альтернативного переноса электронов в митохондриях//Biopolymers and Cell. 2012. V. 28. P. 363-367.
- Boyes D.C., Zayed A.M., Ascenzi R., McCaskill A.J., Hoffman N.E., Davis K.R., Görlach J. Growth Stage-Based Phenotypic Analysis of Arabidopsis: A Model for High Throughput Functional Genomics in Plants//The Plant Cell. 2001. V. 13. P. 1499-1510.
- Priault P., Vidal G., De Paepe R., Ribas-Carbo M. Leaf Age-Related Changes in Respiratory Pathways are Dependent on Complex I Activity in Nicotiana sylvestris//Physiol. Plant. 2007. V. 129. P. 152-162.
- Umbach A.L., Fiorani F., Siedow J.N. Characterization of Transformed Arabidopsis with Altered Alternative Oxidase Levels and Analysis of Effects on Reactive Oxygen Species in Tissue//Plant Physiology. 2005. V. 139. P. 1806-1820.
