Сверхмедленные колебания потенциалов нейромодуляторных центров головного мозга и корковых отделов сенсорных систем

Автор: Пугачев К.С., Кребс А.А., Филиппов И.В., Зюзин Е.В.
Журнал: Известия Коми научного центра УрО РАН @izvestia-komisc
Рубрика: Биологические науки
Статья в выпуске: 1 (17), 2014 года.
Бесплатный доступ
Выявлена и проанализирована динамика сверхмедленных колебаний потенциалов нейромодуляторных центров головного мозга крыс (голубого пятна, дорсального ядра шва и базального крупноклеточного ядра). В этих структурах установлено присутствие сверхмедленной активности секундного и обнаружены средние значения (0,45≤r≤0,65) коэффициентов линейной корреляции многосекундных волн, что указывает на их участие в модулирующих влияниях указанных структур на процессы переработки сенсорной информации в первичной зрительной слуховой и вкусовой коре головного мозга.
Сверхмедленные колебания потенциалов, голубое пятно, дорсальное ядро шва, базальное крупноклеточное ядро, первичная зритель- ная, слуховая и вкусовая кора
Короткий адрес: https://sciup.org/14992662
IDR: 14992662 | УДК: 612.826.5:612.826.8:612.825.1
Infraslow brain potentials of neuromodulatory centers and highest cortical sensory areas
We identified and studied the dynamical properties of the infraslow brain poten- tials of different neuromodulatory centers of the rats - locus coeruleus, nucleus raphe dorsalis and nucleus basalis magnocellularis. Moderate linear correlation (0.45≤r≤0.65) of multisecond waves between neuromodulatory centers and cortical sensory areas indicates the possible involvement of this infraslow domain in modulatory influences of locus coeruleus, dorsal raphe nucleus and nucleus basalis magnocellularis on sensory information neuroprocessing in the primary visual auditory and gustatory cortices.
Текст научной статьи Сверхмедленные колебания потенциалов нейромодуляторных центров головного мозга и корковых отделов сенсорных систем
Известно, что различные биоэлектрические процессы головного мозга являются коррелятами его функциональной активности [1].Тем не менее в клинических и фундаментальных дисциплинах не отводится должного внимания сверхмедленным биоэлектрическим процессам, протекающим в ЦНС. В связи с этим все большую актуальность приобретают исследования, направленные на понимание роли сверхмедленных колебаний потенциалов (СМКП) в функционировании головного мозга [2 – 4]. В соответствии с общепринятыми взглядами, СМКП представляют собой совокупность сложноорганизованной квазипериодической динамики биопотенциалов головного мозга человека и животных с частотами менее 0,5 Гц [5]. Исходя из существующей классификации [1], СМКП подразделяются на следующие частотные диапазоны: секундные (период от 2 до 10 с), многосекундные (от 10 до 60 с) и минутные (более 1 мин) волны. Ранее было доказано присутствие СМКП в различных сенсорных представительствах головного мозга: в первичной зрительной коре (ПЗК), первичной слуховой коре (ПСК), во вкусовой коре (ВК), а также в латеральном коленчатом теле, медиальном коленчатом те- ле и в вентральном задне-срединном ядре таламуса [2–4]. Также установлено участие СМКП в процессах переработки сенсорной информации в этих структурах и во взаимодействиях перечисленных отделов ЦНС при переработке сенсорной информации различных модальностей. На основании полученных в наших исследованиях данных была выдвинута гипотеза о возможной обусловленности динамики СМКП в первичных корковых представительствах сенсорных систем с динамикой аналогичных процессов на уровне различных нейромо-дуляторных центров. Это представляется возможным в контексте существующих сведений о присутствии норадренергических (от голубого пятна), серотонинергических (от дорсального ядра шва) и ацетилхолинергических (от базального крупноклеточного ядра) волокон в различных отделах коры больших полушарий, в том числе в сенсорных представительствах неокортекса [6, 7]. Однако до настоящего времени данные о наличии СМКП и об их сравнительном анализе в различных нейромо-дуляторных центрах отсутствовали, в частности голубом пятне (ГП), дорсальном ядре шва (ДЯШ) и базальном крупноклеточном ядре (БКЯ), а также в высших корковых представительствах зрительной, слуховой и вкусовой сенсорных систем. Все эти вопросы представляют существенный интерес, поскольку позволяют более детально понять вклад (норадреналина, серотонина и ацетилхолина) в модуляцию как функциональных состояний различных отделов неокортекса, так и переработку афферентной информации на уровне высших сенсорных представительств ЦНС.
Учитывая актуальность проблемы, цель настоящей работы: выявить в условиях отсутствия действия сенсорных стимулов сверхмедленные колебания потенциалов в голубом пятне, дорсальном ядре шва и базальном крупноклеточном ядре и сопоставить у анестезированных крыс в условиях хронического эксперимента динамику СМКП в указанных нейромодуляторных центрах с аналогичными процессами на уровне первичной зрительной, слуховой и вкусовой коры.
Основные задачи исследования:
-
1. Выявить и проанализировать динамику сверхмедленных колебаний потенциалов в голубом пятне, дорсальном ядре шва и базальном крупноклеточном ядре головного мозга крыс при отсутствии действия зрительных, слуховых и вкусовых раздражителей.
-
2. Сопоставить временные и амплитудночастотные характеристики сверхмедленной активности голубого пятна, дорсального ядра шва и базального крупноклеточного ядра с аналогичными процессами в области первичной зрительной, слуховой и вкусовой коры в тех же условиях.
Материалы и методы
Работа проведена на 30 самцах взрослых нелинейных крыс альбиносов с массой тела 230– 280 г (n=150 повторных экспериментальных наблюдений). Все опыты выполнены согласно основным положениям Европейской конвенции об обращении с животными при проведении биомедицинских исследований (2010/63/EU). Первоначально, руководствуясь атласом головного мозга крыс [8], каждому экспериментальному животному под нембуталовым наркозом производилась стереотаксическая имплантация золотых долгосрочных интрацеребральных электродов (золото 98 %, площадь активной поверхности 0,2 мм2) в ГП, ДЯШ, БКЯ, а также в ПЗК, ПСК и ВК. Использование золотых электродов объясняется современными представлениями о том, что металлические электроды оптимальны для изучения очень медленных форм биоэлектрической активности ЦНС в условиях хронического эксперимента [9]. До начала проведения стереотаксической операции каждая электродная пара была предварительно протестирована в физиологическом растворе, а для имплантации использовались лишь те электродные пары, которые имели стабильный межэлектродный потенциал. После операции крысы помещались на 14 суток в стандартные условия вивария с целью завершения репаративных процессов в исследуемых структурах ЦНС.
Затем осуществляли многократную монопо-лярную синхронную регистрацию СМКП в голубом пятне, дорсальном ядре шва и базальном крупноклеточном ядре, а также в ПЗК, ПСК и ВК. Для этого животные помещались в звуко-свето-непроницаемый индивидуальный контейнер, который располагался в экранированной камере. Эксперименты проводились в темноте (0 люкс) и тишине (0 дБ). Крысы во время исследований находились под легким уретановым наркозом (0,2–0,3 г/кг, интраперитонеально). Использование уретана объясняется тем, что, по данным литературы, он практически не искажает нативную динамику СМКП головного мозга [10]. Для усиления, записи и анализа СМКП применялся компьютерный электрофизиологический комплекс с высокоимпен-дансным многоканальным усилителем биопотенциалов (модель УУ-93, ЭПМ НИИЭМ РАМН), аналогоцифровым преобразователем (модель Е-154, Л-Кард) и персональным компьютером с набором соответствующего программного обеспечения.
Изучение динамики СМКП предусматривало амплитудно-временной анализ различных диапазонов СМКП, основанный на применении алгоритма быстрого прямого преобразования Фурье, а также корреляционный анализ. Статистическую значимость отличий оценивали с использованием однофакторного дисперсионного анализа. Отличия с p<0,05 рассматривались как статистически значимые. После завершения экспериментальных исследований осуществлялся забой крыс (уретан, 2-2,5 г/кг, интраперитонеально) и производилась морфологическая верификация расположения активной части электродов в исследуемых структурах головного мозга.
Результаты и обсуждение
Анализ и обобщение полученных в ходе настоящего исследования данных позволили установить, что для основных структур норадренергической (ГП), серотонинергической (ДЯШ) и холинергической (БКЯ) системы характерно наличие фоновых спонтанных сверхмедленных колебаний потенциалов различных частотных диапазонов. Это были регулярные синусоидальные секундные волны с преобладанием частот в диапазоне от 0,1 до 0,32 Гц и амплитудой до 0,3 мВ в голубом пятне (рис. 1а; 2а). В дорсальном ядре шва доминировали секундные СМКП с частотами от 0,1 до 0,33 Гц и амплитудой до 0,15 мВ (рис. 3а, 4а). Схожие по частотно-амплитудным характеристикам колебания потенциалов секундного диапазона зарегистрированы и в базальном крупноклеточном ядре – преобладающие частоты находились в пределах от 0,1 до 0,2 Гц и с амплитудой до 0,25 мВ (рис. 5а, 6а).
Помимо секундных СМКП были обнаружены регулярные синусоидальные многосекундные волны во всех изученных структурах в диапазонах частот 0,0167–0,04 Гц и амплитудой до 0,27 мВ в голубом пятне (рис. 1б, 2б), 0,0167–0,035 Гц и амплитудой до 0,15 мВ в дорсальном ядре шва (рис. 3б, 4б), 0,0167–0,047 Гц и амплитудой до 0,2 мВ в базальном крупноклеточном ядре (рис. 5б, 6б).
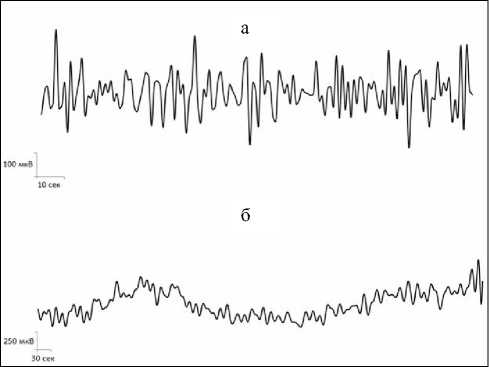
Рис. 1. Примеры нативных записей сверхмедленных колебаний потенциалов различных диапазонов в голубом пятне головного мозга крысы: (а) волны секундного диапазона, (б) волны многосекундного и минутного диапазонов.
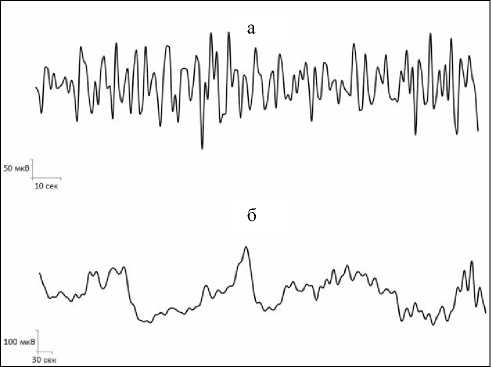
Рис. 3. Примеры нативных записей сверхмедленных колебаний потенциалов различных диапазонов в дорсальном ядре шва головного мозга крысы: (а) волны секундного диапазона, (б) волны многосекундного и минутного диапазонов.
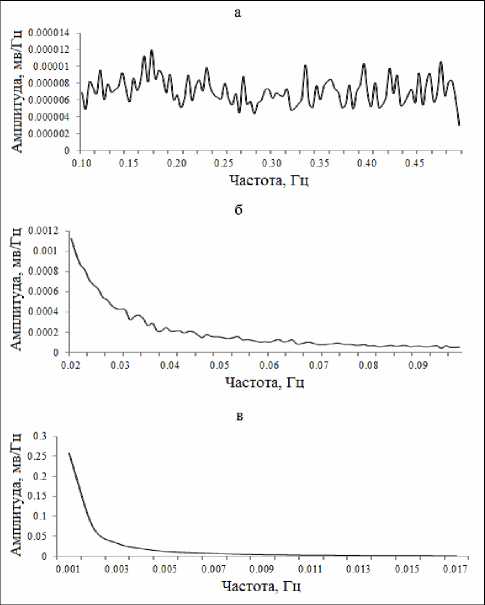
Рис. 2. Суммарные арифметически усредненные спектрограммы (по данным n=50 экспериментов) СМКП различных диапазонов в голубом пятне головного мозга крыс: (а) секундные, (б) многосекундные и (в) минутные волны.
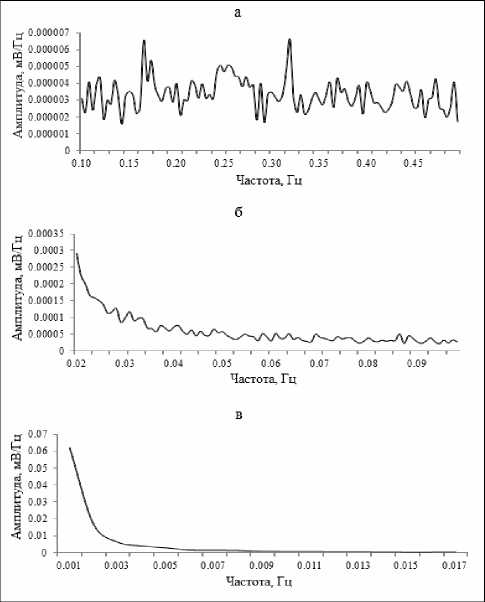
Рис. 4. Суммарные арифметически усредненные спектрограммы (по данным n=50 экспериментов) СМКП различных диапазонов в дорсальном ядре шва головного мозга крыс: (а) секундные, (б) многосекундные и (в) минутные волны.
В голубом пятне, дорсальном ядре шва и базальном крупноклеточном ядре также обнаружены спонтанные одиночные минутные волны синусоидальной формы с частотой менее 0,006 Гц (рис. 1б, 2в; 3б, 4в; 5б, 6в). Подобные результаты позволяют сделать заключение о том, что для каждого из изученных нейромодуляторных центров характерно явление спонтанной фоновой сверхмедленной ритмической биоэлектрической активности.
Кроме того, нами зарегистрированы и проанализированы фоновые сверхмедленные колебания потенциалов в первичных корковых представительствах сенсорных систем головного мозга крыс – в первичной зрительной, первичной слуховой и вкусовой коре. Данные о динамике СМКП первичной зрительной коры, первичной слуховой коры и вкусовой коры головного мозга крыс описаны и ранее [2–4]. Несмотря на это, они проанали-
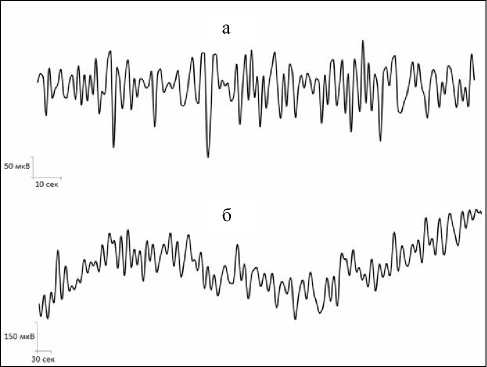
Рис. 5. Примеры нативных записей сверхмедленных колебаний потенциалов различных диапазонов в базальном крупноклеточном ядре головного мозга крысы: (а) волны секундного диапазона, (б) волны многосекундного и минутного диапазонов.
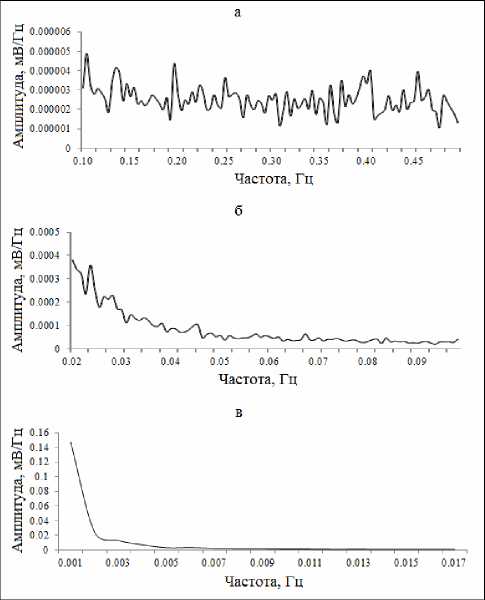
Рис. 6. Суммарные арифметически усредненные спектрограммы (по данным n=50 экспериментов) СМКП различных диапазонов в базальном крупноклеточном ядре головного мозга крыс: (а) секундные, (б) многосекундные и (в) минутные волны.
зированы нами в контексте взаимосвязи динамики СМКП изученных нейромодуляторных центров и первичных корковых сенсорных представительств. Это связано с необходимостью сопоставить фоновые взаимосвязи СМКП в интрацеребральных функциональных системах нейромодуляторный центр – первичные корковые сенсорные представительства (т.е. ГП-ПЗК, ГП-ПСК, ГП-ВК; ДЯШ-ПЗК,
ДЯШ-ПСК, ДЯШ-ВК; БКЯ-ПЗК, БКЯ-ПСК и БКЯ-ВК) в условиях отсутствия действия зрительных, слуховых и вкусовых раздражителей. С этой целью проведена линейная корреляция фоновых СМКП различных диапазонов в парах ГП-ПЗК, ГП-ПСК, ГП-ВК; ДЯШ-ПЗК, ДЯШ-ПСК, ДЯШ-ВК; БКЯ-ПЗК, БКЯ-ПСК и БКЯ-ВК.
В результате анализа указанных данных установлены слабые положительные корреляционные связи в диапазоне секундных волн во всех изученных парах (таблица). Коэффициенты линейной корреляции, отражающие взаимосвязь СМКП в многосекундном и минутных диапазонах (т.е. в едином объединенном диапазоне частот 0,0167–0,1 Гц) можно охарактеризовать как положительные средние.
Коэффициенты линейной корреляции фоновых нативных записей (по данным n=150 экспериментов) различных диапазонов СМКП
|
Пары сравнения |
Коэффициент линейной корреляции (r) секундных СМКП |
Коэффициент линейной корреляции (r) многосекундных и минутных СМКП |
|
ГП-ПЗК |
0,24 |
0,64 |
|
ГП-ПСК |
0,22 |
0,63 |
|
ГП-ВК |
0,14 |
0,61 |
|
ДЯШ-ПЗК |
0,28 |
0,63 |
|
ДЯШ-ПСК |
0,24 |
0,65 |
|
ДЯШ-ВК |
0,14 |
0,60 |
|
БКЯ-ПЗК |
0,25 |
0,64 |
|
БКЯ-ПСК |
0,20 |
0,57 |
|
БКЯ-ВК |
0,17 |
0,45 |
Примечание: пары сравнения – голубое пятно – первичная зрительная, слуховая и вкусовая кора; дорсальное ядро шва – первичная зрительная, слуховая и вкусовая кора; базальное крупноклеточное ядро – первичная зрительная, слуховая и вкусовая кора.
При этом следует обратить внимание на положительные средние значения коэффициентов линейной корреляции, которые указывают на фоновую спонтанную взаимосвязь СМКП многосекундного и минутного диапазонов во всех проанализированных парах сравнения (ГП-ПЗК, ПСК, ВК; ДЯШ-ПЗК, ПСК, ВК; БКЯ-ПЗК, ПСК, ВК). Эти данные сопоставимы с результатами исследований связей нейромодуля-торных центров с различными отделами неокортек-са (норадренергические, серотонинергические и холинергические проекции), в том числе с высшими корковыми представительствами зрительной, слуховой и вкусовой сенсорных систем [11 – 13]. Они указывают на тесные нейроанатомические моноси-наптические связи ГП, ДЯШ и БКЯ с ПЗК, ПСК и ВК, что предположительно является структурной основой подобных функциональных взаимодействий в системах ГП-ПЗК, ПСК, ВК; ДЯШ-ПЗК, ПСК, ВК; БКЯ-ПЗК, ПСК, ВК.
Трудность анализа полученных данных сопряжена с отсутствием в литературе сведений о динамике СМКП в ГП, ДЯШ и БКЯ. Для интерпретации в парах изученных структур корреляционных связей многосекундных СМКП, а следовательно и характера взаимодействий нейромодуляторных, можно привлечь существующие нейрофизиологические и нейрохимические работы. Например, показано, что активация ГП при действии сенсорных стимулов (как и его электростимуляция) вызывает увеличение выделения норадреналина и его производных метаболитов в коре больших полушарий [14, 15]. Установлено также, что различные ритмы ЭЭГ (низкоамплитудная высокочастотная активность и/или высокоамплитудные медленные волны) в коре, по-видимому, связаны с импульсной активностью ГП, которое оказывает регулирующее влияние на уровень возбудимости корковых нейронов [16].
Изучение особенностей тета-ритма у крыс выявило участие ДЯШ и серотонина в этих процессах, и существенный вклад серотонинергической системы ДЯШ в регуляцию уровня бодрствования. Продемонстрированы непосредственные влияния ДЯШ на процессы поддержания уровня активности популяций нейронов неокортекса [17, 18].
Существуют данные об участии БКЯ в модуляции функциональных состояний коры и уровня ответных реакций «настораживания» при действии краткосрочных сенсорных стимулов [19]. Также предполагается, что БКЯ изменяет эндогенную ос-цилляторную активность нейронов пейсмейкеров неокортекса, что указывает на важнейшую роль холинергических нейронов БКЯ в модуляции биоэлектрической активности коры и ее функциональных состояний [20].
Приведенные работы (косвенно) сопоставимы с полученными нами данными о средней степени обусловленности наиболее медленных диапазонов СМКП, протекающих в ПЗК, ПСК и ВК, c динамикой аналогичных биоэлектрических процессов на уровне рассмотренных нейромодуляторных центров. Однако следует обратить внимание и на то, что в секундном диапазоне СМКП наблюдались слабые значения корреляционных связей в системах ГП-ПЗК, ПСК, ВК; ДЯШ-ПЗК, ПСК, ВК; БКЯ-ПЗК, ПСК, ВК. Эти результаты соответствуют нашим предыдущим результатам о вовлеченности СМКП секундного диапазона в специфические процессы переработки сенсорной информации, тогда как потенциалы многосекундного диапазона отражали глобальные колебания уровня общей возбудимости нейронов неокортекса [2 – 4]. Поскольку в настоящей работе записи СМКП в нейромодуляторных центрах и в высших корковых представительствах изученных сенсорных систем проводились в условиях отсутствия действия соответствующих по своей модальности сенсорных раздражителей, можно обоснованно предположить, что в этих условиях переработка сенсорной информации в ПЗК, ПСК и ВК отсутствовала. Это и объясняет слабые корреляционные связи динамики секундного диапазона в системах ГП-ПЗК, ПСК, ВК; ДЯШ-ПЗК, ПСК, ВК; БКЯ-ПЗК, ПСК, ВК. В то же самое время, по-видимому, даже в этих условиях со стороны ГП, ДЯШ и БКЯ существуют постоянные фоновые спонтанные сверхмедленные регуляторные влияния на общий уровень возбудимости нейронов ПЗК,
ПСК и ВК, что и объясняет выраженные корреляционные связи динамики многосекундных СМКП между этими структурами в состоянии покоя.
Выводы
-
1. В голубом пятне, дорсальном ядре шва и базальном крупноклеточном ядре продемонстрировано наличие фоновых эндогенных спонтанных сложноорганизованных сверхмедленных колебаний потенциалов секундного (0,1–0,5 Гц), многосекундного (0,0167–0,1 Гц) и минутного (менее 0,0167 Гц) диапазонов.
-
2. Голубое пятно, дорсальное ядро шва и базальное крупноклеточное ядро оказывают постоянные регуляторные влияния на глобальный уровень возбудимости нейронов первичной зрительной, слуховой и вкусовой коры, что преимущественно отражается в средних значениях коэффициентов линейной корреляции многосекундных (0,0167–0,1 Гц) волн сверхмедленных колебаний потенциалов между этими структурами.
Данная поисковая научно-исследовательская работа выполнена в рамках реализации ФЦП «Научные и научно-педагогические кадры инновационной России на 2009–2013 гг.», а также при частичной поддержке гранта РФФИ (проект 1404-00028).
Список литературы Сверхмедленные колебания потенциалов нейромодуляторных центров головного мозга и корковых отделов сенсорных систем
- Илюхина В.А. Мозг человека в механизмах информационно-управляющих взаимодействий организма и среды обитания. СПб.: Институт мозга человека РАН, 2004. 321 c.
- Филиппов И.В., Кребс А.А., Пугачев К.С. Сверхмедленная биоэлектрическая активность структур слуховой системы головного мозга//Сенсорные системы. 2006. Т. 20. №3. С. 238-244.
- Филиппов И.В. Сверхмедленные колебания потенциалов латерального коленчатого тела и первичной зрительной коры как корреляты процессов переработки зрительной информации//Сенсорные системы. 2007. Т. 21. № 3. С. 165-173.
- Филиппов И.В., Кребс А.А., Пугачев К.С. Сверхмедленные колебания потенциалов центральных представительств вкусовой системы головного мозга крыс при действии различных вкусовых стимулов//Сенсорные системы. 2008. Т. 22. № 2. С. 162-174.
- Аладжалова Н.А. Медленные электрические процессы в головном мозге. М., 1962. 240 с.
- Nakamura K., Yamamoto M., Takahashi K., et al. State-dependency of neuronal slow dynamics during sleep observed in cat lateral geniculate nucleus//Sleep Res. Online. 2000. Vol. 3. P. 147-157.
- Novak P., Lepicovska V. Slow modulation of EEG//Neuroreport. 1992. Vol. 3. P. 189-192.
- Swanson L.W. Brain Maps: Structure of the Rat Brain//Second revised edition. Elsevier.: Amsterdam, 1998. 267 p.
- Ikeda A., Nagamine T., Yarita M. et al. Reappraisal of the effect of electrode property on recording slow potentials//Electroencephalogr. Clin. Neurophysiol. 1998. Vol. 107. P. 59-63.
- Ebenezer I.S. The generation of cortical slow potentials in the rat anaesthetized with urethane and their modification by nicotine//Neuropharmacol. 1986. Vol. 25. № 6. P. 639-643.
- Waterhouse B.D., Border B., Wahl L. et al. Topographic organization of rat locus coeruleus and dorsal raphe nuclei: distribution of cells projecting to visual system structures//J. Comp. Neurol. 1993. Vol. 336. № 3. P. 345-361.
- Blasiak T., Lewandowski M.H. Dorsal raphe nucleus modulates neuronal activity in rat intergeniculate leaflet//Behav. Brain Res. 2003. Vol. 138. № 2. P. 179-185.
- Robbins T.W., Everitt B.J., Ryan C.N., et al. Comparative effects of quisqualic and ibotenic acidinduced lesions of the substantia innominata and globus pallidus on the acquisition of a conditional visual discrimination: differential effects on cholinergic mechanisms//Neuroscience. 1989. Vol. 28. № 2. P. 337-352.
- Salgado H., Garcia-Oscos F., Patel A., et al. Layer-specific noradrenergic modulation of inhibition in cortical layer II/III//Cereb. Cortex. 2011. Vol. 21. № 1. P. 212-221.
- Anlezark G.M., Walter D.S., Arbuthnott G.W. et al. The relationship between noradrenaline turnover in cerebral cortex and electrical self-stimulation through electrodes in the region of locus coeruleus//J. Neurochemistry. 1975. Vol. 24. № 4. P. 677-681.
- Eschenko O., Magri C., Panzeri S. et al. Noradrenergic Neurons of the Locus Coeruleus Are Phase Locked to Cortical Up-Down States during Sleep//Cereb. Cortex. 2012. Vol. 22. № 2. P. 426-435.
- Follett K.A., Gebhart G.F. Modulation of cortical evoked potentials by stimulation of nucleus raphe magnus in rats//J. Neurophysiol. 1992. Vol. 67. № 4. P. 820-828.
- Juckel G., Hegerl U., Giegling I. et al. Association of 5-HT1B receptor polymorphisms with the loudness dependence of auditoryevoked potentials in a community-based sample of healthy volunteers//Am. J. Med. Gene.t B. Neuropsychiatr. Genet. 2008. Vol. 147B. № 4. P. 454-458.
- Wenk G.L. The nucleus basalis magnocellularis cholinergic system: one hundred years of progress//Neurobiol. Learn. Mem. 1997. Vol. 67. № 2. P. 85-95.
- Metherate R., Cox C.L., Ashe J.H. Cellular bases of neocortical activation: modulation of neural oscillations by the nucleus basalisand endogenous acetylcholine//J. Neurosci. 1992. Vol. 12. № 12. P. 4701-411.
