Свето-температурная характеристика СО2-газообмена растений гречихи обыкновенной

Автор: Дроздов С.Н., Курец В.К., Лаханов А.П., Наполова Г.В., Попов Э.Г., Таланов А.В., Холопцева Е.С.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Биология продукционного процесса
Статья в выпуске: 5 т.39, 2004 года.
Бесплатный доступ
В регулируемых условиях планируемого многофакторного эксперимента изучали влияние облученности и температуры воздуха на интенсивность нетто-фотосинтеза и темнового дыхания интактных растений гречихи разных видов и морфотипов в фазе трех листьев. На основании регрессионного анализа экспериментальных данных оценивали взаимосвязь между CO2-газообменом растений и факторами среды.
Короткий адрес: https://sciup.org/142133009
IDR: 142133009 | УДК: 633.12:581.132.036:58.02
Light-temperature characteristic of CO2-gaseous exchange in buckwheat plants
At the regulated conditions of polyfactor experiment the authors studied the influence of sunniness and temperature of air on intensity of net-photosynthesis and dark respiration in different species of plants and morphotypes of buckwheat at the three leaves stage. On the basis of regression analysis of experimental data the correlation between CO2-gaseous exchange in plants and of the environmental factors was estimated. The data obtained suggest about the divergence of Fagopyrum genus and intraspecial genetic diversity of Fagopyrum esculentum ssp. esculentum. During evolution, cultivation, ecological specialization and selection in buckwheat plants the intensity and productivity of photosynthesis, dark respiration, biological and seed productivity were raised significantly, that were due to the change of donor-acceptor relationship and first adaptive culture strategy - from competitive to ruderal type.
Текст научной статьи Свето-температурная характеристика СО2-газообмена растений гречихи обыкновенной
В задачу нашей работы входило изучение влияния облученности и температуры воздуха на интенсивность СО2-газообмена интактных растений гречихи различных видов, морфотипов и эколого-географического происхождения.
Методика. Объектом исследования служили следующие виды и подвиды гречихи, отражающие эволюцию вида: F. homotropicum — дикая форма (укороченные междоузлия и развитая зона ветвления, мелкие цветки, способность к самоопылению); F. esculentum ssp. ancestrale — подвид гречихи обыкновенной (перекрестное опыление, сильно развита зона ветвления); F. tataricum ssp. tataricum — окультуренный подвид гречихи татарской; F. esculentum ssp. esculentum — культурный подвид гречихи обыкновенной, представленный сортами различных морфотипов, отселекти-рованных в России, — Казанская 3, Дикуль, Скороспелая 86, Богатырь, Молва, Тройка, Баллада — и интродуцированными сортообразцами из других стран — Botansoba (Япония), Mankan Royal (Канада), М-822 (Северный Китай), Коричневая Лимадо (Центральный Китай).
Посев проводили проросшими калиброванными семенами по 7-8 шт. на один сосуд объемом 0,75 л. Растения выращивали в камере искусственного климата в песчаной культуре при 14-часовом фотопериоде, облученности 200-220 Вт/м2 и температуре воздуха 8/15 оС (день/ночь). Для полива использовали питательный раствор (pH 6,5-6,7), в который добавляли микроэлементы; нестандартные растения удаляли. Сосуды с растениями в фазе трех настоящих листьев поочередно помещали в камеру для исследования СО 2 -газообмена. Многофакторный эксперимент осуществляли по модифицированному плану Хартли (6) по 10 вариантам опыта (I-X), экспозиция в каждом из которых составляла 50-60 мин. Интенсивность газообмена рассчитывали на единицу сухой массы целого растения по разности концентраций СО2 на входе и выходе ассимиляционной камеры.
* Работа выполнена при поддержке РФФИ (грант ¹ 02-04-48461).
Темновое дыхание регистрировали в ночное время при изменении температуры со скоростью 5,5 оС/ч. Величину гросс-фотосинтеза и составляющие дыхания определяли расчетным способом по стандартной методике (7, 8). Статистическую обработку данных проводили методом регрессионного анализа (9). Зависимость нет-то-фотосинтеза от дозы облученности и температуры воздуха оценивали по следующему уравнению: Pn = bo + b1E + b2T + b3ET + b4E2 + b5T2, где Рn — интенсивность видимого фотосинтеза, мг/(г⋅ч); Е — облученность, Вт/м2; Т — температура воздуха, оС; b0-b5 — коэффициенты, рассчитанные по данным эксперимента. Среднеквадратичная ошибка моделей не превышала 10 %.
Результаты. Показатели интенсивности нетто-фотосинтеза растений гречихи разных видов, подвидов и сортов и соответствующие им значения облученности и температуры воздуха представлены в таблице 1. В фазе трех настоящих листьев наиболее высокой интенсивностью нетто-фотосинтеза характеризовались растения сортов культурного подвида гречихи Казанская 3, Дикуль, Баллада, Коричневая Лимадо — соответственно 32,5; 30,9; 28,8 и 28,0 мг/(г ⋅ ч) (табл . 2). Наибольшая интенсивность темнового дыхания у растений этих сортов отмечена при температуре, соответствующей максимальной интенсивности фотосинтеза; потенциальный максимум видимого фотосинтеза у исследуемых образцов выявлен при облученности 380-600 Вт/м2 и температуре воздуха 10-21 оС. Так, у растений детерминантного сорта Дикуль максимальная интенсивность фотосинтеза наблюдалась при облученности около 430 Вт/м2 и температуре воздуха 21 оС, индетерминантного позднеспелого сорта Mankan Royal (Канада) — соответственно 650 Вт/м2 и 14 оС, дикого вида F. homotropicum — 600 Вт/м2 и 14 оС.
Однако наиболее важное значение имеют не конкретные параметры окружающей среды, при которых интенсивность фотосинтеза достигает максимума, что редко встречается в естественных ценозах, а условия, обеспечивающие оптимальную фотосинтетическую активность (10). Температурный диапазон оптимума у растений сортов Казанская 3 (фасциированный стебель) и Баллада (ограниченноветвящийся стебель) составлял соответственно 8,0-30,0 и 7,0-27,0 оС, Дикуль (детерминантный) и Тройка (индетерминантный, небольшое число узлов в зоне побегообразования, продолжительный период цветения) — 12,0-21,5 и 11,0-24,0 оС. Наиболее теплолюбивыми оказались растения сортов Дикуль и Тройка, так как нижняя температурная граница области оптимума видимого фотосинтеза располагалась соответственно около 12,0 и 11,0 оС, жароустойчивыми — сорта Казанская 3 (верхняя температурная граница достигала 30,0 оС). Эти показатели соответствуют условиям зоны возделывания этих сортов: на юге европейской части России (Дикуль) и в Среднем Поволжье (Казанская 3).
Область оптимума нетто-фотосинтеза у ~ 1/2 исследованных сортов достигалась при умеренной температуре воздуха — от 4-5 до 19-23 оС. У наиболее светолюбивых растений (сорта Mankan Royal и Казанская 3, дикая форма F. homotropicum ) оптимум нетто-фотосинтеза в фазе трех настоящих листьев обеспечивался при дозе облученности соответственно не ниже 410, выше 380 и 390 Вт/м2, у менее светолюбивых образцов (представители F. esculentum ssp. ancestrale и сорт Молва) — не ниже 260 и 270 Вт/м2. Границы оптимума нетто-фотосинтеза по облученности и температуре воздуха одного из исследованных сортов гречихи (Богатырь), полученные на основе модели, приведены в качестве примера на рисунке 1.
-
1. Зависимость интенсивности нетто-фотосинтеза интактных растений гречихи разных видов, подвидов и сортов в фазе трех настоящих листьев от облученности и температуры воздуха
-
2. Cвето-температурная характеристика CO 2 -газообмена интактных растений гречихи разных видов, подвидов и сортов в фазе трех настоящих листьев
-
3. Морфобиологические характеристики, продуктивность фотосинтеза и урожай растений гречихи разных видов, подвидов и сортов в зависимости от продолжительности вегетационного периода
Вид, подвид, сорт (происхождение)
Продолжительность вегетационного периода, сут
Высота растений, см
Урожай, г/раст.
Масса семян, г/раст.
Чистая продуктивность фотосинтеза, г/(м2 ⋅ сут)
биологи
ческая
хозяйственная
F. tataricum ssp. tataricum
91
182
12,7
4,45
3,89
2,09
F. homotropicum (2 n )
116
132
14,0
2,22
4,25
0,54
F. esculentum ssp. ancestrale
110
126
15,0
2,67
4,63
0,65
F. esculentum ssp. esculentum
89
121
9,53
3,58
5,46
1,85
В том числе сорта:
Богатырь (Россия)
77
106
7,53
3,57
6,08
2,24
Баллада (Россия)
80
122
8,31
3,71
5,93
1,82
Скороспелая 86 (Россия)
75
88
7,83
3,65
7,28
2,62
Дикуль (Россия)
79
87
8,38
3,85
6,36
2,21
M-822 (Китай)
106
155
11,08
3,20
4,21
1,25
Botansoba (Япония)
108
149
10,17
2,49
3,97
1,16
Mankan Royal (Канада)
98
143
13,41
4,56
4,41
1,66
|
Вариант опыта |
Облученность, Вт/м2 |
Температура воздуха, оС |
Интенсивность нетто-фотосинтеза, мг СО 2 /г сухой массы ⋅ ч - 1 |
|||||||||||||
|
1 |
2 |
3 |
4 1 |
5 |
6 |
1 7 |
8 1 |
9 |
10 |
11 |
12 |
13 1 |
14 |
|||
|
I |
250 |
20 |
14,9 |
19,9 |
15,7 |
13,6 |
14,9 |
9,3 |
8,9 |
14,5 |
10,4 |
16,9 |
7,0 |
16,8 |
17,7 |
15,9 |
|
II |
0 |
20 |
–6,1 |
–6,8 |
–3,9 |
–1,9 |
–3,9 |
–3,9 |
–5,4 |
–4,4 |
–3,2 |
–5,6 |
–6,0 |
–5,3 |
–7,7 |
–4,4 |
|
III |
400 |
33 |
19,9 |
20,6 |
13,1 |
11,7 |
15,1 |
5,8 |
0,7 |
15,1 |
10,1 |
17,3 |
14,6 |
10,1 |
16,5 |
16,0 |
|
IV |
0 |
33 |
–8,8 |
–11,4 |
–8,0 |
–5,2 |
–7,1 |
–6,1 |
–8,4 |
–7,4 |
–6,4 |
–8,8 |
–11,2 |
–9,4 |
–11,6 |
–7,8 |
|
V |
100 |
20 |
7,6 |
9,2 |
7,1 |
8,0 |
9,5 |
4,4 |
4,2 |
8,8 |
3,8 |
11,7 |
6,8 |
4,8 |
6,1 |
8,7 |
|
VI |
100 |
7 |
7,4 |
10,7 |
9,0 |
8,9 |
11,3 |
3,9 |
6,9 |
10,2 |
3,2 |
14,1 |
8,0 |
6,0 |
9,2 |
10,3 |
|
VII |
0 |
7 |
–2,0 |
–1,8 |
–1,5 |
0 |
0 |
0 |
–1,2 |
–1,4 |
–2,3 |
–0,4 |
–3,6 |
–3,6 |
–3,2 |
–0,3 |
|
VIII |
400 |
20 |
23,6 |
25,2 |
19,6 |
17,1 |
18,3 |
13,4 |
15,2 |
21,5 |
17,9 |
26,9 |
21,0 |
20,0 |
22,6 |
22,4 |
|
IX |
250 |
33 |
15,1 |
15,1 |
12,7 |
11,0 |
12,6 |
5,6 |
6,1 |
12,9 |
10,7 |
14,1 |
11,0 |
9,2 |
11,0 |
12,8 |
|
X |
250 |
7 |
11,9 |
16,0 |
14,0 |
13,2 |
14,7 |
9,5 |
13,7 |
18,4 |
9,2 |
24,9 |
15,2 |
13,0 |
17,4 |
16,2 |
П р и м е ч а н и е. 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14 — соответственно сорта и морфотипы Казанская 3, Дикуль, Скороспелая 86, Богатырь, Botansoba, F. homotropicum (2 n ), F. esculentum ssp. ancestrale , Молва,
Mankan Royal, F. tataricum ssp. tataricum , M-822, Тройка, Коричневая Лимадо, Баллада.
|
Вид, подвид, сорт (морфотип) |
р n max |
P n max , мг/(г ⋅ ч) |
P n opt |
р_ g max , мг/(г ⋅ ч) |
R d при температуре P n max , мг/(г ⋅ ч) |
R g max , мг/(г ⋅ ч) |
K max , отн. ед. |
о T max K , |
||
|
Е , Вт/м2 1 |
T , оС |
Е , Вт/м2 |
Т, оС |
|||||||
|
F. tataricum ssp. tataricum |
400 |
10,0 |
18,60 |
>280 |
4,0-20,0 |
20,59 |
1,99 |
2,10 |
1,75 |
10,0 |
|
F. homotroptcum (2 n ) |
600 |
14,0 |
15,90 |
>390 |
4,0-19,0 |
16,73 |
0,84 |
1,10 |
1,50 |
17,5 |
|
F. esculentum ssp. ancestrale |
380 |
10,0 |
18,00 |
>260 |
4,0-23,0 |
18,74 |
0,75 |
0,85 |
0,85 |
16,0 |
|
F. esculentum ssp . escularicum |
380 |
10,0 |
21,09 |
270 |
4,0-27,0 |
24,58 |
3,49 |
3,60 |
2,60 |
10,0 |
|
В том числе сорта: |
||||||||||
|
Богатырь (индетерминантный) |
420 |
15,0 |
17,60 |
>320 |
8,0-24,0 |
19,31 |
1,71 |
2,50 |
2,20 |
17,5 |
|
Скороспелая 86 (индетерминантный) |
400 |
17,0 |
19,80 |
>310 |
10,0-25,0 |
22,85 |
3,06 |
3,10 |
2,00 |
17,0 |
|
Баллада (ограниченноветвящийся) |
480 |
15,0 |
24,20 |
>370 |
7,0-27,0 |
28,84 |
4,41 |
4,00 |
4,20 |
10,0 |
|
Молва (ограниченноветвящийся) |
380 |
10,5 |
16,20 |
>270 |
4,0-21,0 |
17,35 |
4,64 |
6,20 |
1,45 |
10,0 |
|
Тройка (индетерминантный) |
420 |
18,0 |
20,20 |
>300 |
11,0-24,0 |
24,61 |
1,15 |
1,25 |
2,70 |
10,0 |
|
Дикуль (детерминантный) |
430 |
21,0 |
25,30 |
>320 |
12,0-21,5 |
30,91 |
5,62 |
4,20 |
2,50 |
13,0 |
|
Казанская 3 (фасциированный) |
600 |
19,0 |
25,40 |
>380 |
8,0-30,0 |
32,47 |
7,08 |
6,80 |
3,80 |
10,0 |
|
M-822 (индетерминантный) |
430 |
10,0 |
20,50 |
>300 |
4,0-19,0 |
22,90 |
2,41 |
3,00 |
2,20 |
10,0 |
|
Коричневая Лимадо (индетерминантный) |
470 |
17,0 |
23,20 |
>320 |
8,0-27,0 |
28,00 |
4,80 |
5,10 |
3,50 |
10,0 |
|
Botansoba (индетерминантный) |
380 |
18,0 |
18,30 |
>290 |
9,0-28,0 |
19,86 |
1,57 |
2,00 |
2,70 |
17,5 |
|
Mankan Royal (индетерминантный) |
650 |
14,0 |
21,30 |
>410 |
5,0-23,0 |
23,24 |
1,94 |
1,50 |
1,35 |
10,0 |
П р и м е ч а н и е. P n max и P g max — соответственно максимальная интенсивность нетто-фотосинтеза и гросс-фотосинтеза, E и T — значения облученности и температуры воздуха, соответствующие максимуму ( P n max ) и оптимуму ( P n opt ) нетто-фотосинтеза, R d — интенсивность темнового дыхания при температуре, соответствующей максимальной интенсивности нетто-фотосинтеза, R g max и T max K — максимальные значения дыхания роста и температуры, при которых соотношение между интенсивностью дыхания роста и дыхания поддержания ( К ) достигает наибольшей величины.
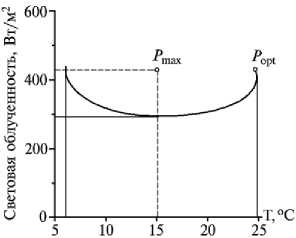
Рис. 1. Область оптимума нетто-фотосинтеза по облученности и температуре воздуха у интактных растений гречихи сорта Богатырь в фазе трех настоящих листьев: P max и P opt — соответственно максимум и оптимум нетто-фотосинтеза, составляющие 17,6 и 15,4 мг СО 2 /г сухой массы ⋅ ч - 1.
Следовательно, род Fagopyrum является достаточно гетерогенным по показателям максимальной интенсивности СО2-газообмена и соответствующим им значениям облученности и температуры воздуха. В процессе эволюции, естественного и искусственного отбора, окультуривания, селекции и экологической специализации у растений гречихи обыкновенной и особенно культурного подвида (F. esculentum ssp. esculentum) существенно повысилась интенсивность газооб- мена.
Весьма важными для экологофизиологической характеристики сорта являются компоненты темнового дыхания: дыхание роста и дыхание поддержания, сопряженные с интенсивностью роста и устойчивостью растений к экстремальным условиям внешней среды; соотношение между этими показателями (К), характери- зующее долю затрат на рост; температура, при которой К достигает наибольшей величины (TmaxK) (см. табл. 2, рис. 2). Максимальное количество ассимилятов расходуется в ночное время на прирост биомассы растения при температуре, соответствующей Кmax (11). У растений дикой формы F. homotropicum (2n), сортов Богатырь, Bo-tansoba, Скороспелая 86, Дикуль и подвида F. esculentum ssp. ancestrale температура Кmax составляла соответственно 17,5, 17,5, 17,5, 17,0, 13,0 и 16,0 оC; у представителей остальных подвидов, видов и сортов —
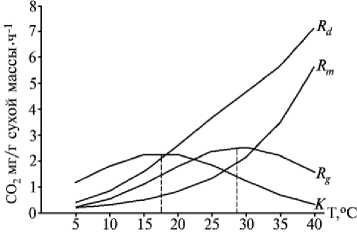
Рис. 2. Интенсивность темнового дыхания ( R d ), дыхания роста ( R g ) , дыхания поддержания ( R m ) и соотношение между интенсивностью дыхания роста и дыхания поддержания ( K, отн. ед.) интактных растений гречихи сорта Богатырь в фазе трех настоящих листьев при различной температуре воздуха.
10,0 оС. Температурный минимум дыхания роста у исследованных образцов гречихи располагался в диапазоне 4,5-0,5 оС. Наиболее устойчивыми оказались растения окультуренного подвида F. tataricum ssp. tataricum (дыхание роста наблюдалось при 0,5 оС), менее устойчивыми — сорта Botan-soba (Япония) (рост прекращался при 4,5 оС). Интенсивность дыхания интактных растений возрастала при повышении температуры воздуха в пределах 5-40 оС (см. рис. 2).
Если рассматривать интенсивность темнового дыхания и соотношение между интенсивностью дыхания роста и дыхания поддержания растений гречихи разных видов и подвидов в эволюционном аспекте, то можно сделать вывод о повышении этих показателей по мере окультуривания и создания более скороспелых сортов. Большинство сортов культурного подвида гречихи обыкновенной, отселектированных в России, характеризовались более коротким вегетационным периодом, высокой семенной продуктивностью и интенсивностью темнового дыхания по сравнению с позднеспелыми сортами, интродуцированными из Японии, Китая и Канады; при этом их температурные характеристики существенно не менялись.
Температура воздуха и доза облученности, при которых достигались максимальная и оптимальная интенсивность как фотосинтеза, так и дыхания роста, в целом соответствуют эколого-географическим условиям обитания диких видов и возделывания культурных сортов гречихи обыкновенной, формирования у них биологической и хозяйственной продуктивности (табл. 3).
Наиболее высокая чистая продуктивность фотосинтеза отмечена у растений сорта Скороспелая 86, соотношение между максимальной интенсивностью нетто-фотосинтеза и темнового дыхания которых составляло ~ 6,5. У растений сортов Ди-куль и Баллада (лидеры по нетто-фотосинтезу) этот показатель не превышал соответственно 4,3 и 1,2, что свидетельствует о менее рациональном использовании ассимилятов на формирование хозяйственно полезного урожая.
Таким образом, полученные нами данные свидетельствуют об относительно широкой гетерогенности рода Fagopyrum и внутривидовом генетическом разнообразии Fagopyrum esculentum ssp. esculentum по показателям облученности и температуры воздуха, обеспечивающим максимальную и оптимальную интенсивность нетто-фотосинтеза и темнового дыхания, в том числе дыхания роста и дыхания поддержания, обусловливающих формирование биологической и семенной продуктивности в естественных условиях обитания и возделывания растений. В целом это соответствует изменению первичной адаптивной стратегии культуры — от конкурентного (C), через промежуточный (C/R) к рудеральному (R) типу, произошедшему в процессе эволюции, окультуривания и селекции (12).
Л И Т Е Р А Т У Р А
-
1. К р о т о в А.С. Культурная флора СССР. Крупяные культуры, 1975, 3: 3-118.
-
2. Л а в р и н е н к о Г.Т. Производство гречихи в современных экономических условиях. Зерновые культуры, 1993, 2: 2-6.
-
3. Л а х а н о в А.П. Физиологические аспекты селекции гречихи. Аграрная наука, 1997, 3: 25-27.
-
4. З а д о р и н А.Д. Проблемы научного обеспечения производства зернобобовых и крупяных культур.
-
5. O h n i s h i O. Discovery of wild ancestor of common buckwheat. Fagopyrum, 1991, 11: 5-10.
-
6. К у р е ц В.К., П о п о в Э.Г. Моделирование продуктивности и устойчивости растений. Л., 1979.
-
7. Т о р н л и Дж.Г.М. Математические модели в физиологии растений. Киев, 1982.
-
8. Г о л о в к о Т.К. Дыхание растений. СПб, 1999.
-
9. Н а л и м о в В.В., Ч е р н о в Н.А. Статистические методы планирования экстремальных экспериментов. М., 1965.
-
10. Л а р х е р В. Экология растений. М., 1978.
-
11. П о п о в Э.Г., Т а л а н о в А.В., К у р е ц В.К. и др. Влияние температуры на суточную динамику СО 2 -
газообмена интактного растения огурца. Физиол. раст., 2003, 50, 2: 200-204.
-
12. Л а х а н о в А.П. Морфофизиологические аспекты эволюционного становления гречихи культурной ( Fagopyrum esculentum ). Физиол. раст., 2001, 48, 5: 670-676.
Поступила в редакцию 16 июня 2003 года
Институт биологии Карельского научного центра РАН, 185610, Петрозаводск, ул. Пушкинская, 11; Всероссийский НИИ зернобобовых и крупных культур, Орел;
Вест. РАСХН, 2002, 5: 43-48.
Орловский государственный аграрный университет
LIGHT-TEMPERATURE CHARACTERISTIC OFCO2-GASEOUS EXCHANGE IN BUCKWHEAT PLANTS
S.N. Drozdov, B.K. Kurets, A.P. Lakhanov, G.V. Napolova, E.G. Popov, A.V. Talanov, E.S. Kholoptseva
S u m m a r y
At the regulated conditions of polyfactor experiment the authors studied the influence of sunniness and temperature of air on intensity of net-photosynthesis and dark respiration in different species of plants and mor-photypes of buckwheat at the three leaves stage. On the basis of regression analysis of experimental data the correlation between CO 2 -gaseous exchange in plants and of the environmental factors was estimated. The data obtained suggest about the divergence of Fagopyrum genus and intraspecial genetic diversity of Fagopyrum esculentum ssp. esculentum. During evolution, cultivation, ecological specialization and selection in buckwheat plants the intensity and productivity of photosynthesis, dark respiration, biological and seed productivity were raised significantly, that were due to the change of donor-acceptor relationship and first adaptive culture strategy — from competitive to ruderal type.
Новые книги
К о л о с о в а Е.Н. Технологические приемы эффективного возделывания гречихи. Курск: изд-во КГСХА, 2003, 121 с.
В книге изложены научные основы возделывания растений гречихи в условиях Центрально-Черноземной зоны — важнейшего аграрного экономического района страны. Дана оценка влияния способов и сроков посева, нормы высева семян на урожайность растений. Показано, что максимальная продуктивность растений гречихи может быть достигнута при посеве широкорядным способом (ширина междурядий 45 см) в оптимально ранние сроки при норме высева 2 млн всхожих семян на 1 га. Обоснована целесообразность окучивания и совмещения двух сроков широкорядных посевов культуры на одной площади. Предложен новый полосный способ возделывания растений гречихи совместно с озимой рожью, предусматривающий проведение сеникации 10 % раствором аммиачной селитры с целью ускорения естественного созревания гречихи, что обеспечивает сокращение вегетационного периода, проведение однообразной уборки урожая и сокращение потерь зерна. Представлены рекомендации по совместному возделыванию растений гречихи и проса широкорядным способом чередующимися рядками при половинной норме высева семян. Проанализирована возможность использования сидеральных культур в паровом поле севооборота и пожнивно непосредственно перед посевом гречихи с целью сокращения дозы минеральных удобрений. Описаны основные принципы возделывания растений гречихи в био-логизированных севооборотах. Рассматриваются биологические факторы повышения продуктивности растений этой культуры.
Диагностика и методы учета злаковых тлей. Метод. реком. /Сост. М.Н. Брим, Е.Е. Радченко. М.: ФГНУ «Росинформагротех», 2002, 24 с.
В рекомендациях на основании данных литературы и собственных исследований авторов представлено подробное описание биологии четырех видов тлей, являющихся опасными вредителями зерновых культур. Приведены методы оценки (в баллах) заселенности растений тлями в полевых условиях, а также методика учета численности черемуховой тли в период зимовки яиц и ранней весной. Показано, что оценка плотности популяции этого вида на первичном растении-хозяине позволяет прогнозировать дальнейшее развитие вредителя и потенциальную опасность последнего как переносчика вирусных заболеваний злаковых культур. Рассматриваются факторы, оказывающие влияние на вспышки массового размножения злаковых тлей.
