Свето-температурные характеристики фотосинтеза у двух видов амаранта

Автор: Дроздов С.Н., Холопцева Е.С., Коломейченко В.В.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Фотосинтез и продуктивность: роль температурного фактора
Статья в выпуске: 5 т.49, 2014 года.
Бесплатный доступ
Из многочисленных видов рода Amaranthus L. 12 окультурены и используются как овощные, зерновые, кормовые, лекарственные и декоративные растения. Один из сдерживающих факторов интродукции амаранта в нашей стране - его недостаточная эколого-физиоло-гическая изученность, что препятствует селекционной работе при выведении сортов, особенно для северных регионов с низкими температурами в весенний период вегетации. В регулируемых условиях среды в двухфакторном планируемом эксперименте мы изучили влияние интенсивности света и температуры на нетто-фотосинтез у интактных растений амаранта багряного ( Amaranthus cruenthus L.) сорта Султан и амаранта овощного ( A. hypochondriacus L.) сорта Крепыш. В фазу 6-7 листьев часть растений в течение 3 сут подвергали закаливанию при 8 °С для амаранта овощного и 10 °С - для амаранта багряного. Концентрацию СО 2 измеряли на оптико-акустическом инфракрасном газоанализаторе Infrаlуt-IV («SAXON Junkalor GmbH», Германия), включенном по дифференциальной схеме. Были получены нелинейные уравнения (модель), описывающие связи СО 2-газообмена с факторами среды: NP = a 0 + a 1E + a 2T + a 3ET + a 4E 2 + a 5T 2, где NP - интенсивность видимого фотосинтеза, мг СО 2/(г?ч); Е - освещенность, Вт/м 2; Т - температура воздуха, °С; a 0-a 5 - коэффициенты, вычисленные по экспериментальным данным. Достоверность уравнений проверяли по коэффициенту множественной детерминации (R 2 = 85-94) и F -критерию Фишера (F = 4,1-5,6) при уровне значимости р = 0,05. Полученная регрессионная модель дает возможность оценить величину видимого фотосинтеза в конкретную фазу развития растений для каждого сорта при разных условиях и рассчитать сочетания значений факторов среды, обеспечивающих достижение соответствующей интенсивности фотосинтеза, в том числе при наличии лимитирующего фактора. С их помощью мы определили освещенность и температуру, которые обеспечивают потенциальные максимум и оптимум нетто-фотосинтеза растений при естественном содержании СО 2 в воздухе. Потенциальный максимум нетто-фотосинтеза у изученных сортов составлял 37-38 мг СО 2/(г?ч). Сорт Султан проявил себя более свето- и теплолюбивым, а сорт Крепыш - холодоустойчивым. Диапазон оптимума фотосинтеза по температуре и освещенности у сорта Султан находился в пределах 26,7-47,0 °С и 335-580 Вт/м 2. У сорта Крепыш диапазон оптимума составлял соответственно 23,5-39,6 °С и 284-501 Вт/м 2. Закаливание растений повышало их терморезистентность, значительно снижало максимум нетто-фотосинтеза и приводило к сужению границ значений освещенности и температуры, необходимых для достижения его оптимальных значений.
Планируемый многофакторный эксперимент, нетто-фотосинтез, свето-температурная характеристика, со 2-обмен, со 2-exchange
Короткий адрес: https://sciup.org/142133546
IDR: 142133546 | УДК: 633:582.663:574.24:581.132
Light and temperature parameters to optimize photosynthesis in two Amaranthus L. species
Among the numerous taxonomic group of Amaranthus L., 12 species are cultivated as vegetable, cereal and fodder crops, medicinal and ornamental plants. A limitation for their introduction in Russia results particularly from lack of data on the physiological and ecological parameters of plant growth and development, which prevents the effective breeding investigations, especially with a view to create varieties for cultivation in northern regions, where the temperatures during spring vegetation are low. In controlled conditions of two factorial preplanned experiment we studied the effect of light intensity and temperature to net photosynthesis in intact Amaranthus cruenthus L. plants (Sultan variety) and A. hypochondriacus L. plants (Krepysh variety). At 6-7 leaf phase, the experimental plants were subjected to hardening for 3 days at 8 °С and 10 °С (for A. hypochondriacus and A. cruenthus, respectively). To evaluate the CO 2 concentration, the infrared gas analyzer Infrаlуt-IV (SAXON Junkalor GmbH, Germany) was used according to differential scheme. To describe the relationship between CO 2 exchange and external factors, the nonlinear equations (a model) were obtained: NP = a 0 + a 1E + a 2T + a 3ET + a 4E 2 + a 5T 2, with NP as the intensity of observed photosynthesis, mg СО 2/(g?h); Е as illumination, W/m 2; Т as air temperature, °С; a 0-a 5 as the coefficients calculated basing on the experimental data. A reliability of the equations was verified by the multiple determination index (R 2 = 85-94) and F-Fisher test (F = 4.1-5.6) at р = 0.05. This model can be applied for estimating photosynthetic activity at definite vegetation phase for each cultivar under different conditions and predicting parameters necessary to achieve definite net photosynthesis even under limiting factors. Thus, using multiple regression analysis, the equations were obtained, which allow estimating favorable combinations of the light intensity and temperature for maximal and optimal net photosynthesis at natural CO 2 concentration in the air. In both examined varieties, the net photosynthesis value of 37-38 мг СО 2/(g?h) was registered as a potential maximum. Sultan variety was more light- and heat-loving, while Krepysh variety demonstrated more cold-resistance. The photosynthetic optimum was provided within the limits of 26.7-47.0 °С and 335-580 W/m 2 for Sultan variety and at 23.5-39.6 °С and 284-501 W/m 2 for Krepysh variety. Plant hardening increased their tolerance to low temperature, decreased significantly the peak of net photosynthesis and led to a narrowing the range of light intensity and temperature, necessary to achieve the optimal net photosynthetic parameter.
Текст научной статьи Свето-температурные характеристики фотосинтеза у двух видов амаранта
Амарант-щирица — древняя культура, которая в течение 8 тыс. лет кормила население американского континента. Продукты из зерен амаранта очень питательны, вкусом и ароматом напоминают орехи. Из многочисленных видов рода Amaranthus 12 окультурены и используются как овощные, зерновые, кормовые, лекарственные и декоративные растения.
В России первые исследования амаранта проведены в 1930-х годах по инициативе академика Н.И. Вавилова, но были приостановлены и возобновлены только во второй половине XX века профессором И.М. Магомедовым. К настоящему времени эта уникальная культура нашла широкое распространение в Северной и Южной Америке и Азии (1-3). Один из сдерживающих факторов интродукции амаранта в нашей стране — его недостаточная эколого-физиологическая изученность, что препятствует селекционной работе при выведении сортов, особенно для северных регионов с низкими температурами в весенний период вегетации (4).
Возрастающая роль экологических показателей в решении практических вопросов требует их перевода из качественного описания в количе-
* Исследования выполнены на оборудовании ЦКП НО Института биологии Карелького НЦ РАН.
ственное (5, 6). Для оценки реакции растения на действие внешней среды в активном многофакторном планируемом эксперименте нужен лабильный интегральный показатель, характеризующий первичный процесс продуктивности (СО2-обмен), быстро реагирующий на изменения условий среды и доступный для регистрации без контакта с растением, дистанционно и непрерывно. Одним из методов получения экофизиологической характеристики растений служит определение параметров основных факторов внешней среды, которые обеспечивают достижение оптимального нетто-фотосинтеза (7, 8). При этом необходимо учитывать зональность их влияния на пойкилотермные организмы, поскольку изменение значений, соответствующее переходу из зоны с одним характером действия фактора в зону с другим, затрагивает геном и приводит к изменению направленности обмена веществ (9).
Мы провели многофакторный планируемый эксперимент с целью изучения влияния интенсивности света и температуры на нетто-фотосинтез.
Методика. В эксперименте использовали амарант багряный ( Ama-ranthus cruenthus L.) сорта Султан и амарант овощной ( A . hypochondria-cus L.) сорта Крепыш, различающиеся по холодоустойчивости и хозяйственному назначению.
Растения выращивали в контролируемых факторостатных условиях среды в пластиковых сосудах объемом 0,5 л с песчаным субстратом. Калиброванные проросшие семена высаживали по 12-15 шт. на сосуд. Растения выращивали под люминесцентными лампами, обеспечивающими освещенность 110-120 Вт/м2, при 14-часовом фотопериоде и температуре воздуха 20/18 °С (день/ночь). Полив осуществляли ежедневно питательным раствором Кнопа, дополненным микроэлементами с pH 6,2-6,5. Отстающие в росте и имеющие видимые отклонения растения выбраковывали, оставляя в сосуде по 10 шт.
В фазу развития 6-7 листа часть растений в течение 3 сут подвергали закаливанию при ранее установленных для исследуемых сортов положительных температурах: 8 °С — для амаранта овощного, 10 °С — для амаранта багряного. Изменение холодоустойчивости в процессе заливания определяли методом ЛТ50 по температуре, вызывающей гибель 50 % клеток листовых высечек, после 40-минутного промораживания в электрическом микротермостате ТЖР-02/02 («Интерм», Россия) при последовательном изменении температур с шагом 0,4 °С (10). За жизнеспособностью клеток следили с помощью светового микроскопа ЛОМО МИК-МЕД-2 (Россия).
Далее сосуды с контрольными и закаленными растениями поочередно помещали в установку (11) для исследования СО2-газообмена при освещенности 0-600 Вт/м2 и температуре 5-40 °С, где проводили двухфакторный эксперимент по трехуровневому плану (12) при сочетании значений температуры (10, 20 и 30 °С) и освещенности (150, 250 и 350 Вт/м2) на девяти ступенях в 3 повторностях.
Концентрацию СО2 определяли в оптико-акустическом инфракрасном газоанализаторе Infrа1уt-IV («SAXON Junkalor GmbH», Германия), включенном по дифференциальной схеме. После 40-60-минутной выдержки на каждой ступени газообмен растения оценивали по разности концентраций СО2 на входе и выходе в ассимиляционной камере установки с учетом скорости поступления наружного воздуха, пересчитывая на единицу сухой массы целых растений.
Экспериментальные данные обрабатывали методом множественного регрессионного анализа с использованием пакета программ KyPlotSta- tistics v. 2.0 и Microsoft Excel 2000.
Результаты. Нами были получены нелинейные уравнения (модель), описывающие связи СО2-газообмена с факторами среды:
NP = a 0 + a 1 E + a 2 T + a g ET + a 4 E2 + a g T2, где NP — интенсивность видимого фотосинтеза, мг СО2/(г^ч); Е — освещенность, Вт/м2; Т — температура воздуха, °С; a0-a5 — коэффициенты, вычисленные по экспериментальным данным.
Достоверность уравнений проверяли по коэффициенту множественной детерминации (R2 = 85-94) и F -критерию Фишера (F = 4,1-5,6) при уровне значимости р = 0,05.
Полученная регрессионная модель дает возможность оценить величину видимого фотосинтеза в конкретную фазу развития растений для каждого сорта при разных условиях и рассчитать сочетания значений факторов среды, обеспечивающих достижение соответствующей интенсивности фотосинтеза, в том числе при наличии лимитирующего фактора.
Мы определили максимальные значения видимого фотосинтеза при естественном содержании в воздухе СО2, а также области оптимума (90 % от максимума) и интенсивность света и температуры, необходимые для его достижения, у закаленных к холоду и незакаленных растений. Сочетание факторов среды, обеспечивающее достижение потенциального максимума видимого фотосинтеза, в естественных условиях наблюдается крайне редко. Интенсивный рост и развитие растений происходят в основном в фоновой зоне — зоне оптимума (13, 14), экологические характеристики которой и входят в понятие оптимума экологической ниши экотипа (7, 8). Однако в некоторых районах ночные температуры значительно снижаются, оказывая на растения, особенно теплолюбивые, закаливающий эффект и тем самым влияя на экологическую характеристику их СО2-газообмена.
Потенциальный максимум нетто-фотосинтеза у интактных растений амаранта сортов Султан и Крепыш составлял соответственно 37,5 и 37,8 мг СО2/(г^ч). При этом растения сорта Султан были более тепло- и светолюбивы по сравнению с представителями сорта Крепыш и имели значительно более широкую свето-температурную зону оптимума, особенно в области повышенных температур (табл.).
Показатели нетто-фотосинтеза и свето-температурные условия, обеспечивающие его достижение, у двух видов амаранта в фазу 6-7 листьев в условиях холодового закаливания (лабораторный опыт)
|
Вид, сорт |
Вариант |
Нетто-фотосинтез |
|||||
|
максимум |
оптимум |
||||||
|
NP |
1 Т 1 |
Е |
NP | |
Т I |
Е |
||
|
Амарант багряный Amaranthus |
Контроль |
37,5 |
36,8 |
459 |
> 33,8 |
26,7-47,0 |
335-580 |
|
cruenthus L., сорт Султан |
Закаливание |
15,8 |
24,1 |
303 |
> 14,3 |
17,7-30,5 |
218-388 |
|
Амарант овощной A . hypo- |
Контроль |
37,8 |
31,5 |
393 |
> 34,0 |
23,5-39,6 |
284-501 |
|
chondriacus L., сорт Крепыш |
Закаливание |
10,4 |
21,1 |
323 |
> 9,6 |
15,9-26,2 |
226-420 |
|
П р и м е ч а н и е. NP — интенсивность видимого фотосинтеза, мг |
СО2/(г - ч); Т |
— температура воздуха, |
|||||
|
°С; Е — освещенность, Вт/м2. |
|||||||
Максимум видимого фотосинтеза у сорта Султан достигался при температуре 36,8 °С и освещенности 459 Вт/м2, у сорта Крепыш — соответственно при 31,5 °С и 393 Вт/м2. Оба сорта заморозкоустойчивы и выдерживали кратковременное промораживание ниже -4 °С. Несколько более устойчивым оказался сорт Крепыш, у которого закаливание начиналось при более низкой положительной температуре.
Холодовое закаливание растений резко уменьшало интенсивность видимого фотосинтеза и значительно ослабляло влияние температуры, 98
особенно пониженной, на этот показатель (рис. 1). Как продемонстрировали ранее проведенные исследования (15, 16), снижение интенсивности видимого фотосинтеза у закаленных растений происходит в результате возрастания интенсивности дыхания, главным образом за счет составляющей поддержания.
Воздействие температуры 35 °C на закаленные растения менее теплолюбивого сорта Крепыш приводило к отрицательному газообмену независимо от интенсивности освещения. Температурные кривые у незакаленных растений обоих сортов имели выраженную куполообразную форму, а максимум нетто-фотосинтеза с повышением температуры наступал при большей освещенности (400 Вт/м2).
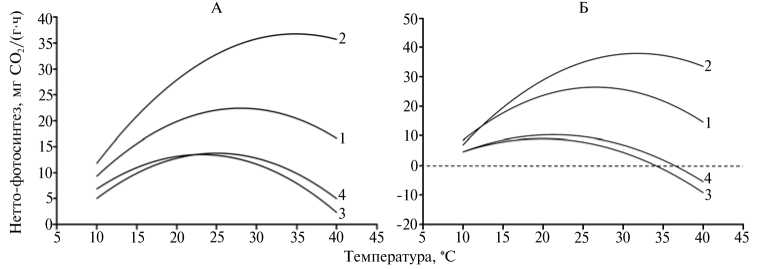
Рис. 1. Интенсивность нетто-фотосинтеза у интактных растений амаранта сортов Султан ( Ama-ranthus cruenthus L.) (А) и Крепыш ( A . hypochondriacus L.) (Б) в фазу 6-7 листьев в зависимости от температуры воздуха: 1 и 2 — контрольные растения при освещенности соответственно 200 и 400 Вт/м2; 3 и 4 — закаленные растения при освещенности соответственно 200 и 400 Вт/м2 (лабораторный опыт).
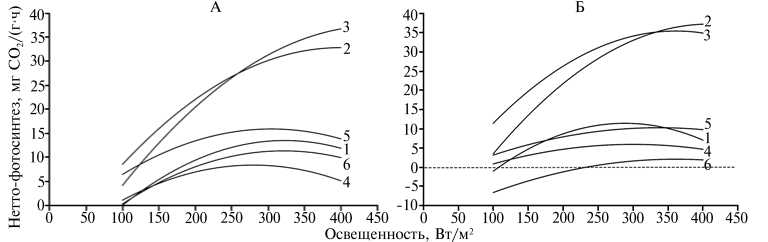
Рис. 2. Интенсивность нетто-фотосинтеза у интактных растений амаранта сортов Султан ( Ama-ranthus cruenthus L.) (А) и Крепыш ( A . hypochondriacus L.) (Б) в фазу 6-7 листьев в зависимости от освещенности: 1, 2 и 3 — контрольные растения при температуре соответственно 10, 25 и 35 ° C; 4, 5 и 6 — закаленные растения при температуре соответственно 10, 25 и 35 ° C (лабораторный опыт).
Световые кривые нетто-фотосинтеза, полученные в результате двухфакторного эксперимента в регулируемых условиях внешней среды, тоже стремились к куполообразной форме (рис. 2). Таким образом, выход видимого фотосинтеза на плато при усилении освещенности в однофакторных экспериментах (13, 17) происходит не по причине светового насыщения, как предполагают авторы этих работ, а в результате проявления лимитирующего фактора, возможно температуры. Против светового насыщения свидетельствуют данные о влиянии избыточной освещенности на появление синглетного кислорода и его разрушительном действии на фотосинтетический аппарат (18). В то же время световые кривые обоих сортов подтверждали необходимость увеличения освещенности при высоких температурах воздуха (см. рис. 2). Холодовое закаливание растений повышало их холодоустойчивость и изменяло степень влияния света, особенно в области пониженных температур.
Итак, потенциальный максимум нетто-фотосинтеза растений при естественном содержании в воздухе СО2 у амаранта багряного сорта Султан и амаранта овощного сорта Крепыш составляет 37-38 мг СО2/(г-ч), но наступает при разных свето-температурных условиях внешней среды. Сорт Султан значительно более свето- и теплолюбив. Его световая зона оптимума в фазу 6-7 листьев лежит в пределах 335-580 Вт/м2, температурная — 26,7-47,0 °С. В то же время у сорта Крепыш диапазон оптимума по освещенности составляет 284-501 Вт/м2, по температуре — 23,5-39,6 °С. У обоих сортов закаливание растений низкими положительными температурами (8-10 °С) повышает их терморезистентность, значительно снижает максимум нетто-фотосинтеза и приводит к сужению границ, в пределах которых освещенность и температура влияют на достижение его оптимальных значений.
Л И Т Е Р А Т У Р А
-
1. Магомедов И.М., Ярошевич М.И., Чернов И.А. Амарант: агротехника возделывания и использования. Л., 1980.
-
2. Же л е з о в А.В. Амарант — хлеб, зрелище и лекарство. Химия и жизнь, 2005, 6: 56-61.
-
3. Магомедов И.М. Физиологические основы конкурентно способности амаранта. Успехи современного естествознания, 2008, 5: 57-59.
-
4. Чиркова Т.В. Амарант — культура 21-го века. Соросовский образовательный журнал, 1996, 10: 23-27.
-
5. Урманцев Ю.А. Системный подход к проблеме устойчивости растений (на примере исследования зависимости содержания пигментов в листьях фасоли от одновременного действия на нее засухи и засоления). Физиология растений, 1976, 26(4): 762-777.
-
6. Курец В.К., Попов Э.Г. Статистическое моделирование системы связей растение— среда. Л., 1991.
-
7. Т о о м и н г Х.Г. Солнечная радиация и формирование урожая. Л., 1977.
-
8. Суворова Г.Г. Фотосинтез хвойных деревьев в условиях Сибири. Новосибирск, 2009.
-
9. Дроздов С.Н., Курец В.К. Некоторые аспекты экологической физиологии растений. Петрозаводск, 2003.
-
10. Дроздов С.Н., Курец В.К., Будыкина Н.П., Балагурова Н.И. Определение устойчивости растений к заморозкам. В сб.: Методы оценки устойчивости растений к неблагоприятным условиям среды. Л., 1976: 222-228.
-
11. Таланов А.В., Безденежных В.А., Хилков Н.И. Установка для исследования газообмена интактных растений. В сб.: Влияние факторов внешней среды и физиологически активных веществ на терморезистентность и продуктивность растений. Петрозаводск, 1982: 142-150.
-
12. Голикова Т.И., Панченко Л.А., Фридман М.З. Каталог планов второго порядка. М., 1974.
-
13. Лархер В. Экология растений. М., 1978.
-
14. Дроздов С.Н., Курец В.К., Титов А.Ф. Терморезистентность активно вегетирующих растений. Л., 1984.
-
15. Курец В.К., Дроздов С.Н., Попов Э.Г., Дембо Е.Д., Хилков Н.И., Трофимова С.А. Действие и последействие температуры на дыхание интактных растений. Физиология растений, 2003, 50(3): 81-87.
-
16. Дроздов С.Н., Сычева З.Ф., Попов Э.Г., Таланов А.А., Холопцева Е.С., Курец В.К. Роль дыхания в формировании терморезистентности растений. Физиология и биохимия растений, 2005, 37(1): 73-78.
-
17. Малкина И.С., Цельникер Ю.Л., Якшина А.М. Фотосинтез и дыхание подроста. М., 1970.
-
18. Мокроносов А.Т., Гавриленко В.Ф. Фотосинтез. Физиолого-экологические и биохимические аспекты. М., 1992.
1ФГБУН Институт биологии Карельского научного Поступила в редакцию центра РАН, 11 января 2012 года
LIGHT AND TEMPERATURE PARAMETERS TO OPTIMIZE PHOTOSYNTHESIS IN TWO Amaranthus L. SPECIES
-
|S.N. DrozdovV , E.S. Kholoptseva1, V.V. Kolomeichenko2
-
2 Orel State Agrarian University, 69, ul. Generala Rodina, Orel, 302019 Russia, e-mail borpli@rambler.ru
Study was carried out on the equipment of Ñenter for collective equipment use, Institute of Biology, the Karelian Scientific Center of Russian Academy of Sciences
Научные собрания

МЕЖДУНАРОДНАЯ НАУЧНО-ПРАКТИЧЕСКАЯ КОНФЕРЕНЦИЯ «СЕЛЕКЦИЯ НА УСТОЙЧИВОСТЬ К БИОТИЧЕСКИМ И АБИОТИЧЕСКИМ СТРЕССОРАМ У ОВОЩНЫХ КУЛЬТУР»
(11-13 августа 2014 года, Всероссийский НИИ селекции и семеноводства. Московская обл.)
Ïîñâÿùåíà 100-ëåòèþ àêàäåìèêà ÂÀÑÕÍÈË Ï.Ô. Ñîêîëà, ïàìÿòè ÷ëåíà-êîððåñïîíäåíòà ÀÍ ÐÌ Í.Í. Áàëàøîâîé, 100-ëåòèþ äîêòîðà ñåëüñêîõîçÿéñòâåííûõ íàóê Î.Â. Þðèíîé.
Основные направления конференции:
-
❖ Фитопатологический мониторинг и идентификация патогенов на овощных и дру-ãèõ ñåëüñêîõîçÿéñòâåííûõ êóëüòóðàõ
-
❖ Cелекция на устойчивость к биотическим и абиотическим стрессорам: генетические ðåñóðñû, èõ èñïîëüçîâàíèå; èììóíîãåíåòèêà, ìåòîäû
-
❖ Профилактика и защита овощных растений
-
❖ Адаптация к стрессам
Контакты и информация: , ;
