Связь заселяемости сортов роз паутинным клещом с элементами структуры куста в условиях применения хищного клеща фитосейулюса в теплицах

Автор: Моор В.В., Козлова Е.Г., Анисимов А.И.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Биометод контроля вредителей
Статья в выпуске: 3 т.58, 2023 года.
Бесплатный доступ
Сорта роз, выращиваемых на срез, различаются по степени затрат на защиту от вредителей, в первую очередь от обыкновенного паутинного клеща Tetranychus urticae Koch., для борьбы с которым проводят от 6-8 до 25 и более обработок акарицидами. Хищный клещ фитосейулюс Phytoseiulus persimilis A.-H. может быть использован в качестве альтернативы или дополнения к обработкам химическими препаратами. В настоящей работе благодаря многолетнему мониторингу развития вредителя на производственных площадях впервые дана количественная оценка связи отдельных морфологических особенностей структуры куста, а именно площади дольки сложного листа и общей площади листовой поверхности куста, со степенью развития паутинного клеща на разных сортах розы Rosa hybrida в присутствии акарифага фитосейулюса, то есть в системе триотрофа в защищенном грунте. На основе этого получены уравнения, позволяющие прогнозировать развитие вредителя и объемы внесения акарифага на разных сортах. Целью работы была многолетняя оценка заселяемости различных сортов роз обыкновенным паутинным клещом, установление ее связи с особенностями отдельных элементов структуры кустов, а также подбор математических моделей, позволяющих прогнозировать заселяемость роз этим вредителем и необходимые объемы использования хищного клеща фитосейулюса для борьбы с ним. Наблюдения за развитием обыкновенного паутинного клеща проводили в блочной стеклянной теплице ООО «Агролидер» (Ленинградская обл., Выборгский р-н) на растениях розы чайно-гибридной группы (18 сортов). Площадь теплицы составляла 45000 м2. Использовали балльную систему оценки заселенности роз паутинным клещом. Для этого теплица была разделена на участки. Каждый участок представлял собой отрезок двойного ряда кустов роз. Длина отрезка - 3,95 м, площадь - 8,02 м2. Обследование заключалось в визуальном осмотре растений на участке и присвоении балла заселенности от 1 до 5. Обследования проводили 2 раза в месяц, общее число учетов за год - не менее 24. Динамику заселенности роз паутинным клещом оценивали на протяжении 8 лет (2011-2018 годы). С 2011 года на отдельных сортах, а с 2012 - на всей площади комбината для борьбы с обыкновенным паутинным клещом применяли хищного клеща фитосейулюса Ph. persimilis , которого вносили, используя сплошной и локальный методы. Сплошное внесение от 3 до 10 особей/м2 по всей площади теплиц осуществляли 1-1,5 раза в месяц, в очаги - от 10 до 60 особей/куст еженедельно, до тех пор, пока продолжали появляться новые значительные очаги вредителя. Акарициды использовали только в тех случаях, когда заселенность роз T. urticae превышала 2,5 балла. Через 7 сут после первой обработки проводили повторную. Определяли следующие морфометрические показатели: число стеблей в верхней части куста (короне) и на всем кусте, длину продуктивных стеблей, число долек сложного листа, листьев на всем стебле и на 10 см стебля, листьев в короне куста и на всем кусте, площадь поверхности дольки и всего листа, площадь листовой поверхности короны и всего куста. Для оценки связи заселяемости отдельных сортов роз с особенностями элементов структуры их кустов использовали корреляционный анализ, а для ее математического описания - регрессионный (уравнения прямолинейной регрессии). Чтобы установить взаимосвязь параметров отдельных элементов структуры кустов роз с заселяемостью паутинным клещом, применяли двухфакторный дисперсионный анализ. При сравнении параметров регрессионных моделей, построенных по выборочным данным, использовали метод наименьших квадратов. Два наиболее контрастных по средней многолетней заселяемости сорта Brazil и Aqua различались в среднем в 17,8 раза. Остальные сорта можно было разделить на 6-8 групп, из которых наиболее контрастные различались в 5 раз. Сорта роз достоверно различались по средним показателям отдельных элементов структуры кустов: числу стеблей в короне и на всем кусте, долек у сложного листа, листьев на всем стебле и на 10 см стебля, листьев в короне и на всем кусте, дли
Сорта роз, куст, элементы структуры, производственные теплицы, заселенность вредителем, корреляционный анализ, регрессионный анализ, прогнозирование, уравнения прямолинейной регрессии
Короткий адрес: https://sciup.org/142238889
IDR: 142238889 | УДК: 582.734:635.918:632.7:632.937 | DOI: 10.15389/agrobiology.2023.3.458rus
Relationship of the rose varieties infestation level by spider mite with the bush structural elements under the Phytoseiulus persimilis application in greenhouses
Varieties of roses grown for cutting differ in the degree of costs for protection against pests, primarily from the two-spotted spider mite Tetranychus urticae Koch. To control this pest, from 6-8 to 25 or more treatments with acaricides are required. The predatory mite phytoseiulus Phytoseiulus persimilis A.-H. can be used as an alternative or addition to chemical treatments. Here, we report on a long-term monitoring of the spider mite abundance in commercial rose greenhouses. The observation allows us, for the first time, to assess a relationship between two Rosa hybrida variety-specific morphometric parameters, the area of a compound leaf segment and the total leaf area per bush, and an abundance of the spider mite in a triotrophic system, i.e., rose plant-spider mite-predatory mite. From this data, we obtained the equations to predict the development of the pest and determined the predatory mite number effective on a certain variety. This work aimed i) to assess the two-spotted spider mite infestation in a set of rose varieties, ii) to establish the relationship of the spider mite infestation level with the bush structure elements, and iii) to choose mathematical models for prediction of the pest infestation levels and the number of the predatory mite phytoseiulus necessary to use for the control of the pest. Observations on the two-spotted spider mite development were carried out in a block glass greenhouse of ZAO Agroleader (Vyborgsky District, Leningrad Province) on rose plants ( Rosa sp., hybrid tea group) of 18 varieties. The area of the greenhouse was 45,000 m2. A scoring system was used to assess the infestation levels of roses by spider mites. The greenhouse was divided into plots. Each plot was a 3.95 m long (8.02 m2 in area) segment of a double row of rose bushes. The survey consisted of a visual inspection of plants and assignment of the infestation level score from 1 to 5. Surveys were carried out twice a month, the total number of counts per year was at least 24. The dynamics of rose plant infestation by spider mites was assessed over 8 years (2011-2018). Since 2011, on particular varieties, and since 2012, on the entire area of the rose greenhouse, the predatory mite Ph. persimilis , introduced continuously or into the infestation foci, was used to control the two-spotted spider mite. Continuous application from 3 to 10 individuals/m2 over the entire area of the greenhouse was carried out 1-1.5 times a month; from 10 to 60 individuals per bush were introduced into foci weekly until new significant foci of the pest continued to appear. Acaricides were used only in cases where the T. urticae infestation level exceeded 2.5 points. Seven days after the first treatment the second treatment was carried out. We determined the number of stems in the upper part of the bush (crown) and on the whole bush, the productive stem length, the number of lobes of the complicated leaf, the number of leaves on the entire stem and on 10 centimeters of the stem, the number of leaves in the bush crown and on the entire bush, the surface areas of the lobule and the entire leaf, the area of the leaves surface in the crown and in the entire bush. Correlation analysis was used to assess the relationship between the occupancy of individual varieties of roses and the structural elements of their bushes, and regression analysis was used to describe it mathematically (rectilinear regression equations). To establish the relationship between the parameters of individual elements of the structure of rose bushes and the infestation level of spider mites, a two-factor ANOVA was used. When comparing the parameters of regression models built from sample data, the least squares method was used. In the most contrasting varieties Brazil and Aqua, the average long-term level of infection differed by 17.8 times. The remaining varieties could be divided into 6-8 groups, of which the most contrasting ones differed by 5.0
Текст научной статьи Связь заселяемости сортов роз паутинным клещом с элементами структуры куста в условиях применения хищного клеща фитосейулюса в теплицах
Культура розы, выращиваемая на срез, характеризуется значительным, постоянно увеличивающимся разнообразием сортов, которые различаются не только декоративными свойствами цветка, но и элементами структуры, архитектурой куста (1). Сорта различаются по степени затрат на защиту от вредителей, в первую очередь от обыкновенного паутинного клеща Tetranychus urticae Koch., для борьбы с которым на некоторых сортах приходится проводить по 25 и более обработок акарицидами, в то время как на других только 6-8 (2-4).
На заселение фитофагами могут влиять высота растения (5), площадь листовой поверхности (6, 7), структурная сложность (8) и их связь (соприкосновение листьев (9), число листьев на растении (1, 10), площадь и толщина листьев, их морфологическая структура — опушенность, плотность расположения трихом, их типы (11-14). Элементы структуры растения определяют наличие убежищ для фитофагов, распределение (16) и обилие фитофагов (17), а также косвенно влияют на естественных врагов за счет пространственного распределения добычи (18, 19). Кроме того, от архитектуры растений зависит выбор естественными врагами растения-хозяина (20), их движение и выживаемость (18, 21), иные особенности поведения хищников и паразитов (22), например хищническая активность акарифагов, их репродуктивное поведение, расселение и поисковая способность (23-25).
Сортовые особенности розы, влияющие на фитофага T. urticae , оценивали в основном в отношении биохимических признаков (содержание терпенов, дубильных веществ, эфирных масел) и морфологических признаков листа (трихомы, железы, толщина листа) (26, 27). Взаимодействие растения, T. urticae и его хищника фитосейулюса Phytoseiulus persimilis A.-H. на сортах роз с разной устойчивостью к фитофагу изучено недостаточно (28). Также мало известно о влиянии особенностей архитектуры растений на их заселяемость паутинным клещом и эффективность фитосейулюса (29), хотя это представляет научный интерес и необходимо для успешной, экономически выгодной защиты культуры от вредителя. Выявление элементов структуры куста розы, определяющих развитие фитофага и его хищника, позволит прогнозировать объемы защитных мероприятий на культивируемых и новых сортах.
Ранее мы установили значительные различия сортов роз, выращиваемых на срез, по степени их заселения паутинным клещом как в условиях применения акарицидов, так и при использовании фитосейулюса (2, 3).
В настоящей работе впервые выявлены элементы структуры куста розы, определяющие развитие фитофага T. urticae и его хищника Ph. persimilis , а также подобраны уравнения, позволяющие прогнозировать объемы защитных мероприятий на разных сортах Rosa hybrida , выращиваемых в теплицах на срез.
Целью работы была многолетняя оценка заселяемости различных сортов роз обыкновенным паутинным клещом, установление ее связи с особенностями отдельных элементов структуры кустов, а также подбор математических моделей, позволяющих прогнозировать заселяемость роз этим вредителем и необходимые объемы использования хищного клеща фито-сейулюса для борьбы с ним.
Методика. Наблюдения за развитием обыкновенного паутинного клеща проводили в блочной стеклянной теплице ООО «Агролидер» (Ленинградская обл., Выборгский р-н) на растениях розы ( Rosa hybrida ) сортов Aqua, Avalanche, Peach Avalanche, Wow, Dark Wow, Grand Prix, Miss Piggy, Penny Lane, Jumilia, Taleya, Myrna, Brazil, Heaven, Dolomiti, Hot Shot, Red Naomi, Deep Water, Fiesta.
Площадь теплицы составляла 45000 м2. Розы выращивали методом малообъемной гидропоники, используя в качестве субстрата минеральную вату Grodan («Grodan B.V.», Нидерланды). В теплицах регулировался микроклимат, применялось капельное орошение, искусственное освещение (4500 лк), зашторивание. В период освещения (с 400 до 000) поддерживалась температура не ниже +20 ° С, относительная влажность воздуха — 60-65 %, в период без освещения — соответственно не ниже +16 ° С и 70-75 % (в автоматическом режиме).
Использовали балльную систему оценки заселенности роз паутинным клещом. Для этого вся теплица была разделена на участки. Каждый участок представлял собой отрезок двойного ряда кустов роз. Длина отрезка составляла 3,95 м, его площадь — 8,02 м2. Число кустов на участке составляло в среднем 60 из расчета 7-8 кустов/м2. Число участков у разных сортов различалось, поскольку площадь под ними была неодинаковой. Минимальная площадь и число участков (всего 76) были у сорта Fiesta, максимальная (988 участков) — у сорта Grand Prix. Обследование заключалось в визуальном осмотре растений и присвоении балла заселенности для каждого участка по следующей шкале: 1 — паутинный клещ встречается в фабрике (части куста, сформированной посредством пригибания вниз побегов для повышения фотосинтеза); 2 — паутинный клещ встречается в короне (продуктивной части куста, состоящей из товарных побегов, цветоносов, и побегов для пригибания), перемещается в средний и верхний ярусы продуктивных стеблей, но еще не достигает бутона (десятки особей на заселенных листьях); 3 — появление на бутоне первых клещей и паутины (сотни особей на растениях участка); 4 — паутина появилась более чем на 50 % листьев, появляются «шапки» из паутины на бутонах (тысячи особей на растениях участка); 5 — все растение в паутине, скопления фитофага на бутонах и кончиках листьев, прекращение роста побегов и их деформация, усыхание и опадение листьев (в теплицах такая ситуация не допускается) (2-4). После каждого обследования определяли средний балл заселенности вредителем для каждого сорта на дату учета. Обследования проводили 2 раза в месяц, общее число учетов за год — не менее 24. Динамику заселенности роз паутинным клещом оценивали на протяжении 8 лет (2011-2018 годы). Для каждого сорта общее число учетов различалось, поскольку за период исследования некоторые сорта убирали из производства и вводили новые. Также для каждого сорта оценивали среднюю многолетнюю заселенность. По итогам минимальное число учетов (всего 48) было проведено на сорте Brazil, максимальное (195 учетов) — на сорте Deep Water. Среднее число учетов для всех сортов за 8 лет составляло 137,5±11,21. На основе результатов обследований принимали решение о проведении защитных мероприятий.
С 2011 года на части сортов, а с 2012 — на всей площади комбината для борьбы с обыкновенным паутинным клещом применяли хищного 460
клеща фитосейулюса Phytoseiulus persimilis , которого вносили, используя сплошной и локальный (в очаги вредителя) методы. Сплошное внесение от 3 до 10 особей/м2 по всей площади теплиц осуществляли 1-1,5 раза в месяц, в очаги — от 10 до 60 особей/куст (30) еженедельно до тех пор, пока продолжали появляться новые значительные очаги вредителя. При таком внесении, как правило, создается необходимое соотношение хищника и жертвы 1:10-1:20 (30, 31). Если в очагах фиксировали присутствие акарифага (1-5 особей/лист), его дополнительное внесение отменяли. Акарициды использовали только в тех случаях, когда заселенность роз T. urticae превышала 2,5 балла. Обработки были спаренными: через 7 сут после первой обработки проводили повторную.
Чтобы изучить влияние элементов структуры куста на развитие обыкновенного паутинного клеща и эффективность фитосейулюса, у всех 18 сортов роз оценили следующие морфометрические показатели: число продуктивных стеблей в верхней части куста (короне) и в пригибаемой части (фабрике) посредством их подсчета у 30 случайно выбранных кустов каждого сорта — сумма стеблей короны и фабрики давала показатель общего числа стеблей на кусте, затем рассчитывали среднее число стеблей на кусте для каждого сорта; длину продуктивных стеблей; число долек сложного листа и листьев на всем стебле посредством подсчета на 30 случайно выбранных стеблях каждого сорта (умноженное на 10 частное от деления числа листьев на всем стебле на его длину давало показатель числа листьев на 10 см стебля); число листьев в короне куста и на всем кусте, которое рассчитывали посредством умножения среднего числа листьев на всем стебле на среднее число стеблей в короне куста и на всем кусте; площадь дольки (простого листа), которую рассчитывали по формуле площади эллипса, для чего измеряли длину и ширину у 30 сложных листьев каждого сорта (количество измерений варьировало от 140 до 173, поскольку число долек листа, отклоняющееся от стандартных пяти, у сортов варьировало); площадь листовой поверхности куста или его короны, которую рассчитывали посредством умножения средней площади сложного листа на количество листьев на всем кусте или на его короне.
Статистическую обработку данных проводили с использованием программы SPSS и электронных таблиц Microsoft Excel. Рассчитывали средние баллы заселяемости сорта за все время наблюдений, средние значения измеренных параметров структуры кустов, а также стандартные ошибки средних (±SEM) для каждого сорта. Достоверность различий оценивали по t-критерию Стьюдента. Для оценки связи заселяемости отдельных сортов роз с особенностями элементов структуры их кустов использовали корреляционный анализ, а для ее математического описания — регрессионный (уравнения прямолинейной регрессии). Чтобы установить взаимосвязь параметров отдельных элементов структуры кустов роз со степенью их заселяемости паутинным клещом, применяли двухфакторный дисперсионный анализ (32). При сравнении параметров регрессионных моделей, построенных по выборочным данным, использовали метод наименьших квадратов (33). Ошибки уравнений регрессии рассчитывали посредством усреднения отклонений фактических значений среднних многолетних уровней заселяемости всех исследованных сортов роз паутинным клещом или потребовавшихся объемов выпусков фито-сейулюса с ожидаемыми по оцениваемой математической модели.
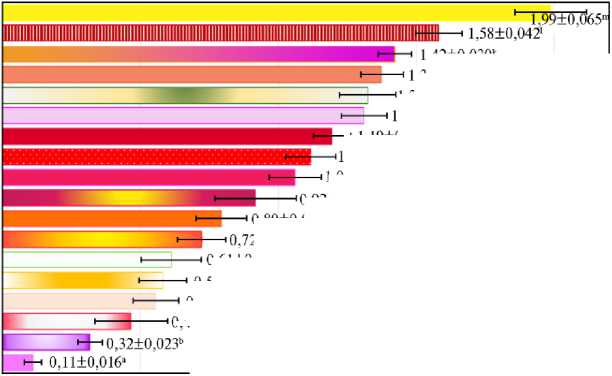
Результаты. Увеличение числа сортов и объединение результатов за несколько лет позволило выявить более широкий по сравнению предыдущими исследованиями (2, 3) состав групп, различающихся по особенностям, которые влияют на развитие обыкновенного паутинного клеща в условиях выпуска на растения фитосейулюса (рис. 1). Так, два сорта с наименьшей (Aqua, ¹ 1) и наибольшей (Brazil, ¹ 18) заселяемостью вредителем высокодостоверно (р < 0,001) отличались от остальных, различаясь между собой (в 17,8 раза), и должны считаться представителями наиболее контрастных по этому признаку групп.
Второй по заселяемости (Grand Prix, ¹ 17) и предпоследний (Deep Water, ¹ 2) сорта также статистически значимо (р < 0,001) отличались от всех остальных (между собой — только в 5 раз), поэтому должны считаться представителями еще двух групп. Пятую группу с относительно низкой за-селяемостью представляли сорта Jumilia, Peach Avalanch, Avalanche, Taleya (¹¹ 3-6) (см. рис. 1), достоверно не различающихся по этому признаку (при объеме выборок от 72 до 168). Сорта Taleya, Wow, Fiesta, Dark Wow, Hot Shot, Myrna, Red Naomi (¹¹ 6-12), показавшие среднюю заселяемость (0,61-1,19 балла), представляли несколько групп, поскольку каждый из них достоверно не отличался от предыдущего, но отличался (в основном при р < 0,05) от сорта, имеющего номер на две единицы меньше. Наконец, еще одну или две группы представляли сорта Heaven, Dolomiti, Miss Piggy, Penny Lane (¹¹ 13-16), среди которых по средней заселяемости паутинным клещом в присутствии фитосейулюса достоверно (р < 0,05) различались только сорта Heaven (¹ 13) и Pany Lane (¹ 16).
Brazil (№ 18)
1,4210,030*
1,3710,039» <1,3310,052» 1,31+0.042
—* 1,1910,035*
l,12±0,046hl
1,0610,049»
0,92+0,074*
0,S0±0,046=r
0 0,50 1,00 1.50 2,00
Заселенность, балл
Grand Prix (Ns 17)
Penny Lane (№ 16)
Miss Piggy (№ 15)
Dolomiti (Ne 14)
Heaven (Ns 13)
Red Naomi (Ns 12)
Myrna (Ns 11) a Hot Shot (Ne 10) 5 Dark Wow (№ 9)
Fiesta (Ne 8)
Wow (Ns 7)
0,6110,055е1 0,5810,043' 0,5610,042= 0,4710,066=
Taleya (Ns 6)
Avalanche (Ns 5)
Peach Avalanch (Ns 4)
Jumilia (Ns 3)
Deep Water (Ns 2)
Aqua (Ns 1)
Рис. 1. Средняя многолетняя заселяемость разных сортов розы ( Rosa sp.) чайно-гибридной группы обыкновенным паутинным клещом Tetranychus urticae Koch. при выпусках хищного клеща фито-сейулюса Phytoseiulus persimilis A.-H. ( M ±SEM, опыт в теплицах, ООО «Агролидер», Ленинградская обл., 2011-2018 годы). Планками погрешностей обозначены доверительные интервалы для вероятности 0,95; одинаковыми буквами отмечены значения, достоверно не различающиеся (р > 0,05) по t -критерию Стьюдента.
Анализ элементов структуры кустов розы показал значимую межсортовую изменчивость (29). Между самыми контрастными сортами различия были высокодостоверными (р < 0,001) по всем исследованным показателям. Однако по абсолютной величине они оказались не столь высокими, как по заселяемости T. urticae . Так, меньше всего стеблей в короне куста и на всем кусте (3,6±0,14 и 6,2±0,21) было у сорта Brazil, а больше всего — у Peach Avalanch (9,5±0,43 и 13,9±0,62). Самые длинные стебли оказались у сорта Grand Prix (74,1±1,49 см), самые короткие — у Heaven (60,4±0,98 см). Меньше всего долек в сложном листе было у Aqua (4,7±0,14), больше всего — у сорта
Heaven (5,7±0,17).
-
1. Показатели листовой поверхности у 18 сортов розы ( Rosa sp.) чайно-гибридной группы ( M ±SEM, опыт в теплицах, ООО «Агролидер», Ленинградская обл., 2011-2018 годы)
Сорт
Площадь листовой поверхности
листа, см2 ( n = 30)
дольки листа, см2 ( n от 140 до 173)
короны куста, м2 ( n = 30)
всего куста, м2 ( n = 30)
Aqua
115,7±3,81efg
24,8±1,00kl
0,79±0,050qr
1,16±0,077 α - δ
Deep Water
129,4±4,05bc
25,4±0,98kl
0,79±0,040qr
1,16±0,057 αβγ
Jumilia
172,5±7,60a
35,2±1,35i
0,61±0,091s-v
0,91±0,066 ζη
Peach Avalanch
110,9±4,13fgh
20,5±0,57no
0,94±0,040q
1,37±0,097 α
Avalanche
112,8±4,43fgh
20,9±0,68no
0,95±0,066q
1,40±0,125 αβ
Taleya
128,2±4,18bcd
24,3±0,70l
0,81±0,042qr
1,32±0,065 αβ
Wow
141,9±7,80bc
29,0±0,95j
0,56±0,040uv
0,98±0,067 δ - η
Fiesta
133,4±6,86bc
27,6±1,00jk
0,64±0,040su
1,14±0,072 β - ε
Dark Wow
140,7±7,60bc
28,7±0,85j
0,60±0,041tuv
0,99±0,069 γη
Hot Shot
127,9±4,57bcd
25,4±0,9kl
0,66±0,034stu
1,00±0,052 δεζ
Myrna
136,7±3,90b
25,2±0,80kl
0,62±0,034stu
1,00±0,055 δ - η
Red Naomi
125,7±3,78cde
23,2±0,76lm
0,59±0,039uv
0,96±0,062 εζη
Dolomiti
114,7±4,08efg
21,2±0,60no
0,66±0,032stu
1,07±0,050 γδε
Heaven
116,0±5,06d-g
20,2±0,76nop
0,71±0,049rst
1,11±0,076 γδε
Miss Piggy
104,8±5,81gh
18,2±0,60p
0,74±0,056rs
1,13±0,079 β - ε
Pany Lane
123,6±5,29b-f
22,2±0,68mn
0,53±0,031v
0,86±0,051 η
Grand Prix
112,2±4,30fgh
20,7±0,68no
0,51±0,035v
0,81±0,057 η
Brazil
101,1±4,89h
20,2±0,70o
0,37±0,024w
0,64±0,040 θ
Пр им еч ан и е. Одинаковыми буквами отмечены показатели,
не имеющие статистически значимых раз-
личий (р > 0,05) по
t -критерию Стьюдента.
-
2. Коэффициенты парной корреляции ( r n ±S r ) заселяемости обыкновенным паутинным клещом Tetranychus urticae Koch. 18 сортов розы ( Rosa sp.) чайногибридной группы с некоторыми показателями структуры кустов, уравнения прямолинейной регрессии и суммы квадратов отклонений фактической засе-ляемости от ожидаемой по уравнениям регрессии (опыт в теплицах, ООО «Агролидер», Ленинградская обл., 2011-2018 годы)
Показатель
r п ±S r
Вероятность отличия r п от нуля
Уравнение регрессии (y = а + bx или y = a + bxz)
Сумма квадратов отклонений
Стеблей в короне (x)
- 0,36±0,233
0,8 < Р < 0,9
y = 1,53 - 0,100x
3,60
Стеблей в кусте (x)
- 0,30±0,238
0,5 < Р < 0,8
y = 1,50 - 0,061x
3,74
Длинна стебля (x)
0,27±0,241
0,5 < Р < 0,8
y = - 0,97+0,031x
3,81
Число долек сложного листа (x)
0,49±0,218
0,95 < Р < 0,99
y = - 3,09+0,779x
3,07
Число листьев:
на всем стебле (x)
- 0,03±0,250
Р < 0,2
y = 1,25 - 0,024x
3,88
на 10 см стебля (x)
- 0,19±0,246
0,5 < Р < 0,8
y = 1,97 - 0,695x
4,05
в короне куста (x)
- 0,42±0,227
0,9 < Р < 0,95
y = 1,73 - 0,014x
3,14
на всем кусте (x)
- 0,35±0,234
0,8 < Р < 0,9
y = 1,70 - 0,008x
3,59
Площадь:
дольки листа (x)
- 0,52±0,214
0,95 < Р < 0,99
y = 2,44 - 0,060x
2,94
всего листа (х)
- 0,44±0,225
0,9 < Р < 0,95
y = 2,55 - 0,013x
3,23
короны куста (x)
- 0,70±0,179
P > 0,998
y = 2,54 - 2,326x
2,12
всего куста (x)
- 0,65±0,189
P > 0,995
y = 2,67 - 1,604x
2,31
короны куста (x) и дольки листа (z)
- 0,95±0,081
P > 0,999999
y = 2,92 - 0,120xz
0,406
всего куста (x) и дольки листа (z)
- 0,89±0,116
P > 0,99999
y = 2,92 - 0,077xz
0,790
-
3. Коэффициенты парной корреляции ( r п14 ±S r ) заселяемости обыкновенным паутинным клещом Tetranychus urticae Koch. 14 сортов розы ( Rosa sp.) чайногибридной группы с некоторыми показателями площади листовой поверхности и средние отклонения фактической заселенности от рассчитанной по уравнениям прямолинейной регрессии (опыт в теплицах, ООО «Агролидер», Ленинградская обл., 2011-2018 годы)
Площадь
r п14 ±S r
Вероятность отличия r п14 от нуля
Уравнение регрессии (y = а + bx или y = а + bxz)
Среднее отклонение
14 сортов
4 сорта
Дольки листа (x)
- 0,58±0,235
0,98 > Р > 0,95
y = 2,70 - 0,069x
0,28±0,074
0,43±0,131
Всего листа (x)
- 0,47±0,254
0,95 > Р > 0,90
y = 2,73 - 0,014x
0,31±0,076
0,43±0,132
Короны куста (x)
- 0,74±0,193
0,998 > Р > 0,995
y = 2,74 - 2,649x
0,14±0,040
0,23±0,069
Всего куста (x)
Короны куста (x) и дольки
- 0,68±0,211
0,995 > Р > 0,990
y = 2,77 - 1,705x
0,30±0,067
0,21±0,090
листа (z)
Всего куста (x) и дольки
- 0,94±0,096
> 0,999999
y = 2,88 - 0,118xz
0,12±0,029
0,12±0,037
листа (z)
- 0,89±0,134
> 0,99999
y = 2,85 - 0,071xz
0,14±0,043
0,21±0,059
Самое большое число листьев на всем стебле было у сорта Fiesta (12,5±0,32), на 10 см стебля — у сорта Deep Water (1,74±0,025), в короне куста — у Avalanche (84,8±7,55), на всем кусте — у Peach Avalanch (125±7,4). В то же время меньше всего листьев по всем четырем показателям было у одного сорта — Jumilia (соответственно 7,8±0,27; 1,22±0,036; 35,5±2,16 и 53±3,5). Наоборот, у сорта Jumilia отмечена самая большая площадь дольки и всего листа (табл. 1). Наименьшей площадь дольки листа была у сорта Miss Piggy, всего листа — у Brazil. При этом самая большая площадь листовой поверхности короны куста и всего куста выявлена у сорта Avalanche, а самая маленькая — у Brazil. Остальные сорта по ряду показателям занимали промежуточное положение. Они статистически значимо различались или не различались между собой.
Более определенную картину связи элементов структуры кустов розы с их заселяемостью паутинным клещом дали результаты корреляционного и регрессионного анализов (табл. 2). Достоверной связи 8 из 12 исследованных морфометрических признаков структуры кустов роз с их засе-ляемостью T. urticae выявить не удалось, поскольку вероятность отличия коэффициента корреляции от нуля была меньше 0,95, а для числа листьев на всем стебле она приближалась к нулю. При этом сумма квадратов отклонений ожидаемой заселенности по рассчитанным уравнениям регрессии для 18 сортов была существенно больше 3.
Отмечена связь (с вероятностью отличия коэффициента корреляции от нуля > 0,95, но < 0,99) заселенности T. urticae сортов роз с площадью листовой поверхности дольки листа (средняя отрицательная) и числом долек сложного листа (средняя положительная). Отрицательная связь заселенности роз паутинным клещом с площадью листовой поверхности короны и всего куста оказалась более высокой, с вероятностью > 0,99, но < 0,999, а сумма квадратов отклонений — более 2.
Однако прогностическая точность ожидаемой заселяемости сорта паутинным клещом при использовании таких простых моделей невысока. Средняя погрешность степени заселяемости будет составлять от 0,27 балла при использовании в качестве предиктора общей площади листовой поверхности куста, до 0,33 балла — при использовании площади дольки листа. В связи с этим мы попытались найти модель, которая учитывает как факторы, определяющие климат в зоне куста (площадь листовой поверхности короны и всего куста), так и фактор, влияющий на микроклимат в ламинарном слое листа (площадь дольки листа), посредством их перемножения. Коэффициенты корреляции таких показателей с заселяемостью исследованных сортов роз T. urticae при выпуске фитосейулюса резко увеличились, а сумма квадратов отклонений снизилась (см. табл. 2). Двухфакторный дисперсионный анализ не показал взаимодействие площади дольки листа и площадей листовой поверхности короны куста или всего куста в их влиянии на заселяемость сортов роз паутинным клещом в теплицах, что свидетельствует о возможности их использования в модели, как независимых предикторов.
Использование в качестве второго аргумента среднего числа долек у сложного листа розы оказалось малоэффективным. Коэффициенты корреляции произведений среднего числа долек листа на площади листовой поверхности короны или всего куста с заселяемостью паутинным клещом составили только 0,52±0,214 (0,99 > Р > 0,95) и 0,46±0,223 (0,95 > Р > 0,90), а их прогностическая погрешность — 0,34±0,055 и 0,33±0,063 балла. К тому же выявлено достоверное взаимодействие среднего числа долек с площадью листовой поверхности короны и всего куста в их влиянии на заселяемость сортов роз паутинным клещом, что затрудняет использование этих показателей в модели прямолинейной регрессии.
Для верификации предлагаемой модели прогнозирования заселяе-мости сортов роз паутинным клещом в условиях применения фитосейулюса по таблице случайных чисел было выбрано четыре сорта: Deep Water, Avalanche, Dark Wow и Miss Piggy. Остальные 14 сортов использовали для расчетов коэффициентов корреляции и уравнений регрессии, на основании которых были спрогнозирована ожидаемая заселяемость четырех исключенных сортов (табл. 3). Средние отклонения их ожидаемой заселяемости практически не отличались от таковых для 14 сортов, включенных в расчеты.
Примечание. 14 сортов служили основой для расчетов коэффициентов корреляции и уравнений ре- грессии, 4 сорта выбраны для верификации по таблице случайных чисел.
Уравнение, основанное на произведении площадей листовой поверхности короны куста и дольки листа, показало лучшие прогностические свойства. При этом средняя погрешность составляла всего 0,12 балла. Следует отметить, что выявление этих элементов структуры кустов в качестве лучших предикторов заселяемости сортов роз T. urticae в условиях использования хищного клеща фитосейулюса не случайно. Оба фактора определяют климат, влияющий на развитие как фитофага, так и акарифага: от общей площади листовой поверхности зависит влажность и температура в зоне куста, от площади дольки листа — влажности в ламинарном слое листа толщиной до 3-5 мм (34, 35). Особое значение влажность имеет для эффективного использования фитосейулюса. Она влияет на его выживаемость, особенно на эмбриональной и личиночной стадиях развития акарифага, и на репродуктивное поведение самок (36-38).
Дальнейшее совершенствование модели осуществляли посредством уточнения коэффициентов в уравнениях, предназначенных для прогнозирования заселяемости сортов роз в первый год применения фитосейулюса и при его стабильном использовании, поскольку они существенно различались (4), а также за счет снижения погрешности прогноза, исключив сорт Aqua, фактическая заселяемость которого выходила за пределы 95 % доверительной зоны регрессии. Для первого года применения фитосейулюса рекомендуется уравнение у п = 2,57 - 0,073xz, погрешность которого составляла 0,102±0,0154 балла, а при стабильном использовании у п = 2,89 - 0,127xz, погрешность которого составляла 0,081±0,0156 балла (рис. 2, А), где у п — средняя заселяе-мость сорта роз обыкновенным паутинным клещом, баллы, х — средняя площадь дольки листа, см2; z — средняя площадь листовой поверхности короны куста, м2. Эти уравнения можно использовать при подборе новых сортов для промышленного выращивания в теплицах, учитывая фактор защиты от обыкновенного паутинного клеща. Полученные данные позволили составить прогностические уравнения и для расчетов объемов фитосейулюса, требующихся при его использовании и в первые годы, и в течение достаточно длительного периода времени. Это важно для планирования объемов массового разведения или закупки хищного клеща.
После сравнения нескольких вариантов уравнений для повышения точности прогнозирования исключили самые контрастные по требовавшемуся объему акарифага сорта Brazil и Aqua, которые выпадали за пределы 95 % доверительной зоны регрессии (39). Для прогнозирования необходимых объемов выпусков фитосейулюса на большинстве сортов роз в первый год рекомендуется уравнение уф = 345 - 11,3xz, погрешность которого составляет 22,0±5,52 особи на м2 в год, а при стабильном использовании — уф = 278 - 11,1xz, погрешность которого составляет 9,8±1,36 особи на м2 в год (см. рис. 2, Б). В этих уравнениях уф — объем фитосейулюса, необходимый для выпусков с целью защиты конкретного сорта роз от паутинного клеща в течение года, особей/м2; х — средняя площадь дольки листа у сорта роз, см2; z — средняя площадь листовой поверхности короны куста у сорта роз, м2.
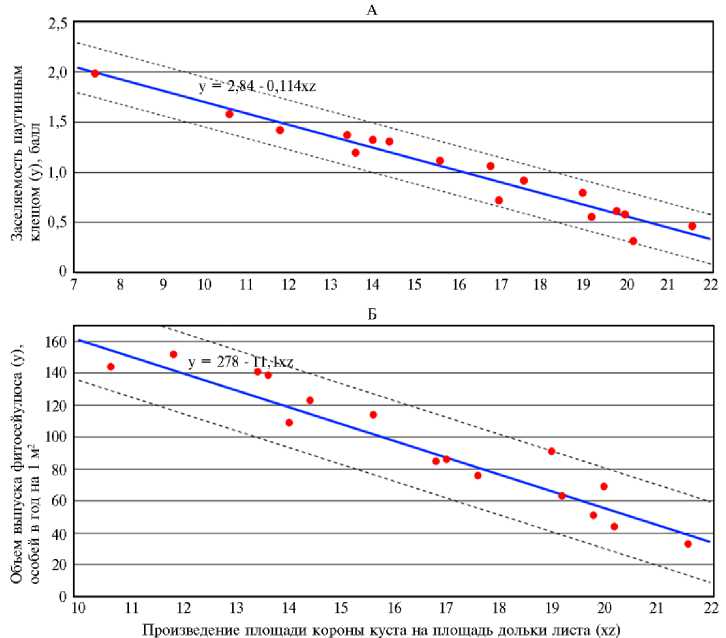
Рис. 2. Зависимость заселяемости розы ( Rosa sp.) чайно-гибридной группы обыкновенным паутинным клещом Tetranychus urticae Koch. (А) и необходимых объемов выпусков хищного клеща фитосейулюса Phytoseiulus persimilis A.-H. (Б) от произведения площадей листовой поверхности короны куста и дольки листа определенного сорта розы: красные точки — фактическая заселенность, синяя линия — расчетная заселяемость, пунктирные линии — граница зоны регрессии (опыт в теплицах, ООО «Агролидер», Ленинградская обл., 2011-2018 годы)
Из 18 исследованных сортов роз защита от паутинного клеща с помощью фитосейулюса оказалась наиболее результативной у сорта Aqua. В первый год потребовалось только 9 выпусков хищника общей численностью 48 особей/м2. В последующие годы число потребовавшихся выпусков снизилось до 4-5, а их объем сначала снизился до 11-19 (2-4-й годы), а затем до 68 особей/м2 в год (4). За 7 лет на сорте Aqua ни разу не потребовалось использовать акарициды. На cорте Brazil, напротив, защита с помощью фито-сейулюса оказалась крайне неэффективной. Выпуски акарифага пришлось проводить по 2 раза в месяц. Общая численность выпущенных хищных клещей на 1 м2 в первый год составила 418, во второй — 390 особей. При этом дополнительно осуществляли соответственно 7 и 6 двойных обработок акарицидами.
В дальнейшем от выращивания сорта Brazil хозяйству пришлось отказаться. Переход на использование только акарицидов для борьбы с T. urticae на этом сорте роз требовал 13-15 двойных обработок в год, что при наличии всего двух препаратов, разрешенных в Российской Федерации к применению на розах в защищенном грунте (39), крайне ограничивало возможность их чередования и привело к быстрому возникновению устойчивой популяции фитофага. В настоящее время этот список расширен до пяти препаратов (40), но в основном за счет химических акарицидов, постоянное применение которых в теплицах нежелательно. Совершенствование биологической борьбы с паутинным клещом с помощью хищного клеща Ph. persimilis остается крайне актуальным. Наше исследование позволяет подбирать более пригодные для биологической защиты сорта роз, а также планировать объемы применения хищного клеща. Подобранные модели, основанные на использовании в качестве предикторов показателей площади дольки листа и площади листовой поверхности короны куста, просты в использовании и представляются удовлетворительными по точности. Так, при прогнозировании заселяемости сорта паутинным клещом и необходимых объемов выпусков фитосейулюса при его постоянном использовании коэффициенты детерминации r 2 составляют соответственно 95,6 и 90,9 %.
Выявление морфологических, биохимических и других факторов (26-28), влияющих на размножение T. urticae и Ph. persimilis в системе трио-трофа (сорт растения, фитофаг, энтомофаг), представляет отдельный научный интерес. Значительных работ на розе в этом направлении исследований не было. Имеются наблюдения австралийских ученых, выполненные на розах, которые выращивались в открытом грунте, где отмечено, что у густо растущих сортов обилие соприкасающихся листьев способствовало быстрому перемещению Ph. persimilis по растению. Долгосрочные взаимодействия между обыкновенным паутинным клещом и хищником на этих розах привели к удовлетворительному результату борьбы с вредителем в сравнении с сортами, где крона не соприкасалась (41, 42).
В Иране на 10 сортах роз были проведены лабораторные исследования по оценке основных жизненных параметров развития обыкновенного паутинного клеща. Некоторые сорта достоверно различались по влиянию на выживаемость, продолжительность развития неполовозрелых и плодовитость взрослых клещей, а также на скорость роста численности популяций, репродуктивную способность и среднее время продолжительности генерации (27). Связь с морфологическими или биохимическими особенностями сортов не оценивали.
В Мексике были показаны существенные различия некоторых из 13 сортов по степени благоприятствования развитию T. urticae , а также отрицательная корреляция этого показателя с содержанием эфирных масел и положительная корреляцию с содержанием терпенов, азота и толщиной листа (26). Там же, в Мексике, через 1 нед после равномерного заселения вредителем двух сортов роз, контрастных по условиям для развития T. urticae , отмечали, что плотность наносимых T. urticae повреждений и содержание хлорофилла в листьях роз не различались. После выпуска Ph. persimilis плотность T. urticae на обоих сортах изменилась. Различий в проценте повреждений и плотности Ph. persimilis между сортами не зафиксировали. Один из сортов имел самое низкое содержание хлорофилла, но на нем Ph. persimilis был более эффективен (28).
Проведенные нами исследования показали значимую отрицательную связь развития обыкновенного паутинного клеща в присутствии фито-сейулюса, то есть в системе триотрофа, с изменениями показателей площади дольки сложного листа и площади листовой поверхности короны куста и всего куста у разных сортов роз. Эта связь отражается в необходимых объемах внесения хищника на сортах с разными показателями этих элементов структуры кустов роз.
Таким образом, обобщение результатов учетов заселенности 18 сортов роз, выращиваемых в теплицах на срез в условиях применения хищного клеща Phytoseiulus persimilis A.-H. для биологической борьбы с обыкновенным паутинным клещом Tetranychus urticae Koch. на протяжении 8 лет, показало значительное разнообразие по заселяемости сортов этим фитофагом. Два наиболее контрастных по средней многолетней заселяемости сорта Brazil и Aqua различались в среднем в 17,8 раза. Остальные сорта можно было разделить на несколько (шесть-восемь) групп, из которых наиболее контрастные различались в 5 раз. Сорта роз достоверно различались по средним показателям следующих элементов структуры куста: числу стеблей в короне и на всем кусте, долек у сложного листа, листьев на всем стебле и на 10 см стебля, листьев в короне и на всем кусте, длине продуктивного стебля, площади дольки и всего листа, листовой поверхности куста и его короны. Из 12 оцененных показателей структуры куста достоверная связь с засе-ляемостью сортов розы паутинным клещом в присутствии фитосейулюса была отмечена только для четырех — для числа долек у сложного листа ( r = 0,49±0,218; 0,95 < Р < 0,99), площади дольки листа ( r = - 0,52±0,214; 0,95 < Р < 0,99), площади листьев короны куста ( r = - 0,70±0,179; P > 0,998) и площади листьев всего куста ( r = - 0,65±0,189; P > 0,995). Очень тесная связь обнаружена между заселяемостью сортов роз вредителем и произведением площади дольки листа на площадь листьев всего куста ( r = - 0,89±0,134; Р > 0,99999) или на площадь листьев короны куста ( r = - 0,94±0,096; Р > 0,999999). Подобраны уравнения прямолинейной регрессии для прогнозирования средней заселяемости сорта паутинным клещом T. urticae в первый год применения фитосейулюса — у п = 2,57 - 0,073xz (погрешность 0,102±0,0154 балла), при стабильном использовании фитосейулюса — у п = 2,89 - 0,127xz (погрешность 0,081±0,0156 балла), а также для прогнозирования необходимых годовых объемов выпусков хищного клеща в первый год — у ф = 345 - 11,3xz (погрешность 22,0±5,52 особи на 1 м2 в год) и при стабильном использовании — у ф = 278 - 11,1xz (погрешность которого составляет 9,8±1,36 особи на 1 м2 в год), где у п — среднеяя заселяемость сорта паутинным клещом, особей/м2; у ф — объем фитосейулюса, необходимый для выпусков, с целью защиты сорта от паутинного клеща в течение года, осо-бей/м2; х — средняя площадь дольки листа, см2; z — средняя площадь листовой поверхности короны куста, м2. Эти уравнения рекомендуется использовать при биологической защите роз от обыкновенного паутинного клеща с помощью Ph. persimilis .
Список литературы Связь заселяемости сортов роз паутинным клещом с элементами структуры куста в условиях применения хищного клеща фитосейулюса в теплицах
- Cloyd R.A., Sadof C.S. Effects of plant architecture on the attack rate of Leptomastix dactylopii (Hymenoptera: Encyrtidae), a parasitoid of the citrus mealybug (Homoptera: Pseudococcidae). Environmental Entomology, 2000, 29(3): 535-541 (doi: 10.1603/0046-225X-29.3.535).
- Козлова Е.Г., Моор В.В. Применение Phytoseiulus persimilis против паутинного клеща на разных сортах роз. Защита и карантин растений, 2012, 12: 16-20.
- Моор В.В., Анисимов А.И., Козлова Е.Г. Вариабельность заселяемости сортов розы паутинным клещом Tetranychus urticae на фоне биологической и химической защиты. Вестник защиты растений, 2021, 104(4): 218-222 (doi: 10.31993/2308-6459-2021-104-4-15129).
- Моор В.В., Козлова Е.Г. Многолетнее применение фитосейулюса против паутинного клеща на розе. Защита и карантин растений, 2021, 11: 15-19 (doi: 10.47528/1026-8634-2021-11-15).
- Thorpe K.W. Effects of height and habitat type on egg parasitism by Trichogramma minutum and T. pretiosum (Hymenoptera: Trichogrammatidae). Agriculture, Ecosystems & Environment, 1985, 12: 117-126 (doi: 10.1016/0167-8809(85)90072-6).
- Kanour W.W., Burbutis P.P. Trichogramma nubilale (Hymenoptera: Trichogrammatidae) field releases in corn and a hypothetical model for control of European corn borer (Lepidoptera: Pyr-alidae). Journal of Economic Entomology, 1984, 77(1): 103-107 (doi: 10.1093/jee/77.1.103).
- Попов С.Я. Пономаренко Е.К. Прогностическая оценка сортов земляники на устойчивость (восприимчивость) к паутинному клещу. Известия Тимирязевской сельскохозяйственной академии 2016, 5: 55-67.
- Andow D.A., Prokrym D.R. Plant structural complexity and host-finding by a parasitoid. Oeco-logia, 1990, 82(2): 162-165 (doi: 10.1007/BF00323530).
- Skirvin D., Fenlon J.S. Of mites and movement: the effects of plant connectedness and temper-ature on movement of Phytoseiulus persimilis. Biological Control, 2003, 27(3): 242-250 (doi: 10.1016/S1049-9644(03)00022-7).
- Stamp N.E., Browers M.D. Presence of predatory wasps and stinkbugs alters foraging behavior of cryptic and non-cryptic on plantain (Plantago lanceolata). Oecologia, 1993, 95(3): 376-384 (doi: 10.1007/BF00320992).
- Krips O.E. Plant effects on biological control of spider mites in the ornamental crop Gerbera. PhD dissertation. Landbouw universiteit Wageningen, Netherlands, 2000.
- Raghu S., Drew R.A.I., Clarke A.R. Influence of host plant structure and microclimate on the abundance and behavior of a tephritid fly. Journal of Insect Behavior, 2004, 17(2): 179-190 (doi: 10.1023/B:JOIR.0000028568.90719.2a).
- Sarwar M. Influence of host plant species on the development, fecundity and population density of pest Tetranychus urticae Koch (Acari: Tetranychidae) and predator Neoseiulus pseudolongispi-nosus Xin, Liang and Ke (Acari: Phytoseiidae). New Zealand Journal of Crop and Horticultural Science, 2014, 42(1): 10-20 (doi: 10.1080/01140671.2013.817444).
- Amoah B., Anderson J., Erram D., Gomez J., Harris A., Kivett J., Ruang-Rit K., Wang Y., Murray L., Nechols J. Plant spatial distribution and predator–prey ratio affect biological control of the twospotted spider mite Tetranychus urticae (Acari: Tetranychidae) by the predatory mite Phytoseiulus persimilis (Acari: Phytoseiidae). Biocontrol Science and Technology, 2016, 26(4): 548-561 (doi: 10.1080/09583157.2015.1133807).
- Freese G. Structural refuges in two stem boring weevils on Rumex crispus. Ecological Entomology, 1995, 20(4): 351-358 (doi: 10.1111/j.1365-2311.1995.tb00467.x).
- Clark T.L., Messina F.J. Foraging behavior of lacewing larvae (Neuroptera: Chrysopidae) on plants with divergent architectures. Journal of Insect Behavior, 1998, 11: 303-317 (doi: 10.1023/A:1020979112407).
- Lawton J.H. Plant architecture and the diversity of phytophagous insects. Annual Review of Ento-mology, 1983, 28: 23-39 (doi: 10.1146/annurev.en.28.010183.000323).
- Stavrinides M.C., Skirvin D.J. The effect of chrysanthemum leaf trichome density and prey spatial distribution on predation of Tetranychus urticae (Acari: Tetranychidae) by Phytoseiulus persimilis (Acari: Phytoseiidae). Bulletin of Entomological Research, 2003, 93(4): 343-350 (doi: 10.1079/BER2003243).
- Gontijo L.M. Effects of plant architecture and prey distribution on the foraging efficiency and behavior of the predatory mite Phytoseiulus persimilis (Acari: Phytoseiidae). M.S. Thesis. Kansas State Uni-versity, Manhattan, KS, 2008.
- Romero G.Q., Vasconcellos-Neto J. The effects of plant structure on the spatial and microspatial distribution of a bromeliad-living jumping spider (Salticidae). Journal of Animal Ecology, 2005, 74(1): 12-21 (doi: 10.1111/j.1365-2656.2004.00893.x).
- Grevstad F., Klepetka B.W. The influence of plant architecture on the foraging efficiencies of a suite of ladybird beetles feeding on aphids. Oecologia, 1992, 92(3): 399-404 (doi: 10.1007/BF00317466).
- Legrand A., Barbosa P. Plant morphological complexity impacts foraging efficiency of adult Coc-cinella septempunctata L. (Coleoptera: Coccinellidae). Environmental Entomology, 2003, 32(5): 1219-1226 (doi: 10.1603/0046-225X-32.5.1219).
- Gontijo L.M., Margolies D.C., Nechols J.R., Cloyd R.A. Plant architecture, prey distribution and predator release strategy interact to affect foraging efficiency of the predatory mite Phytosei-ulus persimilis (Acari: Phytoseiidae) on cucumber. Biological Control, 2010, 53(1): 136-141 (doi: 10.1016/j.biocontrol.2009.11.007).
- Gontijo L.M., Nechols J.R., Margolies D.C., Cloyd R.A. Plant architecture and prey distribution influence foraging behavior of the predatory mite Phytoseiulus persimilis (Acari: Phytoseiidae). Experimental and Applied Acarology, 2012, 56(1): 23-32 (doi: 10.1007/s10493-011-9496-7).
- Skirvin D.J., De Courcy Williams M. Differential effects of plant species on a mite pest (Tetranychus urticae) and its predator (Phytoseiulus persimilis): implications for biological control. Experimental and Applied Acarology, 1999, 23(6): 497-512 (doi: 10.1023/a:1006150521031).
- Flores-Canales R.J., Mendoza-Villareal R., Landeros-Flores J., Cerna-Chávez E., Robles- Bermúdez A., Isiordia-Aquino N. Morphological and biochemical characters of Rosa ½ hybrida against Tetranychus urticae Koch in greenhouse. Revista Mexicana de Ciencias Agrícolas, 2011, 3: 473-482.
- Golizadeh A., Ghavidel S., Razmjou J., Fathi S.A., Hassanpour M. Comparative life table analysis of Tetranychus urticae Koch (Acari: Tetranychidae) on ten rose cultivars. Acarologia, 2017, 57(3): 607-616 (doi: 10.24349/acarologia/20174176).
- Chacon-Hernandez J.C., Camacho-Aguilar I., Cerna-Chavez E., Ordaz-Silva S., Camacho-Agui-lar I., Ochoa-Fuentes Y.M., Landeros-Flores J. Effects of Tetranychus urticae and Phytoseiulus persimilis (Acari: Tetranychidae: Phytoseiidae) on the chlorophyll of rosal plants (Rosa sp.). Ag-rociencia, 2018, 52(6): 895-909.
- Козлова Е.Г., Анисимов А.И., Моор В.В. Изменчивость сортов роз, выращиваемых на срез, по показателям архитектоники их куста. Мат. Межд. науч.-практ. конф. «Агробио-технология-2021». М., 2021: 864-871 (doi: 10.26897/978-5-9675-1855-3-2021-181).
- Чалков А.А. Биологическая борьба с вредителями овощных культур защищенного грунта. М., 1986.
- Gacheri C., Kigen Th., Sigsgaard L. Hot-spot application of biocontrol agents to replace pesti-cides in large scale commercial rose farms in Kenya. BioControl, 2015, 60(6): 795-803 (doi: 10.1007/s10526-015-9685-0).
- Урбах В.Ю. Биометрические методы. М., 1964.
- Магнус Я.Р., Катышев П.К., Пересецкий А.А. Эконометрика. Начальный курс. М., 2007.
- Gaede K. On the water balance of Phytoseiulus persimilis A.-H. and its ecological significance. Experimental and Applied Acarology, 1992, 15: 181-198 (doi: 10.1007/BF01195790).
- Boulard T., Mermier M., Fargues J., Smits N., Rougier M., Roy J.C. Tomato leaf boundary layer climate: implications for microbiological whitefly control in greenhouses. Agricultural and Forest Meteorology, 2002, 110(3): 159-176 (doi: 10.1016/S0168-1923(01)00292-1).
- Bernstein C. Some aspects of Phytoseiulus persimilis (Acarina: Phytoseiidae) dispersal behaviour. Entomophaga, 1983, 28(2): 185-198 (doi: 10.1007/BF02372143).
- Ferro D.N., Southwick E.E. Microclimates of small arthropods: estimating humidity within the leaf boundary layer. Environmental Entomology, 1984, 13(4): 926-929 (doi: 10.1093/ee/13.4.926).
- Le Hesran S., Groot T., Knapp M., Bukovinszky T., Forestier T., Dickeb M. Phenotypic variation in egg survival in the predatory mite Phytoseiulus persimilis under dry conditions. Biological Con-trol, 2019, 130: 88-94 (doi: 10.1016/j.biocontrol.2018.10.007).
- Государственном каталоге пестицидов и агрохимикатов, разрешенных к применению на территории Российской Федерации. Часть 1. Пестициды. М., 2013.
- Государственном каталоге пестицидов и агрохимикатов, разрешенных к применению на территории Российской Федерации. Часть 1. Пестициды. М., 2023.
- Gough N. Long term stability in the interaction between Tetranychus urticae and Phytoseiulus persimilis producing successful integrated control on roses in southeast Queensland. Experimental and Applied Acarology, 1991, 12(1-2): 83-101 (doi: 10.1007/BF01204402).
- Mites (Acari) for pest control /U. Gerson, R. Smiley, R. Ochoa (eds.). Blackwell Science, Oxford, 2003 (doi: 10.1002/9780470750995).
