Таксономическая структура эпифитона малых рек северо-западного побережья Ладожского озера

Автор: Станиславская Е.В.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Гидробиология - биологические науки
Статья в выпуске: 5 т.25, 2023 года.
Бесплатный доступ
Приводятся результаты исследования таксономической структуры и разнообразия прикрепленных водорослей малых рек северо-западного побережья Ладожского озера. В составе эпифитона рек было выявлено 323 вида, разновидности и формы водорослей, принадлежащих 7 отделам: Bacillariophyta - 122 (38%), Charophyta - 69 (22%), Chlorophyta - 54 (17%), Cyanoprokaryota - 45 (12%), Euglenophyta - 25 (8%), Xanthophyta - 5 (2%), Rhodophyta 3 - (1%). Основу альгофлоры составляли диатомовые, харовые, зеленые водоросли и цианопрокариоты, среди них наиболее представительны семейства Desmidiaceae, Closteriaceae, Fragilariaceae и роды - Closterium, Cosmarium, Nitzschia. Структуру и разнообразие альгофлоры определяют географическое положение и особенности водосборного бассейна.
Альгофлора, таксономическая структура, пропорции флоры, эпифитон, малые реки, ладожское озеро
Короткий адрес: https://sciup.org/148327977
IDR: 148327977 | УДК: 574.586.583 | DOI: 10.37313/1990-5378-2023-25-5-175-181
Taxonomic structure of the epiphyton of small rivers of the northwestern coast of lake Ladoga
The taxonomic structure and diversity of attached algae in small rivers of the northwestern coast of Lake Ladoga are presented. A total 323 algal species from 7 divisions were found: Bacillariophyta - 122 (38%), Charophyta - 69 (22%), Chlorophyta - 54 (17%), Cyanoprokaryota - 45 (12%), Euglenophyta - 25 (8 %), Xanthophyta - 5 (2%), Rhodophyta 3 - (1%). Divisions Bacillariophyta, Charophyta,Chlorophyta, Cyanoprokaryota are most various and numerous. The most representative families are Desmidiaceae, Closteriaceae, Fragilariaceae and the genera Closterium, Cosmarium, Nitzschia. The structure and diversity of the algoflora are determined by the geographic location and characteristics of the catchment area.
Текст научной статьи Таксономическая структура эпифитона малых рек северо-западного побережья Ладожского озера
Одной из важнейших задач сохранения природного биоразнообразия является изучение структурных характеристик и распространения автотрофных сообществ различных водоемов и водотоков. Среди этих сообществ значительное место занимают водоросли, обрастающие высшие водные растения (эпифитон). Наименее изученным, в настоящее время, остается эпи-фитон малых рек и ручьев. Малые реки играют важную роль в формировании крупных гидрографических сетей, являясь ее основой. Состояние экосистем малых рек во многом определяют экологическое состояние водотоков и водоемов, в которые они впадают. Развитие эпифитона в реках определяется большим количеством факторов, среди которых важными можно считать наличие подходящего субстрата, состояние берегов, скорость течения, химизм воды. Также большое влияние на прикрепленные сообщества оказывает затенение берегов деревьями и кустарниками, что особенно актуально для небольших рек, протекающих среди полей и лесных массивов [1]. Сочетание этих факторов может приводить как к развитию богатой разнообразной альгофлоры, а также к значительному угнетению обрастаний [2].
В летний период 2019-2022 гг. комплексной экспедицией Института озероведения СПб-ФИЦ РАН проводились исследования малых притоков западного побережья Ладожского озера, что позволило оценить их влияние на прибрежные акватории. Ладожское озеро - крупнейший, пресный водоем Европы, на формирование гидрохимического режима которого, а также на состав и разнообразие его альгоценозов существенное влияние оказывают впадающие в него реки. Альгофлора озера и его крупных притоков достаточно хорошо изучена, первые наблюдения были начаты еще в начале прошлого века [3]. В 60-х гг. прошлого века начались регулярные работы по изучению фитопланктона и перифитона озера, которые продолжаются до настоящего времени [4, 5, 6, 7]. Немного позже началось изучение планктонных сообществ наиболее крупных притоков Ладожского озера [8, 9, 10]. С конца 90-х гг. прошлого века начинаются более детальные исследования прикрепленных сообществ притоков озера. Перифитон ряда рек, протекающих по Ленинградской области, описан в публикациях С. Ф. Комулайнена [11, 1]. Нашими работами был изучен перифитон и эпилитон 20 наиболее крупных притоков, исследования которых проводились на протяжении нескольких лет [12, 13, 14]. Альгофлора малых рек Ленинградской области, имеющих сток в Ладожское озеро практически не изучена.
Целью данной работы было выявление таксономического состава и видового разнообразия водорослей эпифитона малых рек северозападного побережья Ладожского озера.
МАТЕРИАЛ И МЕТОДЫ
Исследования водорослей эпифитона 5-ти малых рек северо-западного побережья Ладожского озера проводились в вегетационный сезон 2019-2022 гг. Были обследованы реки Авлога, Морье, Сосновка, Вьюн, Семужья. Реки Авлога, Морье, Семужья непосредственно впадают в Ладожское озеро. Реки Сосновка и Вьюн имеют сток в Ладожское озеро через озерно-речную систему Вуоксы (рис.1). Водосборные бассейны исследованных рек относятся к Балтийско-Ладожскому бассейновому округу и расположены в юго-восточной части Карельского перешейка.
Морфологические и гидрологические характеристики рек различны, а также различались площади их водосборов (табл. 2). В зависимости от длины рек были выбраны 2-4 станции, которые располагались в истоке, среднем течении и устье (рис.1, табл. 1).
Среди растительности рек преобладали осоки, хвощи, ежголовники, стрелолисты, кубышки, с которых и были собраны пробы эпифитона. Пробы эпифитона отбирались в мае, июле и сен- тябре. Всего было собрано и обработано 70 качественных и количественных проб эпифитона.
Для водосборов, которые дренируют исследованные водотоки, характерны в целом, значительная освоенность, а также высокая заболоченность, наличие хвойных и смешанных лесов. Кроме того, на водосборах исследованных рек встречались торфоразработки, военные полигоны и свалки твердых бытовых отходов, сельскохозяйственные фермы, посёлки и большое количество садоводств. Распределение антропогенных источников по водосборным территориям неравномерно. Так, для рек Морье, Авлога и Сосновка максимум антропогенной нагрузки приходился на верхнее и среднее течение, а для рек Вьюн и Семужья – на нижнее.
Реки имели разные гидрохимические характеристики. Минерализация ( Σ ион) в них была относительно низкой и изменялась от 34 до 130 мг/л. Наименее минерализованы воды рек Вьюн и Морье, максимальная минерализация была отмечена в р. Авлоге. Цветность изменялась от 20о до 220о и была максимальной в р. Морье, которая протекает по торфоразработкам и заболоченным территориям. Величины рН в исследованных реках целом определяли нейтральную реакцию среды и изменялись в незначительных пределах – от 6,5 до 7,7. Самые низкие величины кислотности среды были отмечены в реках Морье и Семужья,
Таблица 1. Координаты станций отбора проб эпифитона на исследованных реках
Таблица 2. Морфологические и гидрологические характеристики исследованных рек
|
Река |
Sводосбора (км2) |
Длина (км) |
Ширина (м) |
Глубина(м) |
Скорость течения (м/сек) |
|
Авлога |
375 |
54 |
4-10 |
0,4-0,6 |
0,01-0,07 |
|
Морье |
478 |
43 |
10-16 |
1,2-2,3 |
0,03-0,05 |
|
Вьюн |
544 |
44 |
10-15 |
0,7-1,5 |
0,3-3,0 |
|
Сосновка |
130 |
15 |
2,5-6,0 |
0,7-2,0 |
0,6-0,4 |
|
Семужья |
120 |
10 |
4-10,0 |
0,5-2,5 |
0,02-0,05 |
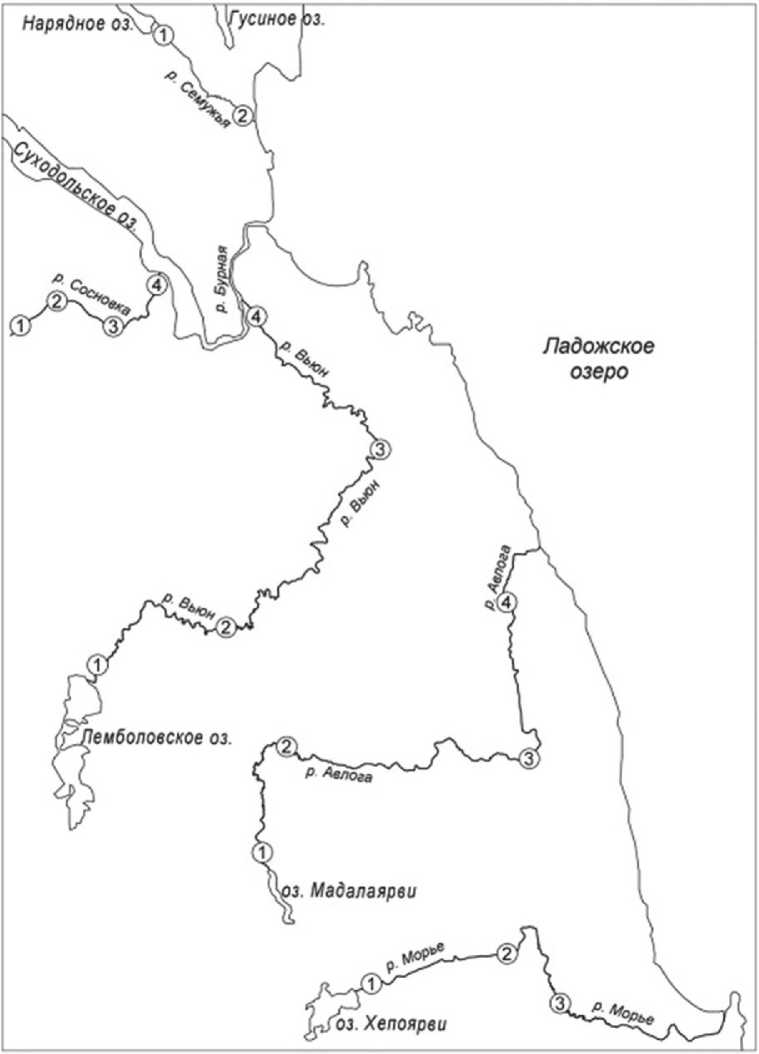
Рис. 1. Карта-схема района исследований и расположение станций отбора проб на реках
Для сбора и обработки материала использовали отработанную методику [15]. Подводные части растений срезали и помещали в 1л профильтрованной озерной воды. Затем водоросли соскабливали с помощью зубной щетки. Полученную, после фиксации 40 % формалином, взвесь водорослей использовали для определения видового состава и основных структурных характеристик эпифитона. Пробы просматривали в камере Нажжота объемом 0,05 мл с использованием микроскопов AxioLab А1 и AxioVert CFL 40 (Carl Zeiss). Для идентификации водорослей использовали отечественные и зарубежные определители и другие номенклатурные разработки.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЯ
В эпифитоне исследованных рек было выявлено 323 вида, разновидности и формы водорослей, принадлежащих к 131 роду, 64 семействам, 29 порядкам и 7 отделам (табл. 3).
По числу таксонов преобладал отдел Bacillariophyta - 122 таксона (37% от общего количества таксонов). Высокое таксономическое богатство диатомовых водорослей характерно и для большинства притоков Ладожского озера [10]. Далее по видовой насыщенности выделя- лись отделы Charophyta – 69 (22%) и Chlorophyta - 54 (17%). Отдел Cyanoprokaryota был менее разнообразен, было идентифицировано 45 таксонов, которые определяли 14% от общего списка видов. Еще менее разнообразным был отдел Euglenophyta, было выявлено 25 таксонов (8%). Отделы Xanthophyta и Rhodophyta были немногочисленными, всего было определено 5 (2%) и 3 (1%) таксонов соответственно. Подобное распределение отделов водорослей характерно для аль-гофлор обрастаний в малых и крупных водотоках разных районов умеренной зоны [16, 17, 18].
На уровне ведущих порядков выделялись Zygnematales – 20,8%, Naviculales – 9%, Chlorococcales - 8%, Euglenales - 7,5%, Cymbellales - 6,6%, Fragilariales - 5%, Oscillatoriales – 5%, Nostocales - 5% (табл. 4). Эти порядки объединяли 67 % всех обнаруженных таксонов. Обращает на себя внимание довольно высокое положение порядков Chlorococcales и Euglenales, что, скорее всего, было связано с наличием в составе эпифитона большого количества планктонных организмов.
Установлено, что при анализе флоры наибольшее значение имеют ведущие семейства, которые в большей степени отражают ее современный облик, а также факторы, ее формирующие [19]. В спектре 10 ведущих семейств
Таблица 3. Таксономическая структура и «пропорции флоры» альгофлоры эпифитона исследованных рек
|
Отдел |
порядки |
семейства |
роды |
таксоны |
Род/ сем |
Вид/ сем |
Вид/ род |
||||
|
n |
% |
n |
% |
n |
% |
n |
% |
||||
|
Cyanoprokaryota |
4 |
13,7 |
16 |
25 |
27 |
21 |
45 |
14 |
1,7 |
2,4 |
1,4 |
|
Bacillariophyta |
13 |
44,8 |
23 |
36 |
37 |
28 |
122 |
37 |
1,6 |
5,3 |
3,2 |
|
Chlorophyta |
3 |
10,4 |
10 |
16 |
28 |
21 |
54 |
17 |
2,8 |
5,4 |
1,9 |
|
Charophyta |
3 |
10,4 |
7 |
12 |
24 |
19 |
69 |
21 |
3,4 |
9,8 |
2,8 |
|
Euglenophyta |
2 |
6,9 |
2 |
4 |
8 |
6 |
25 |
8 |
4,0 |
12,5 |
3,1 |
|
Xanthophyta |
2 |
6,9 |
4 |
6 |
4 |
3 |
5 |
2 |
1,0 |
2,5 |
1,3 |
|
Rhodophyta |
2 |
6,9 |
2 |
3 |
3 |
2 |
3 |
1 |
1,5 |
1,5 |
1,0 |
|
29 |
64 |
131 |
323 |
2,1 |
4,9 |
2,4 |
|||||
Таблица 4. Спектры ведущих порядков, семейств и родов в альгофлоре эпифитона исследованных рек
Анализ родовых спектров показал, что только 5 родов из 10 ведущих входят в 50 % спектра ведущих семейств (табл. 4). К ним относились роды Closterium, Cosmarium, Navicula, Eunotia, Cymbella. Родовые спектры отражают характерные особенности водоемов конкретной территории. По разнообразию родов в эпифитоне преобладают зеленые и диатомовые водоросли. Активное участие в формировании альгофло-ры рек видов из родов Closterium, Cosmarium, Eunotia и Pinnularia связано с относительно низкой минерализацией воды в них и наличием на водосборе заболоченных территорий. Следует отметить, что из ведущего родового спектра выпадали виды семейства Fragilariaceae, которые, как правило, доминировали по биомассе в большинстве малых рек, а также в других притоках Ладоги [13].
Для получения более полной информации о таксономической структуре, исследуемой аль-гофлоры были рассчитаны «пропорции флоры», в целом они составляют 1:2,1:4,9:2,4 (табл. 3). Установлено, что высокие величины этих показателей свидетельствуют о богатстве рассматриваемой флоры [19]. Полученные нами «пропорции флоры» для альгофлоры эпифитона, по сравнению с таковыми в других водотоках, низки, что определяет относительную бедность альгофлоры рек. Общий родовой коэффициент для изученной альгофлоры составлял 2,4, тогда как для более богатых флор он может варьировать от 4 до 9 [21]. Признаками северных флор является преобладание родов с 1-2 таксонами [20]. Одно- и двувидовые роды в данной альгоф- лоре составляли 67 %, и определяли 33% общего видового состава.
Таксономическое богатство водорослей эпифитона по рекам различалось. Наибольшее количество видов отмечалось в реках Авлога и Семужья, минимальное – в р. Вьюн (табл.5). Во всех реках по количеству видов преобладали диатомовые водоросли, среди которых постоянно присутствовали Ulnaria ulna (Nitzsch) Compère, Fragilaria capucina Desm., Tabellaria flocculosa (Roth) Kütz, Meridion circulare (Greville) C. A.Agardh , Navicula cryptocephala Kütz., N. tripunctata (O.Müller) Bory , N. viridula (Kütz.) Ehr., Суmbella gracilis (Rabenhorst) Cleve, C. lanceolata (Ehr.) Kirchn., Gomphonema parvulum (Kütz.) Kütz., G . acuminatum Ehr., Eunotia pictinalis (Dillw.) Rabenh., E. incisa W.Smit ex Gregory, Nitzschia palea (Kütz.) W. Sm., N. heufl eriana Grunow , Achnanthes minutissima Kütz., Cocconeis placentula Ehr. , Frustulia saxonica (Ehr.) D.T ., Pinnularia viridis (Nitzsch.), P. microstauron (Ehr.) Cl., Melosira varians C.A. Agardh. В реках Семужьей, Морье и Авлоге отмечалось наибольшее количество видов десмидиевых водорослей, среди которых чаще всего встречались Closterium aciculare Tiffen West, C. acutum (Lyngb.) Bréb., C. leibleinii Kütz., C. setaceum Ehr., Cosmarium botrytis Menegh., C. depressum (Näg.) Lund. C . turpinii Brébisson, Euastrum dubium Ag . Количество видов зеленых водорослей во всех реках было на одном уровне (табл. 5). Повсеместно были отмечены стерильные виды рода Oedogonium из зеленых водорослей. В реках Вьюн, Сосновка, Булатная и Семужья можно было наблюдать развитие Stigeoclonium tenue (C.Ag.) Kütz., в последней реке была обнаружена Draparnaldia plumosa (Vauch.) Ag. Отмечено, что данный вид широко распространен в реках Карелии, где также может развиваться в массе [16]. Большее количество цианопрокариот было зафиксировано в реках Морье, Авлога, Булатная, в двух других реках было отмечено всего 2-5 видов. Наиболее часто встречающимися во всех реках среди цианокариот эпифитона были Heteroleibleinia kuetzingii (Schmidle) Compère, Chamaesiphon incrustans Grunow, Phormidium limosum (Dilwyn) P.C. Silva. Trachelomonas volvocina (Ehr.) Ehr., T . hispida (Perty) F.Stein, T. planctonica
Таблица 5. Таксономический состав эпифитона в исследованных реках
|
Отдел |
Bacillario-phyta |
Chlorophyta+ Charophyta |
Cyanopro-karyota |
Eugleno-phyta |
Xantho-phyta |
Rhodo-phyta |
Всего |
|
р. Авлога |
45 |
6 (12)* |
10 |
14 |
2 |
1 |
90 |
|
р. Морье |
32 |
4(18) |
13 |
7 |
1 |
2 |
77 |
|
р. Вьюн |
37 |
4 (4) |
2 |
4 |
- |
- |
51 |
|
р. Сосновка |
39 |
5(5) |
5 |
6 |
1 |
- |
61 |
|
р.Булатная |
34 |
3(2) |
11 |
3 |
1 |
- |
54 |
|
р. Семужья |
40 |
7(36) |
4 |
4 |
3 |
- |
90 |
*В скобках приведено количество видов из отдела Charophyta
Svirenko – постоянно встречающиеся виды эвгленовых водорослей. Желтозеленые водоросли – Tribonema minus Hazen встречалась в реках Морье и Семужья, а Tribonema affi ne (Kütz.) G.W. West в р. Булатной. Красные водоросли встречались в реках Морье ( Sirodotia suecica Kylin. и Batrachospermum moniliforme Roth.) и Авлоге ( Chantransia pugmaea Kütz.).
В целом, альгофлора изученных малых рек имеет сходство с альгофлорой крупных притоков и самого Ладожского озера. Доминирующие и постоянно встречающиеся виды водорослей, как правило, характерны как для малых, так и крупных притоков Ладожского озера, а также для его прибрежной акватории.
Таким образом, на основании проведенных исследований можно сделать следующее заключение:
В составе альгофлоры эпифитона малых рек было выявлено 323 таксона водорослей из 7 отделов. На основании анализа таксономического состава водорослей была выявлена относительно высокая видовая насыщенность в ведущем порядке Zygnematales, семействах Desmidiaceae, Closteriaceae, Fragilariaceae и, родах Closterium, Cosmarium , Nitzschia , Navicula, Eunotia, Cymbella. Особенностью спектра семейств является видовое богатство Desmidiaceae. Присутствие в составе эпифитона родов Eunotia и Pinnularia , несмотря на их низкие позиции в родовых спектрах, свидетельствует о формировании в аль-гофлоре «болотного комплекса» видов, который развивается в маломинерализованных, гумифицированных водах с пониженным рН. Выявлено значительное число малонасыщенных семейств и родов в альгофлоре, что подтверждает ее северный характер. В целом, альгофора малых рек относительно бедна, однако имеет достаточный потенциал для обогащения альгофлоры Ладожского озера, что способствует сохранению ее биоразнообразия.
Список литературы Таксономическая структура эпифитона малых рек северо-западного побережья Ладожского озера
- Комулайнен, С.Ф. Экология фитоперифитона малых рек Восточной Фенноскандии / С.Ф. Комулайнен – Петрозаводск, 2004. – 182 с.
- Беляева, П.Г. Структура фитоперифитонных сообществ в речных экосистемах (обзор) / П.Г. Беляева // Известия ПГПУ им. Белинского. – 2011. – № 25. – С. 484-492.
- Балахонцев, Е.Н. Ботанико-биологические исследования Ладожского озера / Е.Н. Балахонцев. – СПб., 1909. – 410 c.
- Давыдова, Н.Н. Эколого-систематическая характеристика водорослей Ладожского озера / Н.Н. Давыдова, Н.А. Петрова // Растительный мир Ладожского озера. – Л., 1969. – С. 20-56.
- Трифонова, И.С. Видовой состав и биомасса фитопланктона притоков Ладожского озера и реки Невы / И.С. Трифонова, А.Л. Афанасьева, О.А. Павлова // Ботанический журнал. – 2001. – Т. 86. – № 11. – С. 10-20.
- Рычкова, М.А. Перифитон литоральной зоны Ладожского озера / М.А. Рычкова //Литоральная зона Ладожского озера. – СПб, 2011. – С.128-138.
- Русанов, А.Г. Перифитон литоральной зоны / А.Г. Русанов // Современное состояние и проблемы антропогенной трансформации экосистемы Ладожского озера в условиях меняющегося климата. – СПб, 2021. – С. 310-316.
- Состояние биоценозов озерно-речной системы Вуоксы. – СПб, 2004. – 148 с.
- Оценка экологического состояния рек бассейна Ладожского озера по гидрохимическим показателям и структуре гидробиоценозов. – СПб, 2006. – 129 с.
- Генкал, С.И. Диатомовые водоросли планктона Ладожского озера и водоемов его бассейна / С.И. Генкал, И.С. Трифонова. – Рыбинск, 2009. – 109 с.
- Комулайнен, С.Ф. Перифитон рек Ленинградской, Мурманской областей и республики Карелия / С.Ф. Комулайнен. – Петрозаводск, 1996. – 38 с.
- Станиславская, Е.В. Водоросли перифитона озерно-речной системы Вуокса // Состояние биоценозов озерно-речной системы Вуоксы / Е.В. Станиславская. – СПб, 2004. – С. 64-72.
- Станиславская, Е.В. Структура перифитона как показатель состояния притоков Ладожского озера и реки Невы / Е.В. Станиславская // Оценка экологического состояния рек бассейна Ладожского озера по гидрохимическим показателям и структуре гидробиоценозов. – СПб, 2006. – С. 91-103.
- Станиславская, Е.В. Разнообразие водорослей перифитона в притоках Ладожского озера / Е.В. Станиславская, А.С. Горченко // Новости систематики низших растений. – 2005. – Т. 39. – С. 79-98.
- Станиславская, Е.В. Продукционная характеристика растительного перифитона / Е.В. Станиславская, И.С. Трифонова // Особенности формирования качества воды в разнотипных озерах Карельского перешейка – Л., 1984. – С. 192 – 206.
- Комулайнен, С.Ф. Альгофлора перифитона рек побережья Баренцева моря / С.Ф. Комулайнен // Труды Карельского НЦ РАН. – Петрозаводск, 2008. – Вып.14. – С.17-32.
- Горохова, О.Г. Сообщества фитобентоса соленых рек Приэльтонья: таксономическая и количественная структура / О.Г. Горохова, Т.Д. Зинченко // Поволжский экологический журнал. – 2020. – № 4. – С.383-397. – DOI: https://doi.org/10.35885/1684-7318-2020-4-383-397
- Беляева, П.Г. Структура и функционирование альгоценозов водных экосистем Пермского Предуралья: Автореф.дис… докт.биол.наук / П.Г. Беляева. – Пермь, 2022. – 45 с.
- Шмидт, В.М. Статистические методы в сравнительной флористике / В.М. Шмидт. – Л., 1980. – 176 с.
- Гецен, М.В. Водоросли в экосистемах Крайнего Севера / М.В. Гецен. – Л., 1985. – 168 с.
- Яценко-Степанова, Т.Н. Эколого-структурный анализ альгофлоры Оренбуржья / Т.Н. Яценко-Степанова, Н.В. Немцева, М.Е. Муравьева // Вестник ОГУ. – 2005. – Т. 12. – С.66-71.
