Таксономическое положение Palaeoa plysina krotov, 1888

Автор: Пономаренко Е.С.
Журнал: Вестник геонаук @vestnik-geo
Рубрика: Научные статьи
Статья в выпуске: 2 (218), 2013 года.
Бесплатный доступ
Проблематичные организмы Palaeoaplysina впервые описаны П. И. Кротовым в 1888 г. С тех пор их таксономическое положение не раз пересматривалось. В разное время палеоаплизин относили к губкам, гидрозоям, зеленым кодиевым или красным водорослям. Обнаруженные гидранты в микробиальной массе на поверхности скелета свидетельствуют о том, что палеоаплизины были гидрозоями. В палеоценозах бентосных сообществ палеоаплизины слагают верхнюю (консументную) часть трофической пирамиды, что говорит о мезотрофности окружающих вод.
Проблематичные организмы, гидрозои, рифогенные палеосообщества, трофические уровни
Короткий адрес: https://sciup.org/149129095
IDR: 149129095
A taxonomic position of Palaeoa plysina krotov, 1888
An enigmatic organisms Palaeoaplysina were first described by P. I. Krotov at the 1888 year. Since then their taxonomic position repeatedly have been revised. At variously times Palaeoaplysina were refered to sponges, hydrozoans, green codiacean algae and red algae. Hydrants were founded in a microbial matter on a surface of skeletons show the Hydrozoan origin. The Palaeoaplysina composed an upper (consumer) part benthic communities of trophic pyramid. It had indicated mesotrophic surrounding waters.
Текст научной статьи Таксономическое положение Palaeoa plysina krotov, 1888
Органические остатки неясного систематического положения широко распространены в верхнекамен-ноугольно-нижнепермских карбонатных отложениях северной гемисферы [10]. Остатки этих организмов часто ассоциируются с нижнепермскими рифогенными образованиями. Несмотря на широкий стратиграфический диапазон развития Palaeoaplysina — от московского яруса среднего карбона [16] до артинс-кого яруса нижней перми [9], все эти организмы определяются только как Palaeoaplysina laminaeformis Krotov, 1888. Ископаемые остатки палеоа-плизин представляют собой пластины (иногда до 1.0 м длиной), имеющие нижнюю ровную и верхнюю бугорчатую поверхности, между которыми развита ячеистая ткань с системой каналов (рис. 1, А). Каждый из этих элементов строения (ячеистая ткань, система каналов) могут быть характерны для разных организмов [15], в том числе представителей разных царств. Палеоаплизины были широко распространены в нижнепермских органогенных сооружениях типа скелетных холмов в Тимано-Североуральском регионе [1] и являлись важным компонентом трофической структуры рифовой экосистемы в то время, но их неясное систематическое положение не позволяет точно установить их роль в пищевом круговороте раннепермских экосистем. Целью работы является
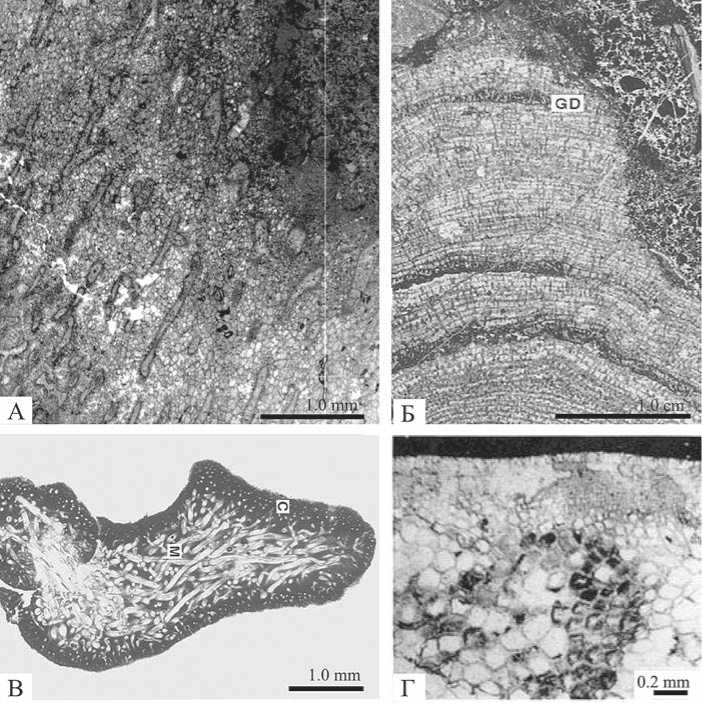
Рис. 1. Palaeoaplysina и возможные варианты их родства. A: Palaeoaplysina Krotov из нижнеассельских отложений р. Унья. Хорошо сохранилась сетчатая ткань между системой каналов. Продольный срез. Обр. П-Ун28/115-2009. Б: Строматопороидея Actinostroma papillosum (Bargatzky). Верхний девон, франский ярус, бассейн Каннинг (Canning Basin), Западная Австралия [по: 15]. В: Таллом зеленой кодиевой водоросли Halimeda . С — внешняя корковая зона, М — центральная зона (медула). Современная рифовая лагуна, Флорида Бэй (Florida Bay), США [по: 15]. Г: Фрагмент гипоталлома и периталлома красной водоросли Archaeolithophyllum missouriense Johnson. Карбон (пенсильваний, известняк Кэптиан Крик (Captian Creek Limestone), округ Уилсон (Wilson County), Канзас [по: 18]
уточнение природы Palaeoaplysina , что имеет принципиальное значение для палеоэкологических построений и определения положения этих организмов в структуре рифовых сообществ.
Эволюция взглядов на систематическое положение палеоаплизин
Название «палеоаплизина» появилось в палеонтологической литературе в 1888 г., когда П. И. Кротов дал описание новой «роговой губки» — Palaeoaplysina laminaeformis Krotov, 1888. Описание сопровождалось рисунками, показывающими внешний вид этих организмов, внутреннее строение ткани с многочисленными мелкими каналами и строение верхней и нижней поверхностей. По сходству с современными роговыми губками Aplysina aerophoda Nardo П. И. Кротов отнес найденные формы к Ceratospongiae [4]. В 1895 г. А. А. Штукенберг описал новый род и вид Mezenia roseni Stuck., который он отнес к новому роду класса гидромедуз [8]. В 1915 г. В. Н. Рябинин описал из каменноугольных отложений Урала и Тимана тонкие пластинки и корки Uralotimania reticulate Riab. и отнес эти остатки к классу Hydrozoa отряда Tubulariae. Позднее автор пришел к выводу, что указанные три рода ( Palaeoaplysina Krot., Mezenia Stuck. и Uralotimania Riab.) являются одним и тем же родом, который был впервые описан П. И. Кротовым [8]. Работами Б. И. Чувашова [9] было показано фациальное распространение палеоаплизин: в ассоциации с эвапоритами (на восточной окраине Русской платформы), в активноводных обстановках в ассоциации с колониальными кораллами и в би-огермных отложениях. Были установлены основные закономерности ячеистой ткани между каналами: трех-, пяти- и четырехугольные ячейки по мере приближения к каналам становятся мельче, и возле каждого канала имеется более или менее широкая зона, сложенная мелкоячеистой тканью, а наиболее крупные ячейки расположены на половине расстояния между каналами. Палеоаплизины были отнесены к строматопороидеям [9]. Долгое время строматопоры относились к гидрозоям, однако сейчас они включены в тип Spongia (рис. 1, Б). Важным признаком такого таксономического положения строматопорат явля- 22
ется сходство астрориз и общей модели скелета (skeletal pattern) с современными склероспонгиями [15]. Высказывалось мнение, что палеоа-плизины являются древними аналогами современных зеленых кодиевых водорослей Halimeda [14]. Авторы отметили, что каналы палеоапли-зин похожи на нити кодиевых морских водорослей Halimeda (рис. 1, В). Сходство форм палеоаплизин с ко-диевыми водорослями из биогермов о-ва Ява отмечали Уоткинс и Уилсон [17]. В последних публикациях палеоаплизины рассматриваются как ассоциация проблематичных трубчатых организмов (система каналов) и красных водорослей (сетчатая ткань) Palaeoaplysinella [13, 16], аналогичных красным водорослям Archaeolithophyllum (рис. 1, Г). Таким образом, имеются четыре точки зрения на природу Palaeoaplysina: как губок, гидрозой, зеленых или красных водорослей.
Таксономическое положение палеоаплизин
Принятие той или иной точки зрения может кардинально изменить палеоэкологическую характеристику рифовых сообществ. Для разрешения данного вопроса в первую очередь необходимо было выяснить, к какому царству живой природы принадлежат палеоаплизины. Нужный результат был получен с помощью метода электронного парамагнитного резонанса (ЭПР). В спектрах ЭПР, по данным Ф. А. Муравьева и его соавторов [6], присутствует пик, положение которого отвечает наличию в пробе радикалов животного или растительного ряда. Исследования методом ЭПР скелетной ткани палео-аплизин выявили свободные радикалы на фрагментах поликонденси-рованных ароматических молекул, характерных для включений в карбонаты морской фауны на начальных
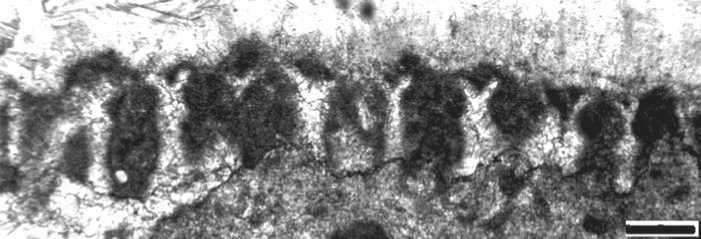
Рис. 2. Остатки гидрантов на поверхности палеоаплизины, сохраненные благодаря микробиальной инкрустации (темное). Шлиф П-Ил41/2-2008. Ассельский ярус, холодноложский горизонт; р. Илыч, Северный Урал. Масштаб — 0.1 мм
стадиях углефикации [5]. Эти данные дают полное основание рассматривать палеоаплизин в качестве животных — представителей консумен-тной части трофической структуры рифовой экосистемы нижнепермских скелетных холмов.
Однако этих данных недостаточно для точной диагностики их трофического положения, поскольку губки являются фильтраторами, а гидрозои — хищниками. Если отнести палеоаплизин к гидрозоям, то необходимо доказать существование мягких гидрантов (питающих зооидов). Сохранение зооидов могло происходить только в случае инкрустации их другими организмами. Прекрасным примером тому служат палеозойские гидроиды Fistulella , мягкие кроновые части которых сохранились благодаря инкрустации цианобактериями Ikella , что позволило выделить у них простые питающие полипы (гидранты) и бластостили с гонофорами [11].
В рифогенных образованиях нижней перми Palaeoaplysina часто заключены в синседиментаци-онный биологически индуцированный цемент [2, 5], который образовался после смерти палеоаплизин и не мог сохранять мягкие ткани. Прижизненные микробиальные нарастания цианобактерий и проблематичных организмов Tubiphytes на па-леоаплизинах — явление довольно редкое, поэтому были проведены специальные исследования, направленные на поиски возможных остатков гидрантов в микробиальных сообществах, нарастающих на пластины Palaeoaplysina . Уникальные образцы встречены в ассельских органогенных постройках р. Илыч и руч. Тягла (междуречье Печоры и Уньи) на западном склоне Северного Урала. В них в области темных микробиальных образований были обнаружены необычные структуры, видимые
в шлифах как полости размером от (0.01—0.05) х 0.1 до 0.1 х 0.5 мм, заполненные спаритом. Полости и мелкие поры на поверхности скелета палеоаплизин часто соединены. Этот факт и позволил интерпретировать эти своеобразные структуры как отпечатки гидрантов палеоаплизин, сохранившихся в микробиальной массе (рис. 2). Такая интерпретация позволяет с большой вероятностью отнести палеоаплизин к гидрозоям, так как губки не имеют гидрантов.
Палеоаплизины в трофической структуре экосистемы нижнепермских скелетных холмов
Анализ сукцессии нижнепермских скелетных холмов показал, что палеоаплизины чаще всего отмечаются в отложениях стадии деструкции, когда постройка достигала активных, хорошо аэрируемых вод [7]. Палеоэкологический анализ палеоценозов в постройках выявил сложную трофическую структуру из четырех уровней [12], где палеоапли-зины как гидрозои-хищники должны быть на верхнем уровне пищевой пирамиды. На стадии деструкции в составе палеоценозов в основном встречаются палеоаплизины в био-цементолитах с незначительным развитием фораминифер (фильтраторы) и цианобактерий (продуценты — хе-мо- и фототрофы). Микробиальные пленки, продуцирующие крустифи-кационный цемент, по всей видимости, являлись организмами-редуцентами (микроорганизмы — бактерии и грибы, разрушавшие отмершие остатки организмов и превращавшие их в неорганические и простейшие органические соединения). Малая доля участия автотрофных организмов в ассоциации с палеоаплизина-ми свидетельствует о мезотрофно-сти окружающих вод. Современные гидрозои питаются мелким планктоном, личинками рыб и ракообразных (остракод). Одним из важных источников питания для Paleoaplysina могли быть зеленые планктонные водоросли Tasmanites, обнаруженные во вмещающих конденсированных отложениях [3].
Проведенные методом ЭПР исследования показали, что в скелетах палеоаплизин присутствуют свободные радикалы на фрагментах поли-конденсированных ароматических молекул, характерных для включений в карбонаты морской фау- ны на начальных стадиях углефикации, что позволило отнести па-леоаплизин к царству животных. Микроскопические исследования возможных остатков гидрантов в микробиальных сообществах, нарастающих на пластины Palaeoaplysina, позволили обнаружить отпечатки питающих зооидов палеоапли-зин, указывающих на систематическое положение этих организмов в классе Hydrozoa. Палеоаплизины как гидрозои-хищники представляют верхнюю часть пищевой пирамиды. Малая доля участия автотрофных организмов основания трофической системы свидетельствует о ме-зотрофности окружающих вод.
Исследования поддержаны Программой Президиума РАН №28/2 в рамках проекта №12-П-5-1006и грантом №12-05-31402.
Список литературы Таксономическое положение Palaeoa plysina krotov, 1888
- Антошкина А. И. Рифообразование в палеозое (на примере севера Урала и сопредельных территорий). Екатеринбург: УрО РАН, 2003. 303 с.
- Антошкина А. И. Биоцементолиты - важный компонент органогенных построек позднего карбона-ранней перми (на примере севера Урала) // Верхний палеозой Роси: стратиграфия и фациальный анализ: Материалы II Всеросийской конференции. Казань: Казан. гос. ун-т, 2009. С. 42-43.
- Горожанина Е. Н., Горожанин В. М., Ефимов А. Г. и др. Депрессионые отложения ранней перми юга Предуральского прогиба (по материалам глубокого бурения) // Верхний палеозой Роси: стратиграфия и фациальный анализ: Материалы II Всеросийской конференции. Казань: Казан. гос. ун-т, 2009. С. 176- 177.
- Кротов П. И. Геологические исследования на западном склоне Чердынского и Соликамского Урала // Тр. Геологического комитета. СПб., 1888. Т. XIII. С. 297-537.
- Лютоев В. П., Антошкина А. И., Пономаренко Е. С. Парамагнитные маркеры стадийности отложения карбонатов палеоаплизиновых биоцементолитов // Рифы и карбонатные псефитолиты: Материалы Всерос. литолог. совещ. Сыктывкар: Геопринт, 2010. С. 205- 207.
