Техногеннные грунты – современная среда обитания микроорганизмов (по результатам сканирующего электронного микроскопирования)
")
Автор: Артамонова В.С., Шавекина А.Ш.
Журнал: Антропогенная трансформация природной среды @atps-psu
Рубрика: Трансформация природной среды
Статья в выпуске: 1 т.11, 2025 года.
Бесплатный доступ
В статье рассматриваются экологические проблемы, существующие в районах добычи и переработки полезных ископаемых в Сибирском регионе. Приводится информация об освоении техногенных отходов рудообогащения – техногенных грунтов (ТГ) микроорганизмами, что влечёт за собой негативные и позитивные последствия. Сообщается, что в колонизации и преобразовании поверхностной толщи многолетних хранилищ сульфидных и сульфоарсенидных отходов рудообогащения, складированных в прошлом веке на дневной поверхности, в гумидной зоне Западной Сибири и в аридной зоне Южной Сибири, участвуют фотогетеротрофные микроорганизмы. Электронное микроскопирование проб техногенных субстратов свидетельствует о присутствии в них метаболически активных цианобактерий и диатомовых водорослей, характерных для первых стадий восстановительных сукцессий наземных экосистем. Сохранение микроорганизмов в жизнедеятельном состоянии обеспечивается колониальным образом жизни одноклеточных и нитчатых особей, продуцированием ими обильной слизи. Отмечается, что химический состав микроорганизмов, определённый с помощью микрозондового (спектрального) анализа элементов, включает тяжёлые металлы и неметаллы. Сообщается, что в крайне экстремальных условиях обитания в слизи и цитозоле цианобактерий в значительном количестве присутствует мышьяк. Предполагается, что он используется в аноксигенном фотосинтезе для получения дополнительной энергии и азотфиксации. Диатомовые водоросли продуцируют слизь, но не участвуют в иммобилизации мышьяка, что, возможно, обусловлено механизмами биохимической защиты. Приводятся сведения о накоплении в среде обитания цианобактерий и диатомей кристаллических форм минералов – барита в гумидной зоне и мышьяковистых минералов – в аридной зоне. Зарождение минералов могло быть вызвано испарительным эффектом и микробиологическими процессами. Полученная информация может быть использована для характеристики ранних стадий почвообразования в экстремальных местообитаниях техногенных ландшафтов, учтена при создании природоподобных технологий эффективной биоремедиации загрязнённых почв, разработке экологически безопасных способов консервации грунтов, при моделировании процессов образования аутигенных минералов в разных природных условиях.
Почвообразование, диатомеи, цианобактерии, сканирующая электронная микроскопия
Короткий адрес: https://sciup.org/147250860
IDR: 147250860 | УДК: 504.75+691.4; 504.4.54 | DOI: 10.17072/2410-8553-2025-1-23-40
Technogenic grounds are a modern habitat for microorganisms (according to the results of scanning electron microscopy)
The article examines the environmental problems existing in the mining and processing areas of the Siberian region. Information is provided on the development of man-made ore enrichment waste – man-made soils (TG) by mi-croorganisms, which entails negative and positive consequences. It is reported that photoheterotrophic microorganisms are involved in the colonization and transformation of the surface layer of long-term storage facilities for sulfide and sulfoarsenide ore enrichment waste stored in the last century on the daytime surface, in the humid zone of Western Siberia and in the arid zone of Southern Siberia. Electron microscopy of samples of technogenic substrates indicates the presence of metabolically active cyanobacteria and diatoms in them, characteristic of the first stages of regenerative successions of terrestrial ecosystems. The preservation of microorganisms in a vital state is ensured by the colonial lifestyle of unicellular and filamentous individuals, producing abundant mucus. It is noted that the chemical composition of microorganisms, determined using microprobe (spectral) analysis of elements, includes heavy metals and non-metals. It is reported that arsenic is present in significant amounts in the mucus and cytosol of cyanobacteria in extremely extreme living conditions. It is assumed that it is used in anoxygenic photosynthesis to obtain additional energy and nitrogen fixation. Diatoms also produce mucus, but are not involved in the immobilization of arsenic, which is probably due to the mechanisms of bio-chemical protection. Information is provided on the accumulation of crystalline forms of barite in the humid zone and arsenic minerals in the arid zone in the habitat of cyanobacteria and diatoms. The origin of minerals could be caused by the evaporation effect and microbiological processes. The information obtained can be used to characterize the early stages of soil formation in extreme habitats of man-made landscapes, taken into account when creating nature-like tech-nologies for effective bioremediation of contaminated soils, developing environmentally safe methods of soil conserva-tion, and modeling the processes of autogenic formation of minerals in different natural conditions.
Текст научной статьи Техногеннные грунты – современная среда обитания микроорганизмов (по результатам сканирующего электронного микроскопирования)
Микроорганизмы, будучи древнейшими организмами на нашей планете, существующими миллиарды лет, являются самыми древними почвообразовате-лями, действовавшими задолго до появления высшей растительности [16]. Их участие в современных процессах почвообразования на обнажённых минеральных субстратах и техногенных грунтах (далее – ТГ) предопределено былыми эпохами. Доказано, что микроорганизмы быстро заселяют вскрышные и вмещающие породы районов угледобычи в Кузбассе и на КАТЭКе (Россия, Западная и Средняя Сибирь), поскольку они не токсичны, представлены лессовидными карбонатными суглинками с примесью алевролитов, песчаников, аргиллитов. Выступая в роли почвообразующих пород, они активно вовлекаются в биогенное преобразование и к 20-25 – летнему сроку самозарас-тания приобретают признаки биогенности, гуматно-сти, профилирования, характерные для молодых (первичных, примитивных, биогенно слаборазвитых почв – эмбриозёмов) [9]. Таким образом, такие техногенные грунты вовлекаются в процессы биогеохимического преобразования с трендом на восстановление утраченных экологических функций, свойственных почвам. Для ускорения их возобновления используются различные приёмы биологической рекультивации, направленные на приближение территорий к исходному состоянию, и предусматривающие возможное целевое использование: лесохозяйственное, рекреационное, в том числе туристическое, и другие.
Другая группа ТГ, представленная сульфидными и сульфо-мышьяковистыми техногенными отходами, возникшими в процессе обогащения полиметаллических руд под воздействием высоких температур и агрессивных технологических реагентов (кислот, щелочей, цианидов, ртути), чрезвычайно проблематична в отношении вовлечения в процессы гипергенеза и первичного (биогенного) почвообразования. Изначально такие объекты биологически стерильны и токсичны, что вызывает много дискуссий в отношения возобновления в них жизни и её роли в судьбе ТГ. Тем не менее, многолетнее нахождение ТГ в окружении природных экосистем (50-100 лет) сопровождается заселением их местными представителями педомикро-биоты. Так, в отходах цианирования сульфидных Au-содержащих руд методом метагеномики выявлен пул жизнеспособных ацидофильных бактерий [22], методом обрастания мелкозёма – диссоцианты азотфикси-рующей бактерии рода Azotobacter [1]. С помощью сканирующего электронного микроскопа (далее – СЭМ) в многолетних отходах выщелачивания полиметаллических руд цианидами в гумидной зоне и амми-ачно-карбонатными растворами – в аридной зоне обнаружены метаболически активные цианобактерии и диатомеи [2]. Можно предположить, что в отдалённом будущем биогенно освоенные ТГ эволюционируют в почвоподобные твёрдофазные тела, как бы почвы – техногенные поверхностные образования (ТПО) группы квазизёмов. В Классификации почв России они частично соответствуют токсифабрикатам. Но этот процесс занимает неопределённо долгое время, отчего ТГ представляют собой современный объект, имитирующий древнее почвообразования на суше Земли и его возможное присутствие на других планетах. Как продуктивный земельный ресурс они не представляют интерес из-за присутствия высоких концентраций металлов и металлоидов (Au, Ag, Zn, Pb, As и другие), вторичного образования минералов, например, барита и мышьяковистых минералов, в образовании которых могут участвовать микроорганизмы. Барий не относится к жизненно важным элементам для биоты, но барит является стратегически значимым минеральным сырьём, отчего осаждение барита с участием микроорганизмов в последние годы оказалось в центре внимания научного сообщества. Наряду с этим, ионы бария, барит, как и мышьяковистые соединения в той или иной степени биотоксичны. Но микроорганизмы сохраняют свою жизнедеятельность в их окружении. Более того, они участвуют в их иммобилизации и осаждении, что представляет фундаментальный и прикладной интерес к такой способности микроскопического населения токсичных ТГ.
Нужно отметить, что в текущем веке интерес к ТГ резко активизировался в экологическом и микробиологическом отношении. Во-первых, в приземном слое воздуха над отходами регистрируются летучие углерод-, азот-, серосодержащие органические соединения [28]. Эмиссия метилированных форм диметилсуль-фида (далее – ДМС) с многолетних хранилищ отвалов сульфидсодержащих отходов Урского месторождения (Западная Сибирь) достигает 420 мкг/м3. Факты образования ДМС были выявлены ранее на поверхности микробных матов морских экосистем [45]. Предполагается, что ДМС образуется в результате реакции фотосинтетически полученного низкомолекулярного органического углерода и биогенного сероводорода, полученного в результате восстановления сульфатов. Существуют и другие версии, и подтверждения участия микроорганизмов в формировании ДМС, который, в свою очередь, может влиять на образование облаков, прежде всего над океанами, где продукция метаболита фитопланктоном наиболее велика. Эти процессы связываются с изменением климата Земли и химическим составом экзопланет Вселенной.
Отмечается, что ДМС образуется только в результате жизнедеятельности. Поэтому метаболит является своеобразным маркером присутствия жизни на других планетах. В 2023 г. американскими учёными НАСА на экзопланете K2-18b обнаружена молекула под названием диметилсульфид, на основании чего высказано предположение о существовании присутствия углеродсодержащих молекул, в том числе метана и углекислого газа в присутствии аммиака, что связывается с наличием водного океана под атмосферой богатой водородом. Планета в 8,6 раза массивнее Земли, вращается вокруг холодной карликовой звезды K2-18, находится в 120 световых годах от Земли [47].
Другой не менее значимый факт влияния микроорганизмов на экологию техногенных ландшафтов – это участие их в образовании летучей ртути. Высокое содержание металла обнаружено над хранилищами сульфидных отходов переработки золото-полиметаллических руд и за их пределами их складирования в Западной Сибири [10, 12]. Доказано, что количество ртути значительно усиливается в присутствии тиосульфата, который выполняет двойную роль, повышая растворение ртути за счет комплексообразования металла и обеспечивая дополнительный метаболический субстрат для тионовых бактерий [44]. Проявление сопряжённого абиотически-биотического механизма превращения Hg (II) в Hg (0) в сульфидных субстратах с участием нейтрофильных хемосинтезирующих бактерий не исключено в сульфидных отходах рудообога-щения Сибири. Содержащаяся в минеральном суб- страте ртуть может представлять собой недооцененный источник газообразной элементарной ртути, которая мигрирует в окружающую среду, усугубляя уже существующий набор экологических проблем в районах активного функционирования предприятий горнодобывающей и металлургической промышленности. Роль микробной компоненты в образовании летучей ртути в ТГ пока до конца не изучена.
Остаются открытыми вопросы участия микроорганизмов в миграции мышьяка. Как известно, биогеохимия мышьяка сложна и включает в себя различные процессы адсорбции и десорбции. В окислительных условиях мышьяк может высвобождаться из пирита или оксидов железа, особенно при повышенном уровне pH, что имеет место в ТГ, особенно в аридной зоне. Проблема заключается в том, что в отходах рудо-переработки, складированных в карты хранения на территории Республики Тыва, часть мышьяка (до 20 %) представлена в форме арсенидов металлов, не разложившихся в процессе выщелачивания [14]. Теоретически мышьяк должен был связаться с аммиачно-кар-бонатным комплексом и перейти в нерастворимое состояние, но в реальности существенная часть соединений мышьяка оказалась представленной его 3-х валентной формой, которая образует с водой растворимые высокотоксичные соединения. Более того, повышенные содержания мышьяка в поровых растворах шламов присущи верхним горизонтам хранилищ. По этой причине мышьяк особенно опасен в период снеготаяния и обильных дождей, поскольку транспортируется за пределы карт хранения отходов в сторону расположения водотока и населенных пунктов.
Считается, что микроорганизмы являются основными движущими силами, которые приводят к преобразованию различных форм мышьяка [ 48].
Трансформационные свойства As включают окисление-восстановление, метилирование и деметилирование, органическое хелатирование, поверхностную адсорбцию и диссоциацию, а также соосаждение ионов. Восстановление, деметилирование органического мышьяка и диссоциация адсорбированного As обра-зуют высокотоксичный As(III), что приводит к по-вышению подвижности и токсичности элемента, в то время как окисление, метилирование, органическое связывание и соосождение мышьяка образуют ме-нее токсичные/подвижные арсенаты, летучий ме-тилмы-шьяк (например, ДМА и ТМА), и остаточный сульфид As. Биоулетучивание мышьяка привлекает внимание исследователей с целью углубления знаний в области биогеохимии и охраны окружающей среды, использования процесса в биоремедиации, в том числе с участием микроорганизмов [46].
Сообщается, что почти все микроорганизмы обладают геном устойчивости к мышьяку [49]. Некоторые виды бактерий используют мышьяк для энергетических нужд, используя арсенаты в качестве доноров электронов и производя арсенаты. Существует мнение, что на протяжении истории фотосинтезирующие организмы производили арсенаты, которые позволяли бактериям, восстанавливающим их, выживать.
В этой связи, исследования метаболических реакций микроорганизмов-первопоселенцев ТГ «техноген- ных месторождений» [6], заслуживают особого внимания. Их роль в проявлении негативных и позитивных последствий колонизации ТГ изучена недостаточно. Учитывая неизбежный рост потерь почвенно-растительных покровов в ходе добычи и переработки полезных ископаемых, особенно полиметаллических руд, метаболическую активность микроскопического населения ТГ следует принимать во внимание.
К настоящему времени суммарный удельный вес в процессе почвенных потерь с участием горнодобывающей и металлургической промышленности достиг 80 % от почти 3 тыс. га земель, нарушаемых ежегодно в Сибири [4]. Это объясняется высоким спросом на минеральное сырьё. При этом производство тонны черного металла сопровождается получением до 17 т отходов, цветных и благородных металлов – до 100 т [15], что требует постоянно новых земель отчуждения под их складирование.
В России общая площадь таких земель, уже занятых шламонакопителями, хвостохранилищами, насыпными отвалами отходов, достигла 1 тыс. км2. В них содержатся в высокой концентрации цветные металлы, тяжёлые (Cu, Zn, Pb, Ni, Sn, Cr, Нg) и драгоценные (Au, Ag, Pt), а также полуметаллы (металлоиды): Si, As, Sb, B, Ge, Te. Наблюдается новообразование минералов Pb, Cu, Fe [25], вторичное золотое обогащение [26]. Отмечается, что площадь, подверженная техногенному загрязнению, в 10-15 раз превышает площадь, занимаемую самими ТГ. В Западной Сибири (Кемеровская обл.) ветровое загрязнение пылевыми наносами зарегистрировано в пределах 5-километровой зоны от расположения отходов Салаирского обогатительного комбината [5]. На прилегающую к хвостохранилищу территорию выносится более 3000 т пыли в год. В Южной Сибири мышьяк регистрируется в почвах в количествах, превышающих ПДК вокруг локаций хранения отходов (Хову-Аксинское месторождение) [20], в донных отложениях р. Енисей – на расстоянии 160 км от хранилищ [14]. Загрязнение компонентов природной среды в ходе транзита химических элементов с воздушными и водными потоками, в том числе с паводковыми, дражными и грунтовыми водами, обусловливает снижение запаса ценных минеральных ресурсов и в самих хранилищах ТГ.
Целесообразность вовлечения отходов производства полиметаллических концентратов в хозяйственный оборот доказана многолетней практикой во многих странах мира. Например, в сырьевом балансе США и Японии они достигают 26%, у большинства экономически развитых стран Европы этот показатель колеблется в пределах 16-20%, в СССР он составлял 15%, в современной России – не превышает 10% [15].
Ослабление негативных последствий долговременного нахождения ТГ на дневной поверхности ожидается в результате применения природоподобных технологий, необходимость развития которых высказана в 2023 году Президентом РФ [23]. При разработке таких технологий необходимо учитывать активность микроорганизмов, которые вовлекаются в биогеохимические процессы, в том числе тех, которые способствуют восстановлению или поддержанию механиз- мов самоочищения и адаптации любой живой системы. Для природоподобных технологий, в том числе ориентированных на создание защитных систем с применением почвоподобной компоненты, где могут проявляться признаки почвоподобия и, следовательно, биогенные свойства, могут быть использованы аборигенные виды микроорганизмов, способные проявлять детоксикационную, аккумуляционную, цементирующую способность. Такие свойства микроорганизмов используются в биоремедиации и биогеотехнологиях металлов, в частности, при их извлечении микробно-индуцированным осаждением.
Микробно-индуцированное осаждение карбонатов кальция активно внедряется в практику закрепления техногенных песков в дальнем зарубежье [23, 31, 37, 38, 50]. В России микробно-индуцированное осаждение кальция рекомендовано для защиты песков в резко континентальном аридном климате Южного Приа-ралья [17], для цементирования техногенных грунтов в Пермском крае [24]. Ранее, на Украине, для укрепления техногенных песков отходов агломерации железной руды использовали водоросли [27].
Применительно к ТГ, складированным в прошлом веке, целесообразно использование местных видов цианобактерий и диатомей, в качестве одного из природных компонентов природоподобных технологий защиты поверхности. Эти виды уже прошли предадапта-цию к повышенным концентрациям токсичных элементов благодаря развитию в экосистемах, расположенных в пределах исходных месторождений. Не исключено, что цианобактерии реализуют, сохранившийся с давних времён аноксигенный фотосинтез в случае избытка в среде обитания серы и мышьяка, цианобактерии и диатомовые водоросли – гетеротрофизм на фоне дефицита органических соединений углерода, диатомеи – синтез и полный гидролиз мочевины, обеспечивающий им восполнение энергии и подщелачивание среды, окружающей клетки. Однако многие аспекты развития фотогетеротрофных микроорганизмов в ТГ отходах неприродного происхождения остаются не раскрытыми. Процесс формирования биогенности почвы чрезвычайно длителен и соизмерим с геологическими периодами, это очень уязвимый природный ресурс – настоящая «шагреневая кожа» планеты [19]. Поэтому отбор микроорганизмов для эффективных природоподобных технологий экранирования ТГ не прост, он неизбежно должен учитывать особенности развития микроорганизмов ранних этапов первичного почвообразования, протекающего в экстремальных условиях.
Наши сегодняшние и будущие комплексные работы направлены на изучение биогеохимической обстановки в ТГ неприродного генезиса. Цель данной работы – представить новые сведения о жизнедеятельности цианобактерий и диатомей in situ в экстремальных местообитаниях техногенных ландшафтов, а также информацию об аккумулировании ими химических элементов, в том числе токсичных (на примере техногенных грунтов гумидной и аридной зоны Сибири, с применением СЭМ).
Материалы и методы
Объектами исследований были ТГ, представляющие собой многолетние отходы рудообогащения, расположенные в разных природных зонах и отличающиеся по происхождению. Одна группа ТГ сформирована в прошлом веке отходами цианирования сульфидных золото-полиметаллической специализации руд в пределах Западной Сибири (Кемеровская обл.) в гумидной зоне: Урское рудное поле (Ново-Урское месторождение) и Салаирское рудное поле (месторождение Тал-мовские Пески). Другая группа ТГ представлена отходами выщелачивания аммиачно-карбонатными растворами сульфоарсенидных руд, складированных в карты хранения в пределах Хову-Аксинского месторождения в аридной зоне Южной Сибири (Республика Тыва). Все объекты не заселены высшей растительностью, внешне представляют собой безжизненные минеральные субстраты.
Пробы для микробиологического и минералогического анализов отбирали с поверхности ТГ летом 2021 г. Они представляли собой монолиты размером 3 х 4 х 2 см. Затем в лабораторных условиях пробы заливали эпоксидной смолой, выполняли приполировки алмазными пастами по аналогии с аншлифами для последующего просмотра с применением СЭМ ИНГГ СО РАН. СЭМ приобретён в фирме TESCAN MIRA 3LMU (Tescan, Чехия). Его энергетическим спектрометр произведён в фирме OXFORD (Oxford Instruments, Великобритания), функционирует в режимах вторичных и обратно-рассеянных электронов при различных увеличениях (ускоряющее напряжение электронного пучка 20 кВ, ток зонда 1,5 нА, время набора спектров – 20 с, размер пучка – 1 мкм). Наше рабочее увеличение объектов не превышало 1500 раз.
Визуализация минералов и жизнедеятельных особей сопровождалась определением в них содержания химических элементов. Для этого использовали установку ОХА-8230 (Jeol Ltd, Япония) с ускоряющим напряжением 20 кВ и током микрозонда 70 нА, (мик-рорентгеновского спектра). Результаты содержания элементов (в массовых, %) фиксировали на экране монитора. Использованная техника занимает лидирующее положение в геологии, медицине, гидробиологии, палеомикробиологии, почвоведении и других науках. Следует сказать, что в текущем году исполняется 110 лет, как метод шлифов был впервые предложен Б.Б. Полыновым в почвоведении для определения минералогического состава почв. Широкое применение метод нашёл после организации в 1944 г. лаборатории минералогии почв в Почвенном институте АН СССР.
Результаты и обсуждение
Исследования показали, что все пробы ТГ, представляющих собой отходы выщелачивания сульфидных полиметаллических руд цианидами в гумидной зоне и сульфоарсенидных руд аммиачно-карбонат-ными растворами – в аридной зоне, обитаемы. В них присутствуют жизнедеятельные цианобактерии и диатомеи. Так, в ТГ гумидной зоны цианобактерии представлены колониями, которые образуются многокле- точными нитевидными особями, трихомы которых плотно прилегают друг к другу, образуя прочные группировки особей. Они представлены видами родов Microcoleus, Phormidium. Такие колонии принадлежат видам с узкими трихомами, в них идентифицируются гормогонии (рис. 1 / fig. 1), способные осваивать новые микрониши. Тяжи нитей обладают механическими свойствами, обычно проявляя себя как вязкоупругие материалы. Однако до сих пор неясно, каким образом нити самоорганизуются в коллективные структуры.
Другая группа колоний цианобактерий представлена многоклеточными нитевидными особями, но объединёнными слизистыми экзометаболитами. В этой группе колонистов присутствуют структуры с общей слизью, например, представленные особями Schizothrix (рис. 2 / fig. 2), а также особи с индивидуальным продуцированием слизи, которая прочно соединяет особи друг с другом в колонии (рис. 3 / fig. 3). Колониальный образ жизни в экстремальных условиях имеет определённую выгоду, особи в колониях наиболее защищены от внешнего воздействия, а также избытка ультрафиолета и других абиотических факторов. Слизистые метаболиты способствуют движению особей и их гормогониев по поверхности минеральных частиц путём скольжения. Предполагается, что скольжение зависит от движущей силы, создаваемой сообществом клеток за счёт расширяющихся сил, вызванных ростом клеток внутри колонии в присутствии поверхностно-активных веществ, которые уменьшают трение между клетками и минеральной поверхностью. Разные виды таксиса – способности двигаться к раздражителям или от них позволяет слизистым колониям сохраняться, размножаться, осваивать новые локусы обитания.
Обнаруженные диатомовые водоросли представлены одиночными особями с толстым кремнезёмистым панцирем и у особи рода Surirella – с выраженным слоем слизи вокруг него (рис. 4 / fig. 4). Помимо слизеобразования виды этого рода способны к вращательному движению.
Слизистые экзометаболиты, окружающие нитчатые цианобактерии и одиночные клетки диатомей, по литературным данным, содержат высокополимерные соединения, преимущественно полисахариды, уроновые кислоты, белки. Полисахариды являются основными местами хранения энергии, кислотные свойства слизи обеспечивают гелеобразование [3]. У цианобактерий полисахариды включают маннозу, рибозу, глюкозу, ксиллозу, глюкороновую килоту и другие соединения [11]. У диатомовых водорослей полисахариды представлены полимером глюкороновой кислоты – по-лиуронидом, 1,4-α-D-галактуроновой кислотой, 1,4-β-глюкуроновой кислотой, 1,4-β-D-маннуроновой кислотой [42]. Диатомеи имеют плотную слизь, цианобактерии – расплывчатую, хрящеватую. У многих представителей Nostocales она легко отделяется, у других, в частности у видов родов Calothrix, Lyngbya, Phor-midium, Tolypothrix, Myxosarcina и др., сохраняются вокруг организмов практически пожизненно [11].
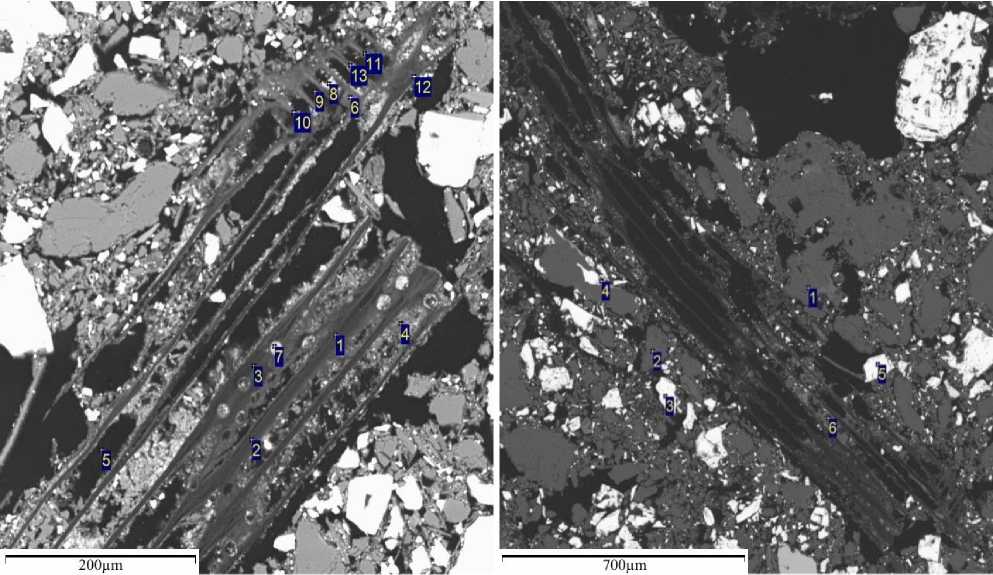
Рис. 1. Колонии нитевидных цианобактерий Fig. 1. Colonies of filamentous cyanobacteria

Рис. 2. Schizothrix sp. (а), увеличенные клетки трихомов (б, в), расположение точек определения химических элементов (д) Fig. 2. Schizothrix sp. (a), enlarged cells of trichome (б, в), location of the chemical element detection points (д)
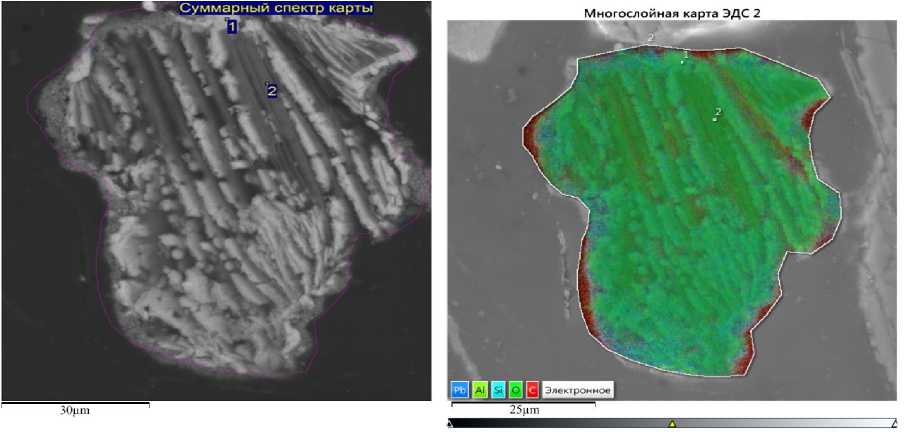
Рис. 3. Вид слизистой частично фоссилизованной колонии цианобактерии и её спектральное изображение Fig. 3. View of a partially fossilized cyanobacterium colony and its spectral image
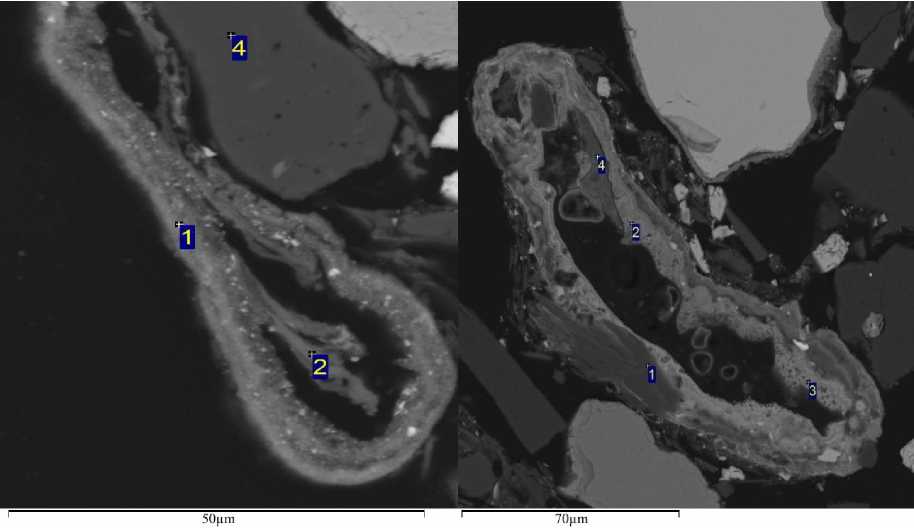
Рис. 4. Диатомеи: а – Surirella spiralis со слизью, б – Eunotia sp.
Fig. 4. Diatoms: а – Surirella spiralis with slime, б – Eunotia sp.
Большинство экзополисахаридов цианобактерий имеет анионную природу из-за присутствия уроновых кислот и/или других заряженных групп, таких как пирувил или сульфат [32]. В этой связи, положительно заряженные ионы некоторых металлов и металлоидов прочно закрепляются внутри отрицательно заряженных слизей. В дальнейшем их судьба может быть разной.
Химический анализ слизей жизнедеятельных микроорганизмов в пробах ТГ показал, что они содержат широкий набор элементов. У цианобактерии рода Schizothrix (согласно точкам определения), представленных выше (рис. 2д / fig. 2д), в слизи преобладает большинство металлов по сравнению с трихомами (табл. 1 / tabl. 1). В слизи обнаружены наибольшие значения Са, а также ионы Ba и Al. Можно предположить, что внеклеточные аморфные карбонаты кальция депо- нируются в слизях и цитозоле цианобактерий в качестве хранилища неорганического C и/или буфера pH. Такая стратегия выживания обеспечивает им развитие в окружении сульфидов и сульфатов, обусловливающих закисление среды обитания. Помимо этого, транзит Al и Ba в слизистый чехол сопровождается ограничением их подвижности, поскольку в цитозоле особей эти элементы не зарегистрированы.
Внеклеточное осаждение Pb, Al, Fe, P, S зафиксировано в слизистой части колонии другой цианобактерии (рис. 3 / fig. 3), где свинец преобладает (табл. 2 / tabl. 2). При этом в цитозоле присутствуют Si, Al, K и Ba. Такая иммобилизация металлов, в том числе токсичных, свидетельствует о высокой биоминерализации особи, что хорошо заметно на спектральном изображении (рис. 5 / fig. 5). Мы предполагаем, что в таком случае колония подвергается частичной фоссилизации.
Таблица 1
Химический состав цианобактерии Schizothrix sp.
по данным микрозондового анализа, мас. %*, точки (1-7) определения элементов в колонии
Table 1
Chemical composition of cyanobacteria Schizothrix sp.
according to microprobe analysis data, wt. %*, points (1-7) of determination of elements in the colony
|
№ точки определения // No. of the point of definition |
Показатель // Indicator |
||||||||
|
Al |
Si |
P |
S |
Cl |
K |
Ca |
Fe |
Ba |
|
|
1 |
0,28 |
1,12 |
0,53 |
0,11 |
4,91 |
||||
|
2 |
0,15 |
0,45 |
0,41 |
1,85 |
0,33 |
0,1 |
0,18 |
7,16 |
1,2 |
|
3 |
0,21 |
0,49 |
2,17 |
0,18 |
0,13 |
0,27 |
9,51 |
0, 77 |
|
|
4 |
0,91 |
3,79 |
0,08 |
0,24 |
14,51 |
0,22 |
|||
|
5 |
0,21 |
0,17 |
0,91 |
0,17 |
0,18 |
2,23 |
|||
|
6 |
0,15 |
0,21 |
0,71 |
1,15 |
|||||
|
7 |
0,09 |
0,78 |
3,62 |
0,07 |
0,26 |
12,87 |
|||
* Примечание : жирным шрифтом выделены значения накопления элементов в слизи.
* Note : the values of the accumulation of elements in the mucus are highlighted in bold.
Таблица 2
Химический состав частично фоссилизованной колонии цианобактерии по данным спектрального анализа, мас. %*, точки (1-2) определения элементов
В составе слизи диатомеи Surirella spiralis обнаружено присутствие жизненно необходимых и физиологически не значимых элементов (табл. 3 / tabl. 3). Внутри клетки диатомеи присутствует Fe, Zn, Pb, Si, Al, Ba. Можно предположить, что в кислой среде обитания, как известно, неблагоприятной для диатомей, их жизнедеятельность усугубляется высоким содержанием токсичных металлов. Аморфный кремнезём панциря и слизь слабо защищают клетки от транзита в них металлов.
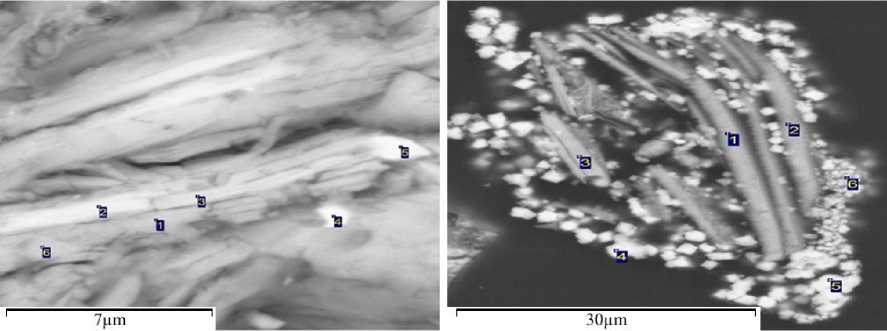
Следует также сказать, что развитие цианобактерий и диатомей происходит в окружении кристаллического барита (рис. 6 а-в / fig. 6 а-в). Кристаллы различаются по составу химических элементов. В одних случаях преобладает Ba на фоне убывания доли Si, S, Fe (рис. 6а / fig. 6а), в других – Ba лидирует на фоне снижения доли S, Ca, Fe (рис. 6в / fig. 6в). При этом содержание Ba в кристаллах превышает таковое в окружающей минеральной массе в десятки раз. Микробиологический анализ проб ТГ, образованных отходами выщелачивания сульфомышьяковистых полиметаллических руд в аридной зоне на горной территории Республики Тыва, показал, что в них развиваются цианобактерии и диатомеи, продуцирующие обильную слизь (рис. 7 / fig. 7). При этом цианобактерии присутствуют в колониальной форме. Колонии образуют как одиночные особи, так и многоклеточные нитевидные. В горной местности на высоте расположения хранилищ ТГ около 1000
м над уровнем моря в условиях аридизации и отсутствия высших растений наблюдаются высокие концентрации ультрафиолета и дефицит влаги.
Продуцирование экзометаболитов обеспечивает микроорганизмам сохранность от высокой инсоляции и пересыхания среды обитания. Помимо слизи в защите колоний могут участвовать мелкодисперсные частицы железа и кремнезёма, взвешенные в поровых водах, а в будущем – по мере формирования биогенно слаборазвитых почв – гуминовые кислоты. Установлено, что в моделируемых, геохимических условиях взвешенные в воде осадки оксигидроксида железа с кремнием, поглощали до 70% бактерицидного ультрафиолетового излучения и смягчали ультрафиолетовый стресс у прокариот [8]. Помимо них цианобактерии могут проявлять механизмы противодействия повреждению ультрафиолетовым излучением, такие как восстановление ДНК, детоксицирующие ферменты, пигменты и солнцезащитные молекулы, поглощающие ультрафиолетовое излучение. Важную роль в защите клеток от радиации играет синтез УФ-поглощаю-щих/экранирующих соединений, таких как микоспо-риноподобные аминокислоты и скитомелин [40].
Анализ содержания химических элементов в обнаруженных жизнедеятельных колониях одноклеточных цианобактерий р. Gloeocapsa sp. и Microcoleus sp. показал, что в них присутствуют разнообразные металлы и металлоид As (табл. 4, 5 / tabl. 4, 5).
Table 2
Chemical composition of a clock-fossilized cyanobacterium colony according to spectral analysis, wt. %*, element identification points (1-2)
|
№ точки определения // No. of the point of definition |
Показатель // Indicator |
|||||||||
|
Mg |
Al |
Si |
P |
S |
K |
Ca |
Fe |
Ba |
Pb |
|
|
1 |
9,76 |
0,49 |
7,27 |
2,32 |
0,25 |
0,18 |
4,64 |
28,69 |
||
|
2 |
0,72 |
16,33 |
20,4 |
0,25 |
6,56 |
0,97 |
2,09 |
|||
* Примечание : жирным шрифтом выделены значения накопления элементов в жизнедеятельных трихомах.
* Note : the values of accumulation of elements in vital trichomes are highlighted in bold .
Таблица 3
Химический состав слизи диатомеи Surirella spiralis.
по данным микрозондового анализа, мас. %*, точки (1-2) определения элементов
Table 3
The chemical composition of the mucus of Surirella spiralis . according to microprobe analysis data, wt. %*, element identification points (1-2)
|
№ точки определения // No. of the point of definition |
Показатель // Indicator |
|||||||||
|
Na |
Al |
Si |
S |
Ca |
Cr |
Fe |
Zn |
Ba |
Pb |
|
|
1 |
1,86 |
5,14 |
19,44 |
6,27 |
2,75 |
4,22 |
||||
|
2 |
1,46 |
0,58 |
8,53 |
1,45 |
1,67 |
0,48 |
13,31 |
6,92 |
5,57 |
|
* Примечание : жирным шрифтом выделены значения накопления элементов в слизи
* Note : The values of the accumulation of elements in the mucus are highlighted in bold
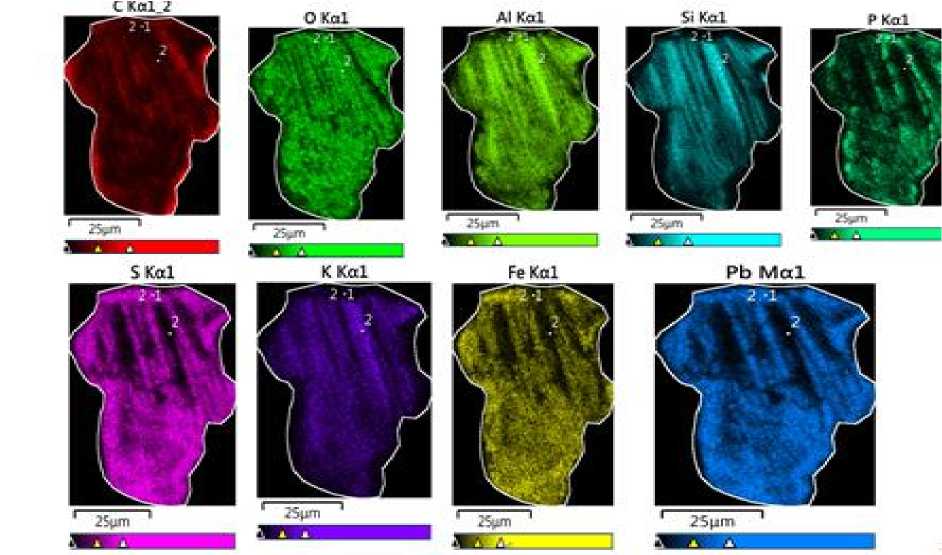
Рис. 5. Спектральное изображение химического состава колонии
Fig. 5. Spectral image of the chemical composition of the colony

Рис. 6. Кристаллические варианты барита (Ново-Урское месторождение-НУМ); точки (1-2) определения элементов в объекте
Fig. 6. Crystalline variants of barite (Novo-Urskoye deposit-NUM);
points (1-2) of determination of elements in the object
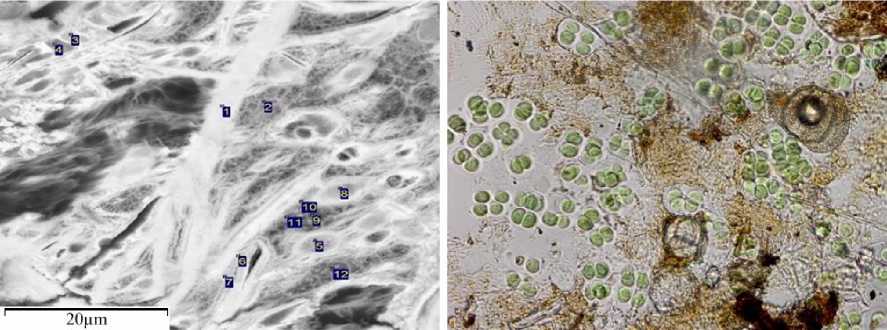
Gloeocapsa sp.
Microcoleus sp.

Pleurosigma sp. Synedra sp.

Synedra sp.
Рис. 7. Слизистые цианобактерии диатомеи Fig. 7. Mucous cyanobacteria and diatoms
Таблица 4
Химический состав Gloeocapsa sp.
по данным микрозондового анализа, мас. %*, точки (1-12) определения элементов
При этом в живых клетках колонии р. Gloeocapsa sp. его количество превышает таковое в слизи (табл. 4 / tabl. 4). Не исключено, что его цианобактерии используют для осуществления аноксигенного фотосинтеза. Такое явление было обнаружено, в высокогорном озере Моно на востоке американского штата Калифорния [35]. Оно было вызвано высокими концентрациями сульфидов и солей мышьяковистой кислоты. Существует мнение, что именно с участием таких доноров электронов мог начаться процесс аноксигенного фотосинтеза на Земле, а оксигенный фотосинтез, который снабжает другие формы жизни способствует азот-фиксации, которая обеспечивается присутствием Ca и Mg. Присутствие железа в колониях, вероятно, способствует защите от действия ультрафиолета.
Экспериментально доказано, что увеличение времени воздействия низких доз As5 и As3 способствует адаптации современных цианопрокариот и позитивно отражается на восстановлении роста и биохимических параметрах [39].
Диатомовые водоросли, представленные в ТГ аридной зоны р. Pleurosigma sp. и р. Synedra sp . (рис. 7 / fig. 7) относятся к подвижным формам, чему способствует слизь. В ней, как внутри клеток наблюдалась иммобилизация широкого набора химических элементов (табл. 6, 7 / tabl. 6, 7). Но As отсутствовал.
Не исключено, что эти организмы проявили стратегию защиты от мышьяка, которая включают усиление регуляции транспортеров, связанных с выведением вредных соединений из клетки, и белков, участвующих в синтезе глутатиона для хелатирования внутриклеточного мышьяка, что установлено ранее в ходе транскриптомного и биохимического анализа [43].
Анализ минералогического состава в среде обитания цианобактерий и диатомей показал, что в ней присутствуют кристаллические формы мышьяковистых минералов (рис. 8 а, б / fig. 8 а, б). В их составе преобладает As: 25,9-26,9 мас., % (рис. 8а / fig. 8a), и 26,726,8 мас., %. (рис. 8б / fig. 8б).
Сопутствующие элементы различаются по составу и содержанию. Ниже приводятся два варианта кристаллов: в одном присутствуют (в порядке убывания) As, Mg, Ni, Co, Cu, Ca, Mn, Fe, в другом – As, Mg, Co, Ni, Cu, Al, Si, Ca, Mn, Fe.
Не исключено, что зарождение минералов обусловлено испарительным эффектом и микробиологическими процессами. Биогеохимия мышьяка сложна и включает в себя различные процессы адсорбции и десорбции. В окислительных условиях мышьяк может высвобождаться из пирита или оксидов железа, особенно при повышенном уровне pH, что имеет место в ТГ аридной зоны. Его высокая иммобилизация происходит в слизистых колониях. Учитывая, что продуцирование слизистых экзометаоболитов – процесс чрезвычайно энергозатратный, на него расходуется до 70% энергии всего запаса фотосинтеза [30], можно предположить, что важную роль в формировании АТФ играет как традиционный фотосинтез (оксигенный), так и аноксигенный, с участием арсенитов, и сульфидов, поставщиков электронов. Не исключено также, что неорганический мышьяк, присутствующий в слизях, частично подвержен хелатированию и мета-болизирован в процессе метилирования в цитозоле с образованием органических соединений мышьяка, что обеспечивает его присутствие практически в нетоксичной для биоты форме [29].
Table 4
Chemical composition of Gloeocapsa sp.
according to microprobe analysis data, wt. %*, points (1-12) of determination of elements
|
№ точки определения // No. of the point of definition |
Показатель // Indicator |
||||||||||
|
Mg |
Si |
Cl |
Ca |
Mn |
Fe |
Co |
Ni |
Cu |
Zn |
As |
|
|
1 |
1,81 |
3,8 |
0,31 |
7,14 |
6,35 |
5.7 |
0,67 |
27,8 |
|||
|
2 |
1,19 |
0,11 |
2,56 |
0,17 |
3,97 |
5,9 |
3,15 |
0,38 |
18,07 |
||
|
3 |
0,9 |
0,16 |
6,29 |
0,15 |
5,59 |
6,7 |
4,08 |
0,76 |
23,43 |
||
|
4 |
0,98 |
4,82 |
0,17 |
4,86 |
5,78 |
3,49 |
0,71 |
19,17 |
|||
|
5 |
1,43 |
3,27 |
0,17 |
5,7 |
6,05 |
4,45 |
0,53 |
21,69 |
|||
|
6 |
1,6 |
3,66 |
0,28 |
6,28 |
7,15 |
5,36 |
0,66 |
26,28 |
|||
|
7 |
1,23 |
3,94 |
0,2 |
6,08 |
7,64 |
4,96 |
0,57 |
0,36 |
25,45 |
||
|
8 |
1,49 |
3,25 |
5.4 |
5,35 |
4,51 |
0,67 |
21,36 |
||||
|
9 |
0,55 |
0,13 |
2,61 |
3,86 |
4,8 |
2,88 |
0,34 |
13,67 |
|||
|
10 |
1,69 |
3,89 |
0,29 |
6,11 |
6,76 |
4,85 |
0,64 |
24,94 |
|||
|
11 |
1,09 |
0,2 |
0,1 |
3,24 |
6,12 |
6,12 |
3,92 |
0,46 |
19,29 |
||
|
12 |
0,73 |
0,13 |
2,63 |
0,17 |
4,98 |
4,98 |
2,81 |
0,39 |
14,26 |
||
* Примечание: жирным шрифтом выделены значения накопления As слизью цианобактерией.
* Note: the values of As accumulation by mucous cyanobacteria are highlighted in bold .
Таблица 5
Химический состав Microcoleus sp.
по данным микрозондового анализа, мас. %*, точки (1–6) определения элементов
Table 5
Chemical composition of Microcoleus according to microprobe analysis data, wt. %*, points (1-6) of determination of elements
|
№ точки определения // No. of the point of definition |
Показатель // Indicator |
||||||||||||||
|
Mg |
Al |
Si |
S |
Ca |
Cr |
Mn |
Fe |
Co |
Ni |
Cu |
As |
Sr |
Ba |
Bi |
|
|
1 |
15,55 |
0,78 |
0,15 |
0,82 |
0,22 |
0,93 |
2,92 |
1,59 |
1,7 |
28,83 |
|||||
|
2 |
9,93 |
0,41 |
0,71 |
0,32 |
2,19 |
4,63 |
9,68 |
0,83 |
25,48 |
||||||
|
3 |
13,36 |
0,9 |
0,14 |
1,51 |
0,16 |
1,73 |
3,37 |
4,78 |
1,49 |
28,6 |
|||||
|
4 |
12,78 |
0,57 |
1,44 |
1,03 |
0,26 |
0,39 |
2,77 |
2,16 |
0,76 |
1,52 |
16,86 |
||||
|
5 |
6,38 |
7,13 |
2 |
0,23 |
1,62 |
2,25 |
0,74 |
0,84 |
0,61 |
20,53 |
2,26 |
0,61 |
4,51 |
||
|
6 |
16,06 |
0,63 |
0,6 |
0,23 |
0,73 |
3,08 |
1,45 |
1,52 |
29,68 |
||||||
* Примечание : жирным шрифтом выделены значения накопления As цианобактерией.
* Note: the values of As accumulation by cyanobacteria are highlighted in bold .
Таблица 6
Химический состав Pleurosigma sp.
по данным микрозондового анализа, мас. %*, точки (1–6) определения элементов
Table 6
Chemical composition of Pleurosigma sp. according to microprobe analysis data, wt. %*, points (1-6) of determination of elements
|
№ точки определения // No. of the point of definition |
Показатель // Indicator |
||||||||||||
|
Mg |
Al |
Si |
S |
Cl |
K |
Ca |
Ti |
Mn |
Fe |
Co |
Ni |
Cu |
|
|
1 |
4,36 |
7,41 |
15,81 |
2,8 |
0,99 |
0,57 |
0,35 |
16,6 |
|||||
|
2 |
4,54 |
7,72 |
16,34 |
3,19 |
0,79 |
0,58 |
0,35 |
17,26 |
0,51 |
0,22 |
|||
|
3 |
4,37 |
8,17 |
17,08 |
4,52 |
0,56 |
0,51 |
0,37 |
17,13 |
0,29 |
0,25 |
|||
|
4 |
2,51 |
4,86 |
10,66 |
0,12 |
2,63 |
0,64 |
0,41 |
0,28 |
12,54 |
||||
|
5 |
4,16 |
7,26 |
15,5 |
3,57 |
0,61 |
0,52 |
0,37 |
15,93 |
0,43 |
0,2 |
0,23 |
||
|
6 |
1,18 |
10,76 |
19,47 |
0,1 |
0,12 |
0,99 |
0,64 |
3,14 |
0,45 |
0,28 |
|||
* Примечание: жирным шрифтом выделены значения накопления элементов в слизи.
* Note: The values of the accumulation of elements in the mucus are highlighted in bold.
Таблица 7
Химический состав Synedra sp.
по данным микрозондового анализа, мас. %*, точки (1–9) определения элементов
Table 7
Chemical composition of Synedra sp.
according to microprobe analysis data, wt. %*, points (1-9) of determination of elements
|
№ точки определения // No. of the point of definition |
Показатель // Indicator |
|||||||||
|
Na |
Mg |
Al |
Si |
Cl |
K |
Ca |
Ti |
Fe |
Ba |
|
|
1 |
0,46 |
0,83 |
3,12 |
5,75 |
0,43 |
0,73 |
3,94 |
1,52 |
||
|
2 |
0,55 |
1,31 |
10,97 |
17,67 |
0,18 |
5,53 |
0,97 |
0,18 |
2,8 |
|
|
3 |
0,31 |
1,49 |
13,93 |
22,15 |
7,78 |
0,39 |
3,19 |
|||
|
4 |
0,34 |
1,57 |
14,96 |
24,34 |
8,13 |
3,22 |
||||
|
5 |
0,22 |
1,36 |
12,1 |
19,49 |
0,12 |
6,51 |
0,35 |
0,17 |
2,7 |
|
|
6 |
0,33 |
1,44 |
12,39 |
20,81 |
7,14 |
0,9 |
0,15 |
3 |
||
|
7 |
0,27 |
1,77 |
14,22 |
23,98 |
7,67 |
1,15 |
0,14 |
3,33 |
||
|
8 |
0,61 |
6,96 |
5,42 |
22,55 |
1,4 |
7,5 |
0,26 |
10,13 |
||
|
9 |
0,17 |
1,23 |
10,52 |
16,92 |
0,15 |
5,37 |
2,39 |
2,39 |
0,27 |
|
*Примечание : жирным шрифтом выделены значения накопления элементов в слизи
*Note : The values of the accumulation of elements in the mucus are highlighted in bold

Рис. 8. Кристаллические мышьяковистые минералы (A, Б)
Fig. 8. Crystalline arsenic minerals (A, Б)
Существует мнение, что такие свойства цианобактерий можно использовать в эффективном связывании токсичных элементов, в том числе мышьяка, для ускорения очистки экосистем [34 ]. В настоящее время для биоремедиации от мышьяка предлагаются суль-фатредуцирующие бактерии [7]. Не исключено применение с этой целью штаммов цианобактерий, присутствующих в окружении мышьяка в экстремальных условиях Южной Сибири.
Заключение
Методом сканирующего электронного микроскопирования были изучены особенности биогенности ТГ, представляющие собой отходы переработки сульфидных золотосодержащих руд (гумидная зона Западной Сибири, климат резко континетальный) и сульфо-арсенидных полиметаллических руд (холодная аридная зона горной территории, юг Сибири). Установлено, что в таких крайне экстремальных местообитаниях, каковыми являются токсичные хранилища ТГ, через десятилетия их пребывания на дневной поверхности, в оксигенном слое поселяются диатомовые водоросли и цианобактерии. Они обнаружены нами в жизнедеятельном состоянии, что обеспечивается формированием колоний с присутствием в большинстве случаев слизистых экзометаболитов. Химический состав слизей и талломов, определённый с помощью микрозон-дового (спектрального) анализа элементов, включает тяжёлые металлы и неметаллы, в том числе не являющиеся физиологически значимыми, относящихся к группе биотоксичных. Установлено, что в техногенных грунтах, представленных сульфидными отходами цианирования руд, диатомовые водоросли аккумулируют барий внутри клеток, цианобактерии в слизи. В сульфо-мышьяковистом субстрате цианобактерии иммобилизируют As внеклеточно и внутриклеточно. Предполагается, что мышьяк используется цианобактериями в аноксигенном фотосинтезе для получения дополнительной энергии и азота путём фиксации из атмосферы. Диатомовые водоросли металлоид не содержат что, возможно, обусловлено их механизмами биохимической защиты. Дефицит трофического резерва диатомеи наряду с цианобактериями, способны компенсировать гетеротрофно [36], возможно, используя собственные экзополисахариды.
Установлено, что цианобактерии и диатомеи сохраняют жизнедеятельность в окружении не только растворённых химических элементов, но и в окружении кристаллических баритовых и мышьяковистых минералов. Не исключено, что причинами зарождения кристаллов являются испарительный эффект и микробиологические процессы.
Наши сведения о жизнедеятельности этих организмов в ТГ расширяют знания о крайне экстремальных местообитаниях, пригодных для их жизни. Ранее сообщалось, что диатомеи и цианобактерии обнаружены во льдах Арктики [18, 19] наряду с бактериями и вирусами [21], цианобактерии – в песках пустыни Атакама в Чили – одном из самых засушливых мест на Земле [33], где они используют не жидкую воду, а кристаллическую – из структуры минерала ангидрида. В жизнедеятельном состоянии цианобактерии и диатомеи обнаружены и в других малопригодных для жизни субстратах: песчаных дюнах, на поверхности застывшей вулканической лавы, гейзеритов, солонцов и др.
Изложенные результаты о биогенности ТГ могут быть использованы для характеристики ранних стадий почвообразования в экстремальных местообитаниях техногенных ландшафтов, учтены при разработке экологически безопасных способов консервации грунтов, создании природоподобных технологий эффективной биоремедиации почв, при моделировании процессов образования минералов.
Сведения об авторском вкладе
В.С. Артамонова – постановка задачи исследования, формулировка идеи статьи, работа с иностранными источниками, написание и научное редактирование статьи.
А.Ш. Шавекина – отбор проб в полевых условиях, подготовка монолитов для сканирующего электронного микроскопирования, определение химических элементов с применением микрозондового анализа, компьютерная обработка снимков.
Contribution of the authors
V.S. Artamonova – formulation of the research task, formulation of the idea of the article, work with foreign sources, writing and scientific editing of the article.
A.Sh. Shavekina – sampling in the field, preparation of monoliths for scanning electron microscopy, determination of chemical elements using microprobe analysis, computer image processing.
