Темная полевка (Microtus agrestis L.) в коренных и антропогенных ландшафтах Восточной Фенноскандии
 в коренных и антропогенных ландшафтах Восточной Фенноскандии")
Автор: Ивантер Эрнест Викторович, Курхинен Юрий Павлович, Соколов Артем Владимирович
Журнал: Ученые записки Петрозаводского государственного университета @uchzap-petrsu
Рубрика: Биология
Статья в выпуске: 8 (129) т.1, 2012 года.
Бесплатный доступ
Рассматриваются популяционно-экологические особенности темной полевки в условиях антропогенной трансформации таежных экосистем Восточной Фенноскандии, закономерности пространственной структуры популяции, изменений численности, взаимоотношения с видами-конкурентами.
Популяция, экосистема, динамика численности, биотопическое размещение
Короткий адрес: https://sciup.org/14750331
IDR: 14750331 | УДК: 599.32
Dark field mouse (Microtus agrestis L.) in original anthropogenic landscapes of Eastern Scandinavia
Ecological and population features of the dark field mouse (Microtus agrestis L.) in conditions of anthropogenic transformation of taiga ecosystem in Eastern Scandinavia are studied. Consistencies for spatial population structure, population size, and interrelationships with other competing species are analyzed.
Текст научной статьи Темная полевка (Microtus agrestis L.) в коренных и антропогенных ландшафтах Восточной Фенноскандии
В Восточной Фенноскандии (к которой относят Финляндию, Кольский полуостров, Карелию и Карельский перешеек Ленинградской области) темная полевка распространена повсеместно и по численности и доминированию в населении мелких млекопитающих занимает твердое третье место (после обыкновенной бурозубки и рыжей полевки). Вместе с тем этот вид заселяет рассматриваемую территории неравномерно. Наибольшая его численность отмечена в южных районах, особенно в местах, сильно затронутых хозяйственной деятельностью человека, а на севере и востоке ареала он встречается спорадически и обычно в небольшом количестве (табл. 1 и 2).
В целом по региону численность темной полевки колебалась по годам в пределах 0,01–2,0 (см. рисунок) и составила в среднем за все годы учетов (1958–2012) 0,3 экз. на 100 ловушко-суток (7,6 % в уловах ловушками). Канавочные учеты за этот же период дали средний показатель 0,38 экз. на 10 канавко-суток (индекс доминирования – 3,2 %) с колебаниями от 0 до 1,6 (0–12,7 %). Сопоставление этих данных с соответствующими показателями учетов численности вида в других частях ареала [1], [3], [6], [7], [9], [14] позволяет говорить о сравнительно невысокой плотности его населения в Восточной Фенноскандии, особенно в северной ее половине. Общая невысокая численность темной полевки на этой территории объясняется сужением экологического ареала вида вблизи северной границы распространения и, в частности, более тесной его связью с антро
погенным ландшафтом, который занимает здесь весьма ограниченные площади.
Таблица 1
Среднегодовые показатели численности темной полевки в Карелии по данным учетов ловушками (июль–октябрь, 1958– 2011 годы)
|
Районы и годы учетов |
Число добытых зверьков |
||
|
Абс. |
На 100 ловушко-суток |
% от улова мелких млекопитающих |
|
|
Лоухский, Калевальский, Кемский (1959, 1961–1962, 1973, 1994–1997) |
66 |
0,21 |
10,1 |
|
Сегежский, Суоярвский (1958, 1960) |
29 |
0,50 |
8,8 |
|
Медвежьегорский (1958, 1962, 1964–1965) |
45 |
0,08 |
6,0 |
|
Кондопожский (1959, 1966–1972) |
107 |
0,24 |
7,4 |
|
Пудожский (1961–1963, 1966) |
9 |
0,03 |
1,3 |
|
Питкярантский, Сортавальский (1966–1967, 1969–2011) |
288 |
0,13 |
3,2 |
|
Лахденпохский (1958–1959, 1966) |
28 |
0,10 |
5,6 |
|
Прионежский (1963, 1966, 1970–1973, 1977–1979) |
215 |
0,36 |
6,4 |
|
Пряжинский (культурный ландшафт, 1966) |
56 |
1,80 |
25,9 |
|
Пряжинский (лесные биотопы, 2002–2011) |
8 |
0,03 |
0,7 |
Таблица 2
Результаты учета темной полевки в Карелии ловчими канавками (июль –октябрь, 1959–2011 годы)
|
Районы и годы учета |
Число добытых зверьков |
||
|
Абс. |
На 10 канавко-суток |
% от улова мелких млекопитающих |
|
|
Кондопожский (1959–1960) |
112 |
0,89 |
6,1 |
|
Медвежьегорский (1962–1964, 1966–1968) |
16 |
1,27 |
17,2 |
|
Питкярантский (1966–1967, 1969–2011) |
75 |
0,12 |
1,08 |
|
Пудожский (1961–1963) |
4 |
0,06 |
0,9 |
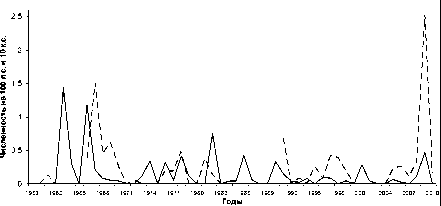
-----2
Динамика численности темной полевки в Карелии по данным учетов ловушко-линиями (1) и канавками (2)
Особенности биотопической приуроченности темной полевки в условиях Восточной Фен-носкандии отражают данные табл. 3. В исследованном регионе она наиболее многочисленна в сельскохозяйственных угодьях и по окраинам населенных пунктов, а также на зарастающих лесосеках с разбросанными порубочными остатками, неубранными бревнами и гнилыми пнями. Довольно часто темная полевка встречается на сырых лугах, по опушкам травяных хвойных и лиственных лесов, по краям болот, в лиственном мелколесье и т. п. В пределах этих местообитаний она выбирает влажные захламленные участки с зарослями высоких травянистых растений и кустарников. В южных частях региона темная полевка распространена более равномерно и населяет многие лесные стации, хотя и с меньшей плотностью, чем луговые и полевые угодья. На севере же она более стенотопна и встречается главным образом на лугах, лесных полянах, вырубках, по опушкам лиственных лесов, обочинам дорог и на полях, избегая закрытых стаций, и особенно монотонных хвойных насаждений. Характер стациального размещения зависит также от сезона и численности зверьков. В годы наиболее высоких пиков (1951, 1957, 1963, 1966, 1967, 1982, 1990, 2010) население полевок широко растекается и они встречаются почти во всех биотопах. В большей степени это характерно для молодых расселяющихся зверьков, тогда как зимовавшие более консервативны и придерживаются лишь наиболее благоприятных мест обитания. Совершенно иная картина наблюдается при стойких депрессиях численности (1953–1960, 1970–1972, 1979–1981, 1991–1996, 2000–2008). В этих случаях популяция состоит из отдельных разрозненных поселений, приуроченных к «стациям переживания», которыми в наших условиях служат поросшие кустарником и высоким разнотравьем и сильно захламленные каменные гряды, межи и прочие «неудобные земли» на полях и сенокосах. В небольшом количестве они также сохраняются на посевах трав, опушках, лесных полянах, молодых вырубках и в понижениях, где есть заросли кустарников, высокий травостой и ягодники.
Таблица 3
Биотопическое распределение темной полевки
|
Биотоп |
Весна |
Лето |
Осень |
По всем сезонам |
|
|
Показатель численности |
Индекс верности биотопу |
||||
|
Учеты ловушко |
-линиями (экз. на 100 ловушко-суток) |
||||
|
Сосняки лишайниковые |
– |
– |
– |
– |
– |
|
Сосняки-зеленомошники |
– |
0,08 (3,3) |
0,08 (1,4) |
0,07 (2,7) |
-0,64 |
|
Ельники-зеленомошники |
0,02 (1,9) |
0,03 (1,1) |
0,05 (0,7) |
0,03 (1,0) |
-1,10 |
|
Смешанные хвойно-лиственные леса |
– |
0,04 (1,1) |
0,61 (6,7) |
0,09 (2,4) |
-0,45 |
|
Спелые лиственные леса |
0,01 (2,0) |
0,22 (5,8) |
0,09 (1,6) |
0,18 (5,1) |
+0,36 |
|
Лиственное мелколесье |
– |
0,21 (4,9) |
0,22 (3,5) |
0,20 (4,8) |
+0,55 |
|
Вырубки |
– |
0,24 (4,4) |
0,02 (0,3) |
0,20 (3,8) |
+0,55 |
|
Окультуренный ландшафт |
0,07 (5,6) |
0,59 (12,0) |
1,03 (13,3) |
0,52 (14,1) |
+3,45 |
|
Окраины болот |
– |
0,13 (10,0) |
– |
0,12 (11,0) |
-0,18 |
|
Опушки леса |
– |
0,20 (5,6) |
1,06 (9,6) |
0,66 (8,8) |
+4,72 |
|
Учеты канавками (экз. на 10 канавко-суток) |
|||||
|
Сосняки-зеленомошники |
– |
0,10 (0,9) |
0,06 (0,5) |
0,09 (0,8) |
-0,59 |
|
Ельники-зеленомошники |
– |
0,29 (2,1) |
– |
0,27 (2,0) |
+0,22 |
|
Смешанные хвойно-лиственные леса |
– |
0,26 (2,6) |
– |
0,24 (2,5) |
0 |
|
Спелые лиственные леса |
0,16 (2,0) |
0,18 (1,0) |
0,46 (3,7) |
0,20 (1,4) |
0 |
|
Лиственное мелколесье |
– |
0,25 (1,8) |
– |
0,24 (1,8) |
0 |
|
Вырубки |
– |
0,46 (3,7) |
5,7 (44,4) |
0,64 (5,4) |
+1,9 |
|
Окультуренный ландшафт |
1,25 (100,0) |
1,00 (7,3) |
3,17 (26,0) |
1,37 (11,4) |
+5,2 |
Сезонные изменения биотопического размещения темной полевки сводятся в основном к перераспределению численности. Весной и в начале лета плотность населения полевок минимальная и зверьки концентрируются в немногих лучших местообитаниях, чаще всего в пределах антропогенного ландшафта, по травяным опушкам и лесным пожням. Летом отдельные поселения постепенно сливаются и зверьки более или менее равномерно заселяют все подходящие биотопы. Осенью эта дисперсия особенно хорошо выражена, но к зиме полевки вновь концентрируются в немногочисленных поселениях и их распространение приобретает очаговый характер. В это время они часто отлавливаются в стогах, на усадьбах и в хозяйственных постройках, безраздельно господствуя в населении мелких млекопитающих антропогенного ландшафта. Таким образом, зимой, весной и в начале лета, а также во все сезоны в годы стойких депрессий численности для темных полевок Восточной Фенноскандии характерен мозаичный тип поселений, а в июле – октябре – диффузный, равномерный. И поскольку перестройка типа поселения носит здесь как сезонный, так и годовой циклический характер при сохранении сети постоянных элементарных поселений, свойственный данной популяции тип пространственной структуры можно по классификации В. Е. Флинта [16], [17] отнести к пульсирующему.
Особого внимания заслуживает вопрос о территориальных отношениях темной полевки с полевкой-экономкой. На сырых лугах, в мелколесье по берегам водоемов, на болотах и сырых участках антропогенного ландшафта их экологические ниши наиболее полно совпадают, что ведет к конкуренции. В наиболее яркой форме территориальный антагонизм, приводящий к четкой пространственной разобщенности этих двух видов, мы наблюдали в 1960–90-х годах в окрестностях полевых стационаров в Прионежском (агробаза Института биологии КарНЦ РАН) и Питкярантском (дер. Карку) районах Карелии. В более благоприятных, влажных, с густым и сочным травостоем местах здесь всегда количественно преобладала более крупная и сильная полевка-экономка, а в более сухих и менее кормных – темная полевка. Вместе с тем отчетливая территориальная конкуренция и вызванное ею количественное доминирование экономки были заметны в основном лишь в годы ее высокой численности, а в остальное время зверьки обоих видов вполне мирно уживались и одинаково часто отлавливались в одних и тех же местах. К аналогичному выводу о зависимости био-топического распределения темной полевки от численности экономки как более сильного конкурента приходит по наблюдениям в Северной Финляндии и Дж. Таст [21]. Так, по его данным, в годы высокой численности экономки темная полевка исчезает почти из всех антропогенных местообитаний и большинства торфяников, а во время очень высоких пиков – даже из светлых травянистых лесов. Между тем в годы низкой численности экономки темная полевка снова занимает эти места обитания и живет бок о бок с немногочисленными особями вида-конкурента. Подробное описание этих явлений, наблюдавшихся в других частях ареала данных видов, можно найти и в трудах известных классиков отечественной териологии [2], [5], [13], [18].
Согласно специальным исследованиям [10], [11], [12], отношение темной полевки к сплошным концентрированным рубкам леса в целом гораздо терпимее, чем других обитателей таежных экосистем. В отдельных биотопах сукцессионного ряда (вырубки, особенно злаковых типов) она составляет до 45 % (учеты давилками) – 76 % (учеты канавками, табл. 4). Анализ данных многолетних учетов показывает, что на вырубках происходит заметное увеличение численности темной полевки, создающей здесь в течение нескольких лет основной фон населения мышевидных грызунов. В последующие годы по мере восстановления древесного полога и угнетения нижних ярусов растительности обилие и индекс доминирования темной полевки постепенно сокращаются. Коэффициент корреляции между давностью рубки (от 1–2 до 25 лет) и индексом доминирования вида составил -0,95 ( p < 0,05), то есть изменения идут в противоположном, по сравнению с рыжей полевкой, направлении. Характер взаимоотношений темной и рыжей полевок, демонстрирующих совершенно разную реакцию на рубку леса, неоднократно обсуждался в литературе. При этом предполагалась возможность конкуренции между этими видами на ранних стадиях вторичной сукцессии лесных экосистем [19]. По материалам исследований в Финляндии [20], рыжая полевка может испытывать конкурентное давление со стороны темной полевки, особенно в годы высокой численности последней. Не исключая конкуренцию, определяющим фактором мы все же считаем резкое изменение экологических условий на вырубках, дающее временное преимущество темной полевке. В Карелии летом это типично зеленоядный вид, зимой в его рационе значительное место занимают кора и побеги древесных пород [7]. Увеличение фитомассы травянистых кормов и количества подроста на вырубках благоприятно сказывается на кормовой базе и, соответственно, на обилии темной полевки. Нами установлена достоверная корреляционная связь между фитомассой травяно-кустарничкового яруса растительности и численностью темной полевки (+0,89, p < 0,01). В дальнейшем восстановление лесной среды в процессе вторичной сукцессии растительности ведет к ухудшению экологических условий для темной полевки. В результате преимущество получает рыжая полевка, которая вновь занимает лидирующее положение среди населения лесных грызунов.
|
Таблица 4 |
|||||||||
|
Изменение численности и соотношения фоновых видов мышевидных грызунов |
|||||||||
|
в коренных и трансформированных |
рубками леса |
биотопах |
|||||||
|
Рыжая полевка |
Темная полевка |
Полевка-экономка |
|||||||
|
Биотоп |
|||||||||
|
I |
II |
III |
I |
II |
III |
I |
II |
II |
|
|
А. Учеты на ловушко-линиях: |
|||||||||
|
Сосняки зеленомошные, спелые |
2,8 |
60 |
94 |
0,14 |
2,5 |
5 |
0 |
0 |
0 |
|
Ельники зеленомошные, спелые |
2,8 |
62 |
98 |
0,02 |
0,4 |
0,7 |
0 |
0 |
0 |
|
Открытые вырубки |
1,9 |
22 |
37 |
2,3 |
26 |
45 |
0,8 |
8 |
16 |
|
Молодняки 6–20 лет |
2,9 |
39 |
66 |
10 |
14 |
23 |
0,4 |
5 |
9,1 |
|
21–40 лет |
2,0 |
72 |
100 |
0 |
0 |
0 |
0 |
0 |
0 |
|
Вторичные древостои 50–70 лет |
2,1 |
43 |
92 |
0,06 |
1 |
2,6 |
0,1 |
3 |
4,4 |
|
Семенные куртины |
2,3 |
58 |
96 |
0,1 |
2 |
4,2 |
0 |
0 |
0 |
|
Недорубы |
3,8 |
55 |
89 |
0,4 |
5 |
9,4 |
0 |
0 |
0 |
|
Экотоны (лес-вырубка) |
4,7 |
39 |
55 |
2,9 |
25 |
34 |
0,9 |
8 |
10,6 |
|
В. Учеты ловчими конусами |
|||||||||
|
Сосняки зеленомошные, спелые |
2,2 |
40 |
95 |
0,6 |
10 |
26 |
0 |
0 |
0 |
|
Ельники зеленомошные, спелые |
1,0 |
27 |
100 |
0 |
0 |
0 |
0 |
0 |
0 |
|
Открытые вырубки |
0,1 |
2 |
5 |
1,4 |
32 |
76 |
0,1 |
3 |
5,4 |
|
Молодняки 6–20 лет |
0,5 |
9 |
40,6 |
0,6 |
14 |
49 |
0 |
0 |
0 |
|
20–30 лет (смешанные) |
2,0 |
45 |
77 |
0,6 |
10 |
23 |
0 |
0 |
0 |
|
30–40 лет (хвойные) |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
Вторичные древостои 50–70 лет |
1,1 |
24 |
50 |
0 |
0 |
0 |
0,2 |
4 |
9,1 |
|
Семенные куртины |
1,6 |
23 |
59 |
0,8 |
14 |
29,6 |
0 |
0 |
0 |
|
Недорубы |
0,7 |
12 |
70 |
0,2 |
3 |
20 |
0 |
0 |
0 |
Примечание. I – численность (экз. на 100 ловушко-суток – А, на 10 конусо-суток – В); II – % от улова мелких млекопитающих; III – % от улова мышевидных грызунов.
Подснежные учеты зверьков в феврале на открытой злаковой вырубке и в сосняке-зеленомошнике показали, что темная полевка в период высокой численности предпочитает селиться на открытых вырубках не только летом, но и зимой (1,3 экз. на 100 ловушко-суток, 21 % в уловах мелких млекопитающих). Однако доминирование темной полевки в отловах грызунов характерно только для территории злаковых (луговиковых, вейниково-луговиковых) вырубок. Правда, они составляют значительную часть всех вырубок региона [4], [8], [15].
На вырубках иных типов темная полевка может не преобладать в уловах, хотя ее обилие и индекс доминирования здесь повышаются. Очевидно, только типом растительности на вырубках и можно объяснить некоторые закономерные географические различия численности и доминирования определенных видов мелких млекопитающих в разных регионах таежной зоны России. В одних регионах доминирует темная полевка (или другие представители рода Microtus), в других наблюдается только рост ее численности при сохранении доминирования за лесными (рыжими) полевками. Вполне естественно, что разные исследователи выбирали в качестве «контроля» различные типы леса, а в качестве «опыта» – разные типы вырубок. Не исключено, что в пределах разных регионов может наблюдаться широкий спектр различных реакций одного и того же вида животного на рубку леса, в зависимости от характера лесного биоценоза, типа вырубки, почв, рельефа и др., как это и установлено нами в Восточной Фенноскандии.
Темная полевка хорошо приспособлена к жизни на вырубках. В годы пиков численности она активно осваивает не только оптимальные для нее злаковые (вейниково-луговиковые) вырубки, но и заболоченные, осоково-сфагновые (в том числе зимой). По нашим наблюдениям, на злаковых вырубках зимой полевки устраивают подснежные гнезда в центральной части дернины злаков, постепенно выедают живые части растений, а отмершие стебли и листья используют как теплоизолятор. Летом зверьки устраивают норы в заросших травянистой растительностью отвалах почвы из-под гусениц трелевочных тракторов. Почва здесь рыхлая, она как бы приподнята над остальной поверхностью. В условиях повышенного поверхностного стока на открытых лесосеках такие норы практически не затопляются.
Численность темной полевки на вырубках испытывает не столь сильные колебания по годам, как в спелых сосняках (коэффициент вариации CV соответственно 107 и 188 %). Летне-осенняя численность этого вида на вырубках обычно в 5–10 раз превышает весеннюю, при этом особенно быстрый рост происходит в первой половине лета (в среднем за 11 лет наблюдений – девятикратное). Впрочем, бывают и исключения из этого правила. На одном из экспериментальных участков вырубки (злаковый тип, очистка – сбор порубочных остатков в валы и кучи, есть лесные культуры ели) было зафиксировано значительное, в 4 раза, сокращение численности темной полевки от весны к осени. Однако это скорее всего было связано с отмеченной здесь достаточно высокой численностью других видов полевок – экономки и рыжей. При этом последняя, в отличие от темной, не только не сократила численность к концу бесснежного сезона, а напротив, увеличила ее в 8 раз. В этих условиях темная полевка смогла достичь достаточно высокой численности только на краю вырубки – на границе с лесом, что для данного вида в целом нехарактерно. Это нельзя объяснить нечем иным, как вытеснением темной полевки с территории вырубки другими видами полевок – экономкой и рыжей, что косвенно подтверждает вероятность обострения их конкурентных отношений при высокой численности последних.
В сомкнутых древостоях по краю вырубок (хвойные недорубы, участки частоствольных молодняков) темная полевка встречается не постоянно и в небольшом количестве, в основном лишь в годы пиков численности и ближе к осени, в периоды массового расселения молодых. Антропогенная трансформация ландшафта в целом позитивно сказывается на состоянии ее популяций, ее численность здесь значительно выше, чем в крупных массивах незатронутых рубками хвойных лесов. В среднем за 11 лет исследования превышение достигло семикратной величины, а в слабо фрагментированных сосняках – даже десятикратной. Помимо прочего, это связано и с более благоприятной для успешного воспроизводства возрастной структурой обитающего на вырубках поголовья: в период массового размножения зимовавшие и половозрелые прибылые зверьки составляли здесь до половины всего населения, тогда как в сомкнутых хвойных насаждениях их либо не было вовсе, либо их количество не превышало 10 %, остальных представляли лишь молодые расселяющиеся животные. Таким образом, в условиях периферии ареала вырубки служат для популяции темных полевок важнейшей стацией размножения. Недаром именно здесь, на вырубках, было отловлено подавляющее большинство (84 %) беременных самок этого вида.
Итак, вызванная массовыми рубками фрагментация хвойных лесов и формирование мозаичного антропогенного ландшафта – в целом позитивный для темной полевки процесс. Особенно благоприятно сказываются на существовании этого вида формирующиеся на хорошо очищенных лесосеках злаковые вырубки, составляющие наряду с сельхозугодьями основные места его обитания в Восточной Фенноскандии. В этих условиях темная полевка проявляет себя как типичный обитатель открытых пространств, хорошо приспособленный к освоению и заселению злаковых вырубок. Выполняя важнейшую функцию первопоселенца, необходимого первичного звена в осуществлении процесса лесовосстановления, этот вид при благоприятных условиях (очистка лесосек, отсутствие конкурентов, мощный травяно-кустарничковый ярус) может достигать очень высокой численности (в отдельные годы – до 18–20 экз. на 100 ловушко-суток) и тем самым подготавливает условия для дальнейшего заселения вырубок другими видами животных, и прежде всего хищниками-миофагами – лесной куницей, горностаем, лисицей, лесным хорьком и др. В дальнейшем, по мере восстановления лесной растительности и формирования исконной таежной фауны, происходит снижение как численности, так и индекса доминирования темной полевки и на смену ей приходят новые, более характерные обитатели спелого леса: рыжие полевки, землеройки-бурозубки, а также целый ряд более крупных охотничьих видов, таких как летяга, белка, лесная куница, заяц-беляк, рысь, барсук, медведь, лось и т. д.
* Работа выполнена при поддержке Программы стратегического развития (ПСР) ПетрГУ в рамках реализации комплекса мероприятий по развитию научно-исследовательской деятельности на 2012–2016 гг.
Список литературы Темная полевка (Microtus agrestis L.) в коренных и антропогенных ландшафтах Восточной Фенноскандии
- Ануфриев В. М. Темная (пашенная) полевка//Фауна европейского Северо-Востока России. Млекопитающие. Т. II, Ч. 1. СПб.: Наука, 1994. С. 254-263.
- Башенина Н. В. Материалы к экологии мелких млекопитающих зоны европейской тайги//Ученые записки Пермского педагогического института. 1968. Т. 52. С. 3-44.
- Воронов Г. А. География мелких млекопитающих южной тайги Приуралья, Средней Сибири и Дальнего Востока. Пермь: Изд-во Перм. ун-та, 1993. 223 с.
- Воронова В. С. К вопросу о классификации растительности вырубок Карелии//Возобновление леса на вырубках и выращивание сеянцев в питомниках. Петрозаводск, 1964. С. 23-32.
- Грибова З. А. К экологии пашенной полевки (Microtus agrestis L.)//Труды ВНИИОЗ. Вып. 18. М., 1959.
- Жигарев И. А. Мелкие млекопитающие рекреационных лесов Подмосковья. М.: Прометей, 2004. 232 с.
- Ивантер Э. В. Популяционная экология мелких млекопитающих таежного Северо-Запада СССР. Л.: Наука, 1975. 244 с.
- Крышень А. М., Гнатюк Е. П. Разнообразие растительных сообществ вырубок Карелии//Антропогенная трансформация таежных экосистем Европы. Петрозаводск: Карельский научный центр РАН, 2004. С. 178-172.
- Куприянова И. Ф. Биотопические группировки мелких млекопитающих и динамика их численных взаимоотношений на юге Архангельской области//Фауна и экология позвоночных животных. М., 1978. С. 114-130.
- Ку рхинен Ю. П. Воздействие сплошных концентрированных рубок на кормовые ресурсы и численность растительноядных млекопитающих Карелии//Влияние хозяйственного освоения лесных территорий Европейского Севера на население животных. М., 1987. С. 18-31.
- Курхинен Ю. П. Млекопитающие и тетеревиные птицы Восточной Фенноскандии в условиях антропогенной трансформации таежных экосистем: Автореф. дис.. д-ра биол. наук. Петрозаводск, 2001. 38 с.
- Курхинен Ю. П., Данилов П. И., Ивантер Э. В. Млекопитающие Восточной Фенноскандии в условиях антропогенной трансформации таежных экосистем. М.: Наука, 2006. 206 с.
- Наумов Н. П. Очерки сравнительной экологии мышевидных грызунов. М.; Л., 1948. 204 с.
- Попов И. Ю. Структура и динамика населения мелких млекопитающих в связи с сукцессиями растительности в Европейской южной тайге: Автореф. дис.. канд. биол. наук. М., 1998.
- Ронконен Н. И. Вырубки и естественное возобновление на них//Лесовосстановление в Карельской АССР и Мурманской области. Петрозаводск, 1975. С. 36-65.
- Флинт В. Е. Типы пространственной структуры популяций у мелких млекопитающих//Популяционная структура вида у млекопитающих. М.: Изд-во МГУ, 1970. С. 161-163.
- Флинт В. Е. Пространственная структура популяций мелких млекопитающих. М.: Наука, 1977. 182 с.
- Формозов А. Н. Мелкие грызуны и насекомоядные Шарьинского района Костромской области в период 1930-1940 гг.//Материалы по грызунам. М.: Изд-во МОИП, 1948. Вып. 3. С. 3-110.
- Henttonen H., Kaikusalo A., Tast J., Viitala J. Interspecific competition between smalol rodents in Subarctic and boreal ecosystems//Oicos. 1977. Vol. 29. P. 581-590.
- Larsson T. Small rodent abundance in relation to reforestation measures and natural habitat variables in Northen Sweden//Bull. OEPP 1977. Vol. 7. № 2. P. 397-409.
- Tast J. Influence of the root vole, Microtus oeconomus (Pallas) in man-made habitats in Finland//Ann. Zool. Fenn. Vol. 5. P. 62-72.
