Температурный стресс и термопокой семян овощных зонтичных культур. Особенности индукции, проявления и преодоления (Часть II)
")
Автор: Бухаров А.Ф., Балеев Д.Н.
Журнал: Овощи России @vegetables
Рубрика: Семеноводство и семеноведение овощных культур
Статья в выпуске: 4 (21), 2013 года.
Бесплатный доступ
При воздействии на семена овощных зонтичных культур высокотемпературным стрессом в течение 5 или 20 суток происходит снижение активности роста зародыша и ингибирование прорастания семян. Постинкубационное проращивание исследуемых семян различных видов на фоне пониженных стратификационных температур способствует возобновлению роста зародыша, однако, как правило, рост менее интенсивен по сравнению с контролем. Изученные культуры семейства зонтичные, проявили особенности прорастания семян и развития зародышей на различных температурных фонах и в зависимости от времени инкубационного воздействия высокой температуры.
Семена, зародыш, прорастание семян, зонтичные, термопокой, высокотемпературный стресс, термотоустойчивость, чувствительность к высокой температуре, температурный коэффициент, степень недоразвития зародыша (снз)
Короткий адрес: https://sciup.org/14025041
IDR: 14025041 | УДК: 635.1/.7:631.531.01
Temperature stress and seeds dormancy of vegetable umbelliferae crops. Features of introduction, expression, and overcoming (second part)
The influence of high temperature treatment of the seeds of vegetable Umbelliferae crops was studied. Features of seed germination and embryo development depending on incubation time were revealed. High-temperature treatment of seeds during 5 and 20 days inhibits the embryo development and seed germination.
Текст научной статьи Температурный стресс и термопокой семян овощных зонтичных культур. Особенности индукции, проявления и преодоления (Часть II)
Впервой части нашей работы рассмотрены вопросы роста зародыша и способности прораста- ния семян овощных зонтичных культур в условиях высокотемпературного стресса [1]. В данной части мы рассмотрим более углубленно вопросы индукции, проявления и преодоления термопокоя.
Сверхоптимальная температура может повреждать органы и ткани, задерживать рост и вызывать гибель растений. В клетке, находящейся в стрессовом состоянии в первую очередь повреждаются и деградируют белки, инактивируются ферменты, нарушается процесс биосинтеза белков de novo. Одним из механизмов, обеспечивающих резистентность в процессе воздействия высокотемпературного стресса, является синтез белков теплового шока (Heat shock proteins – Hsp) [5, 8]. В экстренных условиях Hsp выполняют работу молекулярного шаперона, опекающего мак- ромолекулы белка, обеспечивают функционирование клеточных структур и поддерживают гомеостаз клетки. Одновременно Hsp обеспечивают правильную сборку олигомерных структур, дезагрегацию неправильно собранных макромолекулярных комплексов и реутилизацию дефектных мономеров с помощью убиквитинов [3].
Изучение проявления термопокоя овощных зонтичных культур, одного из вариантов индуцированного покоя семян, интенсивно проводимые на сельдерее, показали, что при определенных условиях (25оС в условиях темноты) у семян индуцируется термопокой [4]. Выявлено, что различные сорта моркови отличаются по чувствительности семян к сверхоптимальным температурам. Выделены генотипы моркови, обладающие термоустойчивостью и способные прорастать при 35оС и сорта, у которых прорастание нарушается при 30°С[6]. Доказано, что терморезистентность и термопокой наследственно обусловлены, однако гены, контролирующие их, не идентифицированы [7].
Изучение термопокоя, а особенно, процессов протекающих в семенах во время его возникновения, поддержания и нарушения, привлекательно как с теоретической, так и с практической точек зрения. Хотя покой семян является важным условием существования растений и поддержания видового разнообразия в природе, с ним сопряжены большие затруднения в практике культивирования растений.
Материал и методы
Объектом исследований являлись семена укропа (сорт Кентавр), моркови (сорт Рогнеда), петрушки корневой (сорт Любаша), сельдерея корневого (сорт Купидон), любистока лекарственного (сорт Дон Жуан), кориандра (сорт Янтарь) и пастернака (сорт Кулинар), хранившиеся в течение 1 года в лабораторных условиях.
Изучение динамики постинкубационного прорастания семян исследуемых культур проводили на разных температурных фонах, в т. ч.: t = +20°С (st); t = +3°С; t = +3 °C (8 час.) / +20 °C (16 час.), при этом другие факторы: влажность, аэрация, свет (все варианты проращивались без доступа света) были равнозначны. Повторность опыта трехкратная, в каждой повторности исследовали 100 шт. семян.
Измерения длины зародыша во время последующего прорастания проводили с помощью микроскопа «Микромед» при 40 кратном увеличе- нии, с использованием программы Scope Photo. Температурный коэффициент (Q10) для скорости роста зародыша рассчитывали по формуле Вант-Гоффа. Статистический и математический анализ осуществляли по Б. А. Доспехову [2] и с использованием пакета программ Statistica 8.0. Повторность опыта трехкратная, в каждой повторности исследовали не менее 10 шт. семян.
Результаты исследований и обсуждение
Изучение динамики прорастания семян после воздействия высокой температурой на исследуемых температурных режимах проращивания имеет свои особенности в зависимости от сочетания изученных факторов. При инкубации в условиях высоких температур в течение 5 суток сельдерея корневого, пастернака и петрушки корневой и последующем проращивании на фоне стандартной температуры доля проросших семян находится на уровне 48; 41 и 59 %, что на 10; 19 и 9 % соответственно ниже контроля. Доля проросших семян моркови находится на одном уровне с контрольным вариантом и составляет 78 %. Процент проросших семян кориандра и укропа оказывается выше, чем в
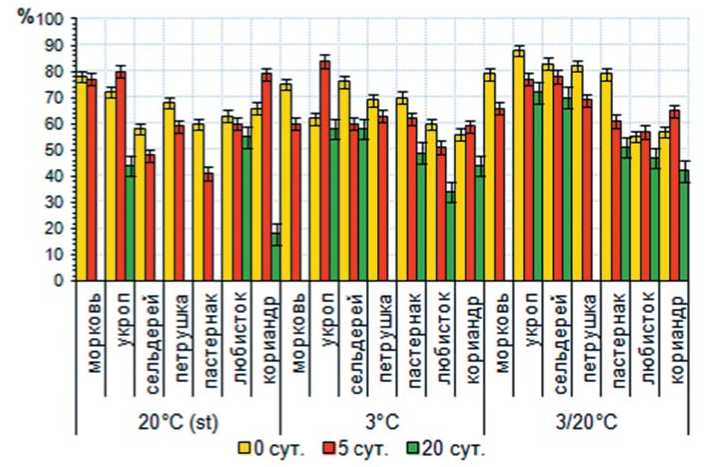
Рис. 1. Доля проросших семян овощных зонтичных культур на различных температурных фонах после воздействия высокотемпературного фактора различной продолжительности
контроле на 13 и 8 % соответственно (рис. 1).
При проращивании в условиях пониженной температуры (t = + 3°C) отмечено, что с увеличением времени инкубации процент проросших семян снижается, особенно отчетливо это заметно у таких культур, как морковь, петрушка корневая, сельдерей корневой, пастернак и любисток лекарственный. При этом доля проросших семян снижается с 69-76% до 34-58% в зависимости от изучаемой культуры. Выделены две культуры – кориандр и укроп, увеличивающие долю проросших семян после 5 суток инкубации на 15 и 22% соответственно по сравнению с контролем.
На рисунке 2 показана тенденция изменения температурного коэффициента (Q10) для скорости роста зародыша изучаемых культур, в зависимости от времени воздействия ингибирующего стрессового фактора. Увеличение времени инкубации семян в условиях высоких температур приводит к снижению температурного коэффициента (Q10) для скорости роста зародыша. Корреляционный и регрессионный анализ показывают, что температурный коэффициент имеют тесную отрицательную зависимость от времени инкубации (коэффициент
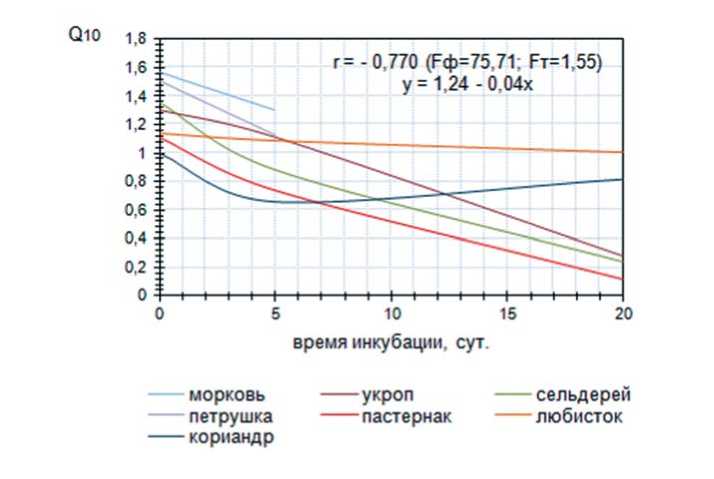
Рис. 2. Динамика изменения температурного коэффициента (Q10) для скорости роста зародыша овощных зонтичных культур в зависимости от продолжительности воздействия высокотемпературного фактора
корреляции имеет значение r = – 0,770) и описывается уравнением регрессии y=1,24-0,04x . При этом выделена группа культур, у которых при увеличении времени воздействия стресса снижение температурного коэффициента происходит плавно, например, у моркови в контроле Q10 составляет 1,57, а после действия высокими температурами в течение 5 суток снижается до 1,30. У семян петрушки корневой Q10 для скорости роста зародыша в контроле составляет 1,51, а под действием инкубации он снижается до 1,13.
Выделены культуры, у которых при увеличении времени действия стресса, температурный коэффициент резко снижается, к таким отнесены сельдерей корневой и пастернак. Для лю-бистока лекарственного не обнаружено значительного влияния времени инкубации на температурный коэффициент для скорости роста зародыша, хотя тенденция снижения последнего имеет место. Температурный коэффициент роста зародыша кориандра в контроле составляет 1,00, затем, под действием высокой температуры в течение 5 суток он резко снижается и составляет 0,66, а после 20 суток инкубации Q10 возрастает до 0,82.
Воздействие переменными темпе- ратурами при постинкубационном проращивании оказывает не однозначное влияние на число проросших семян. При последовательном увеличении времени действия стрессора морковь, петрушка корневая и сельдерей корневой снижают процент проросших семян (на 1-15; 9-13; 10-18 % соответственно). Доля проросших семян пастернака в контроле составляет 79 %, после 5 суток воздействия этот показатель снижается до 61 %, а после 20 суток до 51 %. Процент проросших семян у любистока лекарственного и кориандра, после инкубации в течение 5 суток, возрастает по сравнению с контролем на 2 и 8 %, а при 20 суточном воздействии снижается на 8 и 15 % соответственно. Следует отметить, что в контроле большинство изученных культур прорастают в широком интервале температур, а семена, испытавшие высокотемпературный стресс, успешно прорастают, как правило, только при пониженной или переменной температурах.
Действие стрессового фактора необходимо рассматривать не только с точки зрения его повреждающего действия, но и сточки зрения ответной реакции на него организма. С этой позиции среди изучаемых культур анализ полученных результатов позволяет вы- делить несколько групп культур по отношению к воздействию высокой температурой. Культуры, на которых действие высоких температур при длительном влиянии губительно, при этом происходит автолиз семян: морковь и петрушка корневая. Группа культур (укроп, любисток лекарственный), которые оказались индифферентны к воздействию высоких (t = + 30°C) температур в период прорастания семян. При этом во время действия высокотемпературного фактора у них наблюдается торможение прорастания, но при наступлении благоприятных условий ростовые процессы быстро возобновляются. Культуры, которые под действием высоких температур впадают в термопокой (сельдерей корневой, пастернак и кориандр) с разной глубиной, которая увеличивается при более продолжительном воздействии ингибирующего фактора.
На схеме (рис. 3) представлены особенности роста зародыша отдельных культур из выделенных нами групп по реакции на высокотемпературный стресс и степени проявления термопокоя.
При обсуждении роста зародыша культур, входящих в первую группу «погибшие семена» остановимся подробнее на моркови. В семенах, не испытавших воздействие высокой температуры, зародыш растет интенсивно. Скорость роста зародыша составляет от и 0,10 до 0,14 мм/сутки. Ранее нами предложен показатель «степень недоразвития зародыша», рассчитываемый как отношение величины, на которую увеличился зародыш в процессе прорастания к средней длине зародыша, при которой началось прорастание семян, выраженное в процентах [2]. В условиях стандартного температурного режима степень недоразвития зародыша составляет 25%, при переменной температуре – 30%, а при пониженной достигает 37%, что обусловлено изменением линейных размеров зародыша к моменту прорастания.
В течение первых 5 суток инкубации
при повышенной температуре скорость роста зародыша резко падает и составляет 0,02 мм/сутки, при этом зародыш продолжает рост и увеличивается на 8%. При переносе семян на разные температурные фоны для постинкубационного проращивания ростовые процессы возобновляются. В условиях стандартного температурного режима скорость роста по сравнению с другими вариантами увеличивается и составляет 0,05 мм/сутки. Пониженная температура тормозит скорость роста, которая в этом случае составляет 0,03 мм/сутки, при этом сте-
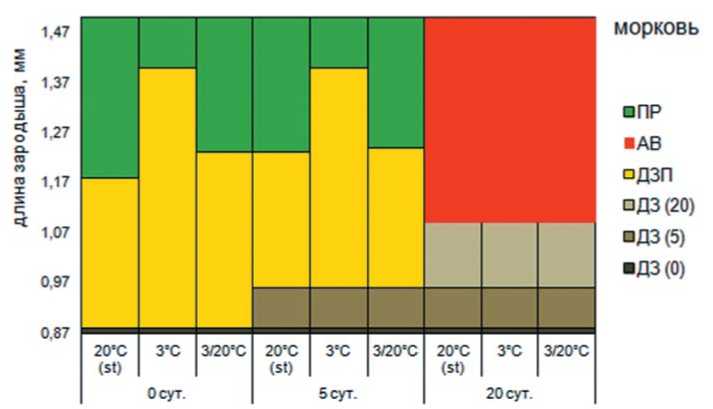
пень недоразвития зародыша увеличивается.
Если воздействие высоких температур во время инкубации семян продолжается 20 суток, то скорость роста зародыша снижается до 0,02 мм/сутки и длина зародыша увеличивается еще на 24 %, достигая 1,08 мм. Таких темпов роста, возможно, было бы достаточно для наступления прорастания не обработанных семян. Однако в семенах подвергнутых длительному воздействию высоких (экстремальных, сверхоптимальных) температур зародыш погибает, что возможно связано с недостаточной жаростойкостью семян
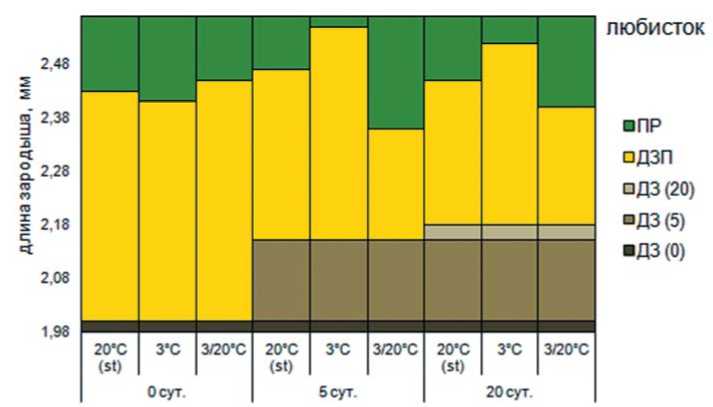
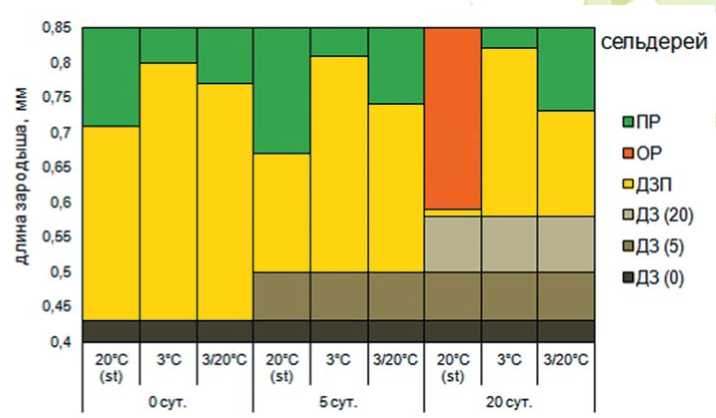
Рис. 3. Изменение размера зародыша под влиянием высокотемпературного стресса и последующих условий проращивания: ДЗ (0) – длина зародыша в семенах не прошедших инкубацию в условиях высокой температуры, мм; ДЗ (5) – длина зародыша при действии высокой температуры в течение 5 суток, мм; ДЗ (20) – длина зародыша при действии высокой температуры в течение 20 суток, мм; ДЗП – длина зародыша перед прорастанием семян, мм; ПР – длина зародыша при прорастании семян, мм; ОР – остановка роста зародыша; АВ – автолиз
моркови и петрушки, а так же возможна гибель семян при недостатке кислорода во время инкубации в условиях высокотемпературного стресса [6].
Рост зародыша в семенах второй группы культур, которые индифферентны к действию повышенной температуры, показан на примере любис-тока лекарственного (рис.5). Необработанные семена в различных температурных условиях проращивания имеют степень недоразвития зародыша в пределах 18-19%. Под влиянием инкубации в течение 5 суток размеры зародыша увеличиваются на 7,5 %, достигая 2,15 мм. Однако степень недо- развития зародыша при этом увеличивается до 22%. В течение последующих 15 суток инкубации в условиях высоких температур доразвитие зародыша в семенах происходит со скоростью 0,008 мм/сутки, а длина зародыша увеличивается менее чем на 1 %, достигая 2,16 мм. Таким образом, темпы роста резко замедляются, однако, семена сохраняют жизнеспособность.
В течение постинкубационного проращивания при различных температурных режимах скорость роста зародыша несколько снижается и варьирует от 0,04 до 0,05 мм/сутки, а степень недоразвития зародыша составляет 18-21%. Сочетание высокотемпературной обработки семян и последующего проращивания при переменной температуре стимулирует прорастание при меньшем размере зародыша, что положительно сказывается на показателях Tmax ν и T50.
На примере сельдерея корневого, яркого представителя группы культур имеющих термопокой, видно как растет зародыш в семени. В контроле зародыш начинает свой рост, на всех температурных режимах, с длинны 0,43 мм. Под действием стандартного (t = + 20°C) режима температур зародыш растет нормально и к 5 суткам проращивания достигает длины 0,71 мм или 39% абсолютного прироста зародыша, после чего наступает прорастание семян.
При действии переменной температуры доразвитие зародыша затягивается, т. к. для наступления прорастания необходимо, чтобы средняя длина зародыша увеличилась на 44%, для чего требуется больше времени (6 суток), хотя скорость роста зародыша при этом составляет 0,06 мм/сутки, что на 0,01 мм/сутки выше, чем при стандартном режиме проращивания. Достигая длины 0,77 мм, зародыш прорастает.
Пониженная температура оказывает тормозящее действие на рост зародыша, при этом прирост зародыша, который необходим для прорастания, составляет 46%, скорость роста зародыша снижается до 0,03 мм/сутки, что значительно увеличивает времени прорастания.
В процессе инкубации при температуре 30°С в течение 5 суток, длина зародыша увеличивается на 14%, скорость роста зародыша в этот период находится в пределах 0,02 мм/сутки. При перенесении семян в условия стандартных температур рост зародыша продолжается, но его скорость не превышает 0,01 мм/сутки. При такой скорости для достижения длины, необходимой для прорастания, требуется больше времени по сравнению с необработанными семенами.
Под действием постинкубационного проращивания в условиях пониженной температуры рост зародыша происходит со скоростью 0,02 мм/сутки. Не смотря на то, что скорость выше, чем в стандартных температурных условиях, прорастание наступает позже, в связи с тем, что прирост зародыша должен составить 47%. Действие переменной температуры так же увеличивает степень недоразвития зародыша по сравнению со стандартным температурным режимом, при этом она находится в пределах 42%, а скорость роста зародыша составляет 0,02 мм/сутки.
Инкубация семян сельдерея корневого при температуре 30оС в течение 20 суток приводит к полной остановке рос- та зародыша, при достижении им длины 0,58 мм. Прирост за это время составил 0,08 мм. После перенесения семян в условия стандартных (t = + 20°C) температур наблюдается возобновление роста зародыша, его длина увеличивается на 3% и достигает 0,60 мм. Однако в дальнейшем рост зародыша останавливается, и прорастание не происходит. Действие пониженной и переменной температуры позволяет преодолеть барьер, обусловленный остановкой роста. Использование переменной температуры более благоприятно, поскольку увеличивается интенсивность роста зародыша, обеспечивает начало прорастания при меньшей длине зародыша, а, следовательно, степень доразвития значительно снижается.
Заключение
По отношению к высокотемпературному стрессу выделены три группы культур: морковь и петрушка корневая – чувствительные культуры, под действием высокотемпературного фактора происходит гибель семян. Укроп, любисток лекарственный – индефе-рентные культуры по отношению к высокой температуре. Сельдерей корневой, пастернак и кориандр – культуры обладающие способностью переходить в состояние термопокоя различной глубины, и тем самым избегать губительного действия стрессора. Для выведения семян, впавших в состояние термопокоя, требуется использование воздействия пониженной температуры различное время в зависимости от исследуемой культуры.
Список литературы Температурный стресс и термопокой семян овощных зонтичных культур. Особенности индукции, проявления и преодоления (Часть II)
- Бухаров А.Ф., Балеев Д.Н. Температурный стресс и термопокой семян овощных зонтичных культур. Особенности индукции, проявления и преодоления (Часть I)//Овощи России.-2013.-№2(19).-С.36-41.
- Доспехов Б.А. Методика полевого опыта. -М.: Агропромиздат.-1985.-351 с.
- Кулаева О.Н. Белки теплового шока и устойчивость растений к стрессу//Соровский образовательный журнал.-1997.-№2.-С. 513.
- Bouzo C., Favaro J. Improving the germination of celery seeds at high temperature//Journal of agriculture and social sciences. -2007. -P.1813-2235.
- Helm K.W., Petersen N. S., Abernethy R. H. Heat Shock Response of Germinating Embryos of Wheat//Plant Physiol.-1989. -Vol. 90.-P.598-605.
- Nascimento W. M., Vieira J. V., Silva G. O. Carrot seed germination at high temperature: effect of genotype and association with ethylene production//Hortscience.-2008. -№43 (5). -P. 1538-1543.
- Senthil-Kumar M., Ganesh K., Srikanthbabu V., Udayakumar M. Assessment of variability in acquired thermotolerance: Potential option to study genotypic response and the relevance of stress genes//J. Plant Physiol.-2007.-Vol. 164.-P. 111-125.
- Tariq M., Waseem S., Bilal H. A. An overview on the small heat shock proteins//African Journal of Biotechnology.-2010. -Vol. 9 (7).-P. 927-949.
