Термографическая регистрация тепловых эффектов у растений при холодовом стрессе

Автор: Ковчавцев А.П., Курышев Г.Л., Базовкин В.М., Половинкин В.Г., Настовьяк А.Е., Финников К.А., Корзун А.М., Колесниченко А.В., Войников В.К.
Журнал: Журнал стресс-физиологии и биохимии @jspb
Статья в выпуске: 4 т.7, 2011 года.
Бесплатный доступ
В настоящей работе изучается возможность длительного наблюдения тепловых эффектов у растений термографическим способом. Обсуждаются проблемы потери точности измерений при низких температурах и способ калибровки камеры по температуре. Исследуется возможность проведения быстрых измерений в процессе охлаждения растений. Устанавливается диапазон температур, приводящий к гибели растений озимой пшеницы.
Растения, термография, температурный стресс
Короткий адрес: https://sciup.org/14323571
IDR: 14323571
Текст научной статьи Термографическая регистрация тепловых эффектов у растений при холодовом стрессе
В настоящей работе изучается возможность длительного наблюдения тепловых эффектов у растений термографическим способом. Обсуждаются проблемы потери точности измерений при низких температурах и способ калибровки камеры по температуре. Исследуется возможность проведения быстрых измерений в процессе охлаждения растений. Устанавливается диапазон температур, приводящий к гибели растений озимой пшеницы.
Изучение адаптивных возможностей растений и их устойчивости к неблагоприятным воздействиям окружающей среды относится к центральным проблемам биохимии и физиологии растений. Особенно актуальными в данном направлении остаются исследования, направленные на установление механизмов адаптации растительных объектов к низким температурам (Knutson, 1974; Vojnikov et al, 1984; Moynihan et al, 1995; Breidenbach et al,
1997; Kolesnichenko et al, 2003; Войников, Колесниченко, 2011).
Тепловое изображение объектов целесообразно наблюдать с помощью тепловизионной камеры, например, как в (Курышев и др, 1998). Изображение формируется инфракрасным объективом и регистрируется с помощью фоточувствительной матрицы (128х128 элементов с шагом 50 мкм), установленной в фокальной плоскости объектива. Элементы матрицы преобразуют кванты света спектрального диапазона 2,65-3,05 мкм в электрические заряды, которые считываются мультиплексором, усиливаются, предварительно обрабатываются электронной схемой и передаются в компьютер. В результате на экране монитора формируется тепловизионное изображение объекта или термограмма. При проведении измерений температуры объекта в области отрицательных величин следует учитывать, что при уменьшении температуры падает излучательная способность объекта (в соответствиии с формулой Планка) и, соответственно уменьшает- ся величина полезного сигнала на элементах матрицы.
Калибровка тепловизионной камеры
На Рис.1 показана минимально разрешимая разность температур (МРРТ) элементом матрицы от температуры наблюдаемого объекта, когда шум элемента ограничивается величиной комнатного фона. Видно, что при охлаждении объекта температурное разрешение фотоприемных элементов камеры ухудшается. При температуре ~ -10 0С температурное разрешение элементов ~ 0,16 0С.
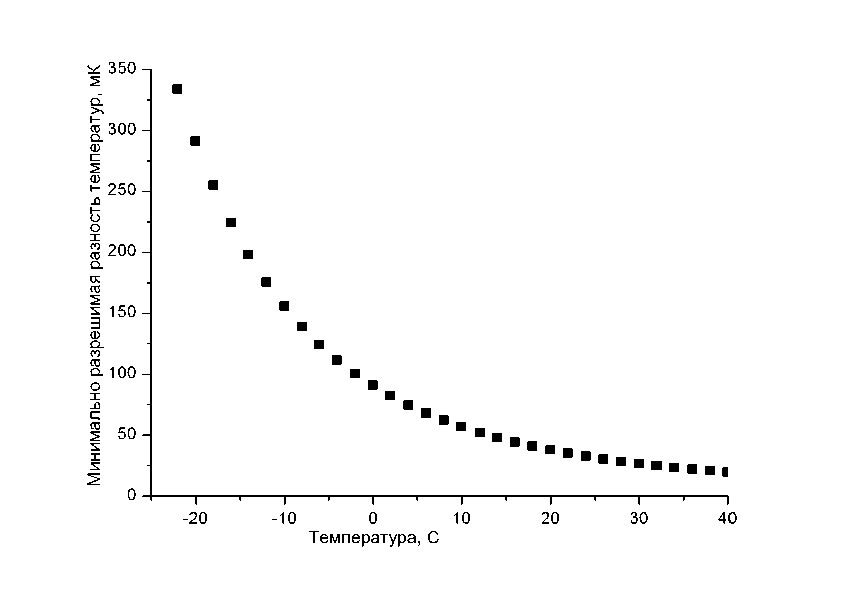
Рисунок 1: Температурная зависимость МРРТ.
В регистрируемые величины значительный вклад вносит погрешность, связанная с изменением температуры корпуса прибора, и как следствие изменением потока фонового излучения. Для компенсации погрешности необходимо проводить перед измерением калибровку прибора по встроенным шторкам - имитаторам потока излучения. Уменьшить погрешность прибора, вносимую изменением фоновой температуры, можно, помещая тепловизор в термостатированные условия. Однако, ошибку в регистрации тепловизором температуры в непрерывном режиме, связанную с воздействием низкотемпературного потока излучения самого объекта исследования на оптическую систему прибора, избежать невозможно. Она устраняется только в результате перекрывания потока излучения теплоизолирующей шторкой, которая открывается на незначительный промежуток времени для регистрации термограммы. Но прерывистый режим регистрации в ручном режиме требует больших трудозатрат и значительно затрудняет проведение серийных экспериментов. На практике, технология регистрации термограмм в области низких и отрицательных температур в непрерывном режиме может давать значительную ошибку в точности измерения (до нескольких градусов) и позволяет проводить лишь качественную оценку изменения температур растительных объектов при низкотемпературном стрессе.
Вклад в общую погрешность дает и ошибка, связанная с нелинейностью и неоднородностью передаточных характеристик фотоприемного устройства (ФПУ), входящего состав тепловизионной камеры. Нелинейность передаточных характеристик элементов ФПУ может быть обусловлена разными причинами: как принципиальными особенностями организации ФПУ, так и особенностями технической реализации. Нелинейность приводит также к невозможности простым вычитанием выделить из полного сигнала составляющую, обусловленную фоновым излучением. Проблему можно решить путём перевода индивидуальных сигналов элементов тепловизионной матрицы в некоторую универсальную характеристику, которую можно считать пропорциональной потоку фотонов. Для определения такой характеристики следует иметь некоторый эталонный источник со строго определенным потоком фотонов, например, абсолютно черное тело (АЧТ). Однако в этом случае для определения потока фотонов, регистрируемых ФПУ, необходимо знать спектральные характеристики элементов. От такого недостатка свободен дифференциальный метод нахождения связи сигнала с интенсивностью потока фотонов, предложенный ранее для тепловизионного микроскопа (Курышев и др, 2011). Суть модифицированного метода для калибровки тепловизионной камеры в следующем: если измерить изменение фотосигнала § S при модуляции потока фотонов на фиксированную величину § I для разных значений основного потока и, следовательно, сигнала S, то мы получим значения величины: § S = F(S)81
или в дифференциальной форме: dS/dI = F ( S ) . Обращая это уравнение интегрированием получим функцию I(S), преобразующую значение сигнала в величину пропорциональную интенсивности потока фотонов (функцию интенсивности). Примерная схема измерений, позволяющая реализовать такие измерения, представлена на Рис.2.
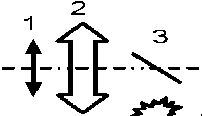
Рисунок 2: Схема измерения дифференциальной чувствительности ФПУ.
1 - матрица ФПУ, 2 – объектив. 3 – светоделительное пластинка, 4 – источник модулирующего излучения, 5 – источник основного излучения.
Точечный источник (например, инфракрасный светодиод) помещается так, чтобы его отражение на светоделительной пластинке находилось примерно в фокусе объектива. В этом случае за объективом будет практически параллельный пучок, обеспечивающий равномерную засветку элементов ФПУ. Размеры и расположение источника основного излучения (АЧТ) также выбираются из условия равномерной засветки всех элементов ФПУ.
Для определения функции интенсивности следует найти подходящее аппроксимирующее выражение для измеренной функции F(S). Модельные расчеты возможного вида такой зависимости предсказывают её экспоненциальное поведение на краях диапазона. Однако использование экспонент в аппроксимирующем выражении для F(S) приводит к сложной формуле для I(S), вычисление которой в реальном времени для всех элементов ФПУ практически невозможно. Поэтому предлагается использовать аппроксимацию полиномом для функции 1/F(S). Достаточная точность аппроксимации достигается при использовании полиномов четвертой – пятой степени. В этом случае требуемая зависимость интенсивности от сигнала I(S) - полином пятой – шестой степени.
Используемый способ определения функции интенсивности I(S) дает два свободных параметра: во-первых, это свободный член, возникающий при интегрировании, а, во-вторых, масштабный множитель. Эти два параметра можно доопределить из следующих условий:
-
• зависимость интенсивности от времени накопления должна экстраполироваться к нулевому значению при времени накопления равном нулю,
-
• наклон зависимости I(S) примерно равен 1, а точнее, I(S2) – I(S1) = S2 – S1, где S1 = min(S) + 0.1 A , S2 = max(S) - 0.1 А , А = max(S) - min(S).
Полученная функция интенсивности позволя- ет провести корректное вычитание фонового сигнала. Для этого необходимо провести измерения зависимости сигнала So от объекта с фиксированной температурой и сигналов при закрытых шторках (Sd, Sm) при разных значениях температуры корпуса. Зависимости интенсивности от объекта Io = I(So) от соответствующих интенсивностей шторок Id = I(Sd), Im = I(Sm) или их разности Id-Im должны быть линейными с коэффициентами определяемыми отношениями соответствующих эффективных (с учетом коэффициентов излучательных способностей) площадей шторок и объектива. Знание этих коэффициентов позволяет исключить влияние интенсивности фонового излучения простым вычитанием соответствующего произведения интенсивности на коэффициент.
Получив свободное от влияния фонового излучения значение функции интенсивности, мы можем перевести его в значение эквивалентной температуры АЧТ. Для реально используемых диапазонов температур (263–320) К зависимость числа фотонов от температуры с хорошей точностью может быть представлена выражением: I ph = I i exp (- E ef/ kT ) . Требуемая зависимость I ph (Т) может быть измерена по схеме (рис.2) без светоделительной пластинки или (при известном коэффициенте пропускания пластинки) определена из измерений дифференциальной чувствительности. На практике эти зависимости с хорошей точностью могут быть представлены выражением:
Iph = 1 ( T) = I0 + I exp (- Eef/kT) . Присут ствие постоянного слагаемого в этом выражении, отражает вклад темновой генерации в накопленный заряд. Следует заметить, что определяемая таким образом температура есть эквивалентная температура АЧТ. Различие излучательных способностей объектов при их одинаковой температуре приводит к видимому различию сигналов и, следовательно, эквивалентных температур. Введенная в данной работе функция интенсивности позволяет, во-первых, учесть этот эффект если известно, что излучательную способность данного объекта можно охарактеризовать некоторым коэффициентом черноты, а во вторых, определить эти коэффициенты, если температура объекта известна. На практике, после реализации такого способа калибровки, температуры, полученные тепловизионной камерой, привязываются в нескольких контрольных точках к температуре наблюдаемого объекта с помощью платиновых термометров. В качестве реперного объекта использовались медные пластинки покрытые чернением тонкого слоя осажденной сажи, полученной при сгорании скипидара. В диапазоне температур от -7 до +10 0С точность совпадения температурных показаний тепловизора и термосопротивлений достигает величины ~ 0,15 К.
Камера быстрого охлажения
Для обеспечения возможности изменения скорости нагрева экспериментальных образцов была опробована камера, поперечное сечение которой показано на рис.3.
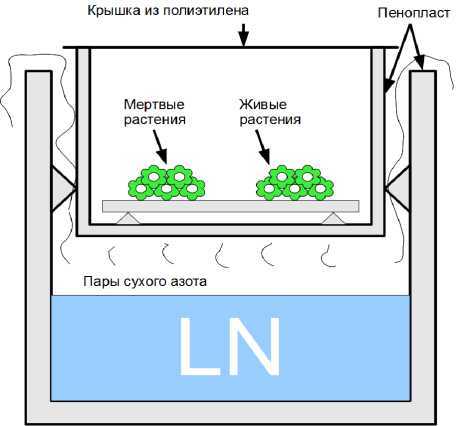
Рисунок 3: Экспериментальная климатическая камера
Растения располагались на дне тонкостенного пенопластового контейнера и охлаждались парами жидкого азота из внешнего контейнера. Скорость охлаждения регулировалась уровнем жидкого азота (LN) во внешнем контейнере. Для уменьшения конвекционных потоков воздуха рабочий контейнер сверху закрывался тонкой пленкой, прозрачной для ИК-излучения. Наблюдение за изменением температуры проводилось тепловизионной камерой сквозь пленку сверху. Тепловизионная камера размещалась на расстоянии 0,7 – 1 м над рабочим контейнером и дополнительно не термостабилизировалась. Дополнительно в экспериментальных образцах размещались два платиновых термосопротивления для контроля температуры.
Экспериментальные результаты.
В качестве экспериментальных образцов использовались трех-четырех недельные растения озимой пшеницы ( Triticum aestivum l., сорт
Иркутская) и пырея ползучего (Agropyrum repens L). Часть образцов пшеницы умертвлялась погружением в горячую воду (~ 100 0C). Затем мертвые и живые растения термостабилизировались выдержкой в воде при комнатной температуре, и осушались до примерно одинаковой влажности. Из живых и мертвых растений связывались пучки одинаковой массы. В середину пучков вводились платиновые термодатчики и пучки помещались в рабочую камеру. После термостабилизации растений при открытой камере и отсутствии жидкого азота во внешнем контейнере растения принимали температуру ~ 23,8 0С. После стабилизации рабочая камера закрывалась прозрачной пленкой. Через пять минут температура живых растений поднималась до ~ 25,3 0C (по-видимому вследствие парникового эффекта). Разность температур между живыми и мертвыми растениями достигала величины 1,5 С. При дальнейшей выдержке разность температур между живыми и мертвыми растениями уменьшается за счет распространения тепла от живых растений. После термостабилизации растений в области комнатных температур растения охлаждались. Кривые охлаждения растений пшеницы показаны на рис. 4. Видно, что при охлаждении растений до температур вблизи 0 0C живые растения во времени охлаждаются медленнее мертвых, что связано с выработкой тепла живыми растениями. После выдержки при нулевых температурах в течение ~ 1 часа проводилась вторая фаза охлаждения до более низких температур. На второй фазе скорость охлаждения на живых растениях замедляется, поскольку тепловыделение растений компенсирует охлаждение за счет отвода тепла холодным воздухом к стенкам камеры. При понижении температуры ниже -7 С тепловыделения живыми растениями явно не достаточно для компенсации тепловых потерь растениями. При этом температура живых растений быстро спадает и уравнивается с температурой мертвых растений. По-видимому на этой стадии тепловыделение живыми растениями прекращается.
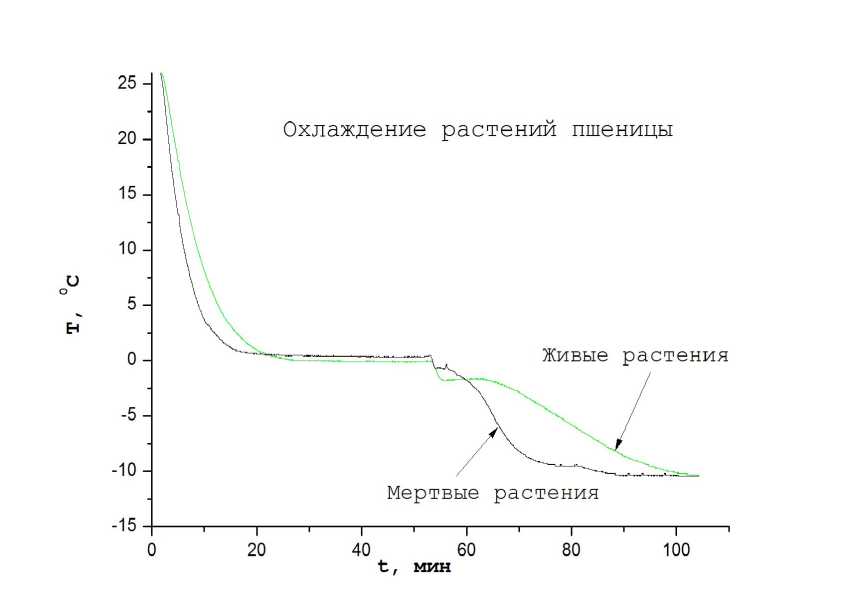
Рисунок 4. Динамическая кривая охлаждения растений озимой пшеницы.
Измерялась также кинетическая кривая охлаждения растений пырея. Контрольная группа растений пырея убивалась охлаждением растений до температуры жидкого азота. После термостабилизации растений при комнатной температуре убитые растения отличались от живых растений наличием насыщенного темнозеленого цвета. Живые растения при этом имели светлозеленую окраску. На рис.5 показана кинетическая кривая охлаждения растений.
На кинетических кривых охлаждения можно выделить несколько характерных участков. После термостабилизации растений при комнатной температуре в течении ~ 50 мин проводилось охлаждение до нулевых температур. При этом тепловыделение живых растений заметно по наличию характерной затяжки кинетической кривой в диапазоне температур от +10 до -1,5 С. В области ~ -1 C наблюдается небольшое повышение температуры мертвых растений, по-ви- димому, за счет выделения тепла при образовании льда в растениях. Затем на всем протяжении кинетических кривых наблюдается тепловыделение живыми растениями. Следует отметить, что в отличие от растений озимой пшеницы тепловыделение живыми растениями пырея сохраняется и при самых низких температурах, когда тепловыделение живыми растениями пшеницы прекращается.
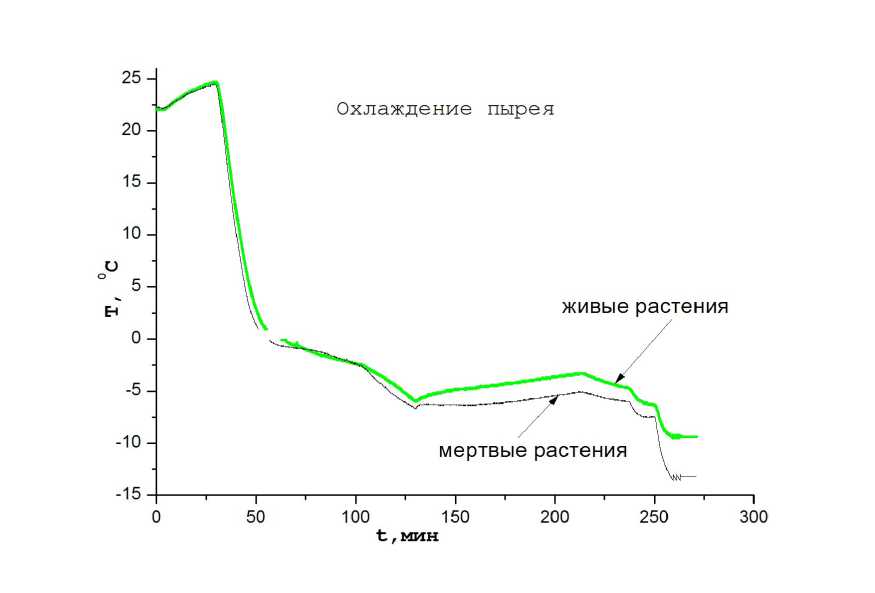
Рисунок 5. Динамическая кривая охлаждения растений пырея.
Таким образом, на основании полученных результатов можно сделать следующие выводы:
-
1. Опробована климатическая камера, позволяющая проводить быстрые эксперименты по влиянию холодовых стрессов на живые растения. Скорость охлаждения растений можно менять в достаточно широком диапазоне.
-
2. На кинетических кривых охлаждения живых растений озимой пшеницы можно выделить несколько характерных участков, характеризующих теплообмен живых растений с окружающей средой, причем охлаждение живых растений можно достаточно быстро проводить до прекращения тепловыделения растениями.
-
3. При охлаждении живых растений пырея наблюдаются кинетические кривые отличающиеся от растений пшеницы только количественными параметрами. При выдержке растений при температуре окружающей среды ниже -12 C растения сохраняют жизнеспособность, т.е. выделяют тепло.
Авторы выражают глубокую признательность Пановой З.В. за помощь в проведении экспериментов.
Работа выполнена при поддержке гранта Конкурса интеграционных проектов СО РАН, №45 (2009-2011г.г.).
Список литературы Термографическая регистрация тепловых эффектов у растений при холодовом стрессе
- Войников В.К., Колесниченко А.В. (2011) Синтез стрессовых белков в растениях при гипотермии. Lambert Academic Publishing, Saarbruken, Deutschland, 264 p.
- Курышев Г.Л., Ковчавцев А.П., Вайнер Б.Г., Гузев А.А., Базовкин В.М., Строганов А.С., Субботин И.М., Захаров И.М, Ефимов В.М., Постников К.О., Ли И.И., Валишева Н.А., Панова З.В. (1998) Медицинский тепловизор на основе матричного ФПУ 128х128 для диапазона длин волн 2,8 -3,05 мкм Автометрия 4: 5-12.
- Курышев Г.Л., Мжельский И.В., Настовьяк А.Е., Половинкин В.Г. (2011) Калибровка фотоприемного устройства в составе ИК-микроскопа Автометрия 47(5): 103-108.
- Breidenbach R.W., Saxton M.J., Hansen L.D., Criddle R.S. (1997) Heat generation and dissipation in plants: Can the alternative oxidative phosphorylation pathway serve a thermoregulatory role in plant tissues other than specialized organs? Plant Physiol 114: 1137-1140
- Knutson R. M. (1974) Heat production and temperature regulation in eastern skunk cabbage Science, 186: 746-747.
- Kolesnichenko A.V., Pobezhimova T.P., Grabelnych O.I., Tourchaninova V.V., Korzun A.M., Koroleva N.A., Zykova V.V., Voinikov V.K. (2003) Difference between the temperature of non-hardened and hardened winter wheat seedling shoots during cold stress J. Thermal Biology. 28: 235-244.
- Moynihan MR, Ordentlich A, Raskin I (1995) Chilling-induced heat evolution in plants. Plant Physiol 108: 995-999
- Vojnikov V., Korzun A., Pobezhimova T., Varakina N. (1984) Effect cold shock on the mitochondrial activity and on the temperature of winter wheat seedlings Biochim. Physiol. Pflanz. 179: 327-330.
