The impact of experimental hypoxia and subsequent normoxia on the content of some ions and markers of physiological stress-adaptation in gastropod species Lymnaea stagnalis

Author: Lubyaga J.А., Axenov-gribanov D.V., Madyarova E.V., Gurkov А.N., Lozovoy D.V., Homich A.S., Golubev A.P., Timofeyev М.А.
Journal: Журнал стресс-физиологии и биохимии @jspb
Article in issue: 4 т.10, 2014.
Free access
The purpose of this study was to evaluate the influence of hypoxia and subsequent normoxiaon the maintenance of respiratory pigment hemocyanin, total protein, lactate and some ions (Na +, K +, Ca 2+, NH 4+, Mg 2+) in the mantle liquid in palaearctic gastropod species Lymnaea stagnalis. It was shown that short-term experimental hypoxia leads to the activation of the physiological mechanisms of stress adaptation in widespread Palaearctic eurybiotic gastropod species and does not lead to the activation of the stress-resistance mechanisms on the biochemical and molecular levels.
Gastropods, hypoxia, ion regulation, normoxia, stress
Short address: https://sciup.org/14323906
IDR: 14323906
Text of the scientific article The impact of experimental hypoxia and subsequent normoxia on the content of some ions and markers of physiological stress-adaptation in gastropod species Lymnaea stagnalis
Экологические характеристики видов в значительной степени определяются физиологическими и биохимическими процессами, происходящими на уровне клеточного метаболизма (Grieshaber et al ., 1994). Одними из ключевых показателей, отражающих состояние организма и происходящие в нём процессы, являются механизмы ионной регуляции и физиологические показатели стресс-адаптации (Frederich et al. , 2000). Исследование данных механизмов стресс-ответа у видов, обитающих в мелких пресноводных водоемах, представляет особый интерес, поскольку такие экосистемы в наибольшей степени подвержены различным антропогенным нагрузкам. В частности, летнее повышение температуры водоема приводит к понижению содержания растворенного кислорода в воде вследствие его диффузии в атмосферу и расходованием на окисление присутствующих в воде органических и неорганических загрязнителей (Konstantinov, 1986).
Эврибионтный палеарктический вид пресноводных легочных моллюсков Lymnaea stagnalis ( Linnaeus, 1758) является широко распространенным модельным объектом для исследования влияния условий среды обитания на физиологические и биохимические показатели стресс-метаболизма (Wijsman et al ., 1998; Gnatishina et al ., 2011; Coutellec et al ., 2006). В теплый период года L. stagnalis дышит преимущественно атмосферным воздухом, который поступает в непарное легкое через крупное закрывающееся дыхательное отверстие
(пневмостом). По этой причине в природе он встречается преимущественно в прибрежных или мелководных участках (до 1,5 – 2 м) водоемов, где имеет возможность периодически подниматься к поверхности воды. Отсюда содержание растворенного кислорода в воде обычно не рассматривается в качестве лимитирующего фактора среды для L. stagnalis . Однако данный вид поглощает атмосферный кислород через легкое и растворенный в воде кислород, через кожные покровы примерно в одинаковых количествах. При этом кислород, поглощенный легким, поступает в гемолимфу, а проникший диффузно через кожу (обычно не глубже 1 мм) - в тканевую жидкость (Stadnichenko et al. , 1996). При необходимости, например, при резком повышении уровня воды в водоеме и в зимний период, когда водоем покрывается льдом, L. stagnalis полностью переходит на дыхание растворенным в воде кислородом, заполняя легкое водой и поэтому, может оказываться в условиях гипоксии. Для оценки адаптивного потенциала L. stagnalis в условиях резких колебаний содержания растворенного кислорода в воде природных водоемах нами проведена экспериментальная оценка влияния гипоксии на содержание дыхательного пигмента гемоцианина, лактата и некоторых ионов (Na+, K+, Ca2+, NH4+, Mg2+) в его мантийной жидкости.
MATERIALS AND METHODS
Сбор взрослых представителей гастропод для данного исследования проводили с водной растительности у поверхности озера, расположенного в черте г. Иркутска на о. Юность (географические координаты: 52.268001, 104.280998; 52.268086, 104.284055.). Оценку влияния гипоксии проводили посредством экспонирования моллюсков в условиях экспериментальной гипоксии при содержании кислорода в воде 3 мг/л в течение шести часов и последующей нормоксии при 8-9 мг/л в течение трех часов при температуре вылова. При проведении экспериментов была использована вода из водоема в местах отлова моллюсков. Эксперименты проведены в гипоксических склянках с притертой крышкой при температуре вылова организмов – 20-22°С. Условия гипоксии достигались путем отсасывания кислорода из воды с применением мембранного насоса.
Оценку содержания кислорода в воде проводили с помощью оксиметра Extech Heavy Duty Dissolved Oxygen Meter 407510 (США).
В ходе исследования проведена оценка содержания дыхательного пигмента гемоцианина, общего белка, лактата и некоторых ионов (Na+, K+, Ca2+, NH4+, Mg2+) в мантийной жидкости палеарктического вида гастропод. Мантийную жидкость для исследования получали путем легкого укола ноги моллюска энтомологической булавкой (Stadnichenko et al. , 1999).
Оценку содержания гемоцианина проводили при длине волны 335 нм, предварительно разбавив пробы в 100 раз и осадив форменные элементы центрифугированием в течение 5’ при 16100 rpm, 5°С (Frederich et al., 2000). Содержание гемоцианина и лактата проводили с помощью спектрофотометра CaryVarian – 50 (США). Измерение концентрации лактата проведено по стандартным протоколам «Лактат-витал» (Витал диагностикс, СПб) при длине волны 505 нм (Axenov-Gribanov, 2013).
Оценку содержания ионов (Na+, K+, Ca2+, NH4+, Mg2+) проводили с применением метода безреагентной ионной хроматографии на приборе ICS – 2000 (Германия). Измерение концентрации ионов проведено по калибровочным графикам для каждого анализируемого иона.
Все эксперименты проведены в 7 биологических повторностях. Биохимический анализ каждой пробы был проведен в 3-х аналитических измерениях. Оценку достоверности проводили, используя двувыборочный u-критерий Манна-Уитни. Статистический анализ проводили с использованием программы Statistica 8.0. На диаграммах указаны доверительные интервалы. * -обозначены случаи достоверного отличия значений от контрольного при доверительной вероятности – 0,95.
RESULTS AND DISCUSSION
В условиях экспериментальной гипоксии происходит повышение содержания гемоцианина с 62,31±10,95 мг/мл до 113,15,95±21,84 мг/мл (рис. 1А), что свидетельствует об активации физиологических реакций стресс-адаптации и об интенсификации транспорта кислорода в стрессовых условиях (Morris et al., 1988: Lorenzon et al., 2008). При этом важно отметить, нормоксия в течении 3 часов после 6 часовой гипоксии не приводила к восстановлению уровня гемоцанина до контрольных значений.
Принимая во внимание, что в ходе работы не отмечали изменения содержания лактата в мантийной жидкости относительно контрольных величин – 0,115±0,02 мМоль/г (рис. 1Б), это указывает на то, что данный тип стрессового воздействия не приводит к увеличению доли анаэробных процессов в клетке (Grieshaber et al ., 1994; Ekkekakis et al ., 2004) и существенных перестроек в метаболизме гастропод, затрагивающих биохимические и молекулярные механизмы стресс-адаптации не происходит.
Материалы по оценке содержания ионов Na+, K+, Mg2+ , NH4+ и Ca2+ в мантийной жидкости контрольной выборки L. stagnalis представлены на рисунке 2. Содержание ионов составило: Na+ -146,29±26,55 мМоль/л; K+ - 4,49±0,33 мМоль/л; Mg2+ - 14,37±2,41 мМоль/л; NH4+ - 1,04±0,11 мМоль/л; Ca2+- 21,08±5,19 мМоль/л.
В ходе исследования показано, что концентрации ионов Na+, K+, Mg2+ и NH4+ оставались неизменными в течение всего эксперимента, в то время как концентрация ионов Ca2+ повысилась уже к 3 часам экспозиции в условиях гипоксии до 35,07±6,52 мМоль/г и сохранялась на повышенном уровне после 3х часовой нормоксии от суммарной 6 часовой гипоксии (рис. 3).
По-видимому, индуцированное гипоксией среды обитания повышение концентрации Ca2+ в мантийной жидкости связано с выходом ионов в межклеточную среду из-за недостатка энергии, снижения интенсивности активного транспорта ионов и вымывания Ca2+ из раковины. Ранее проведенными исследованиями Мартемьянова В.И. (2011) на примере D. polymorpha показано, что повышенная концентрация Ca2+ в гемолимфе пресноводных двустворчатых моллюсков является защитной реакцией в ответ на разные по качеству экстремальные воздействия (температура, токсиканты) (Martem'yanov, 2011), что обеспечивает поддержание pH гемолимфы.
Таким образом, кратковременная индуцированная гипоксия приводит к активации физиологических механизмов стресс-адаптации у эврибионтного вида большого прудовика и не приводит к активации биохимических и молекулярных механизмов стресс-защиты.
В экспериментальных условиях при принудительном кожном дыхании (через покровы тела и заполненное водой легкое) выживаемость большого прудовика при температуре, близкой к 20оС, не превышает 44-50 ч (Stadnichenko et al., 1996), что указывает на ограниченность физиолого-биохимических механизмов адаптации большого прудовика к дефициту растворенного кислорода в воде. Поэтому в теплый период года атмосферное дыхание у данного вида является преобладающим. В зимний период, при температурах, близких к 0оС, потребности в кислороде у большого прудовика, как пойкилотермного организма, резко снижаются, что позволяет ему существовать исключительно за счет растворенного в воде кислорода.
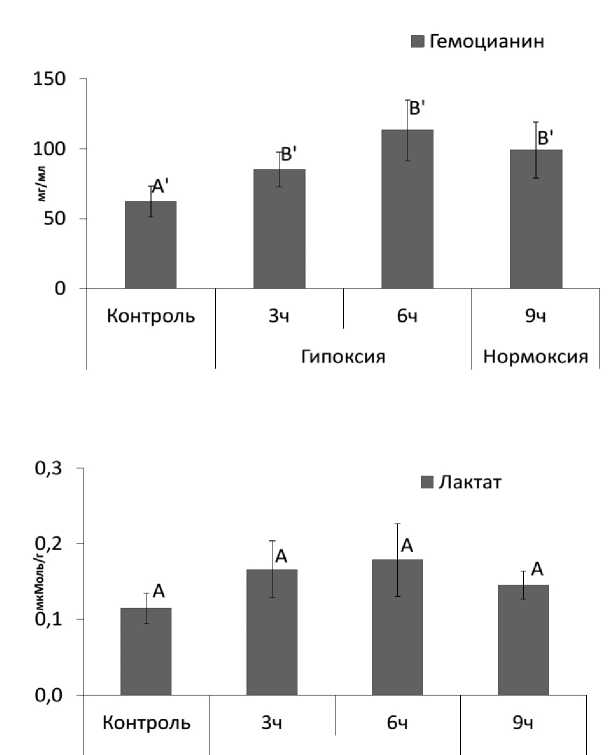
Рисунок 1. Изменение содержания гемоцианина (1-А) и лактата (1-Б), в мантийной жидкости L. stagnalis при экспозиции в условиях гипоксии и нормоксии.
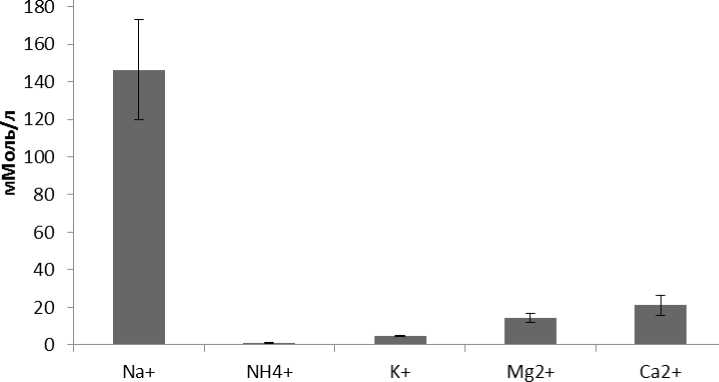
Рисунок 2. Содержание катионов в мантийной жидкости L. stagnalis в контрольных условиях.
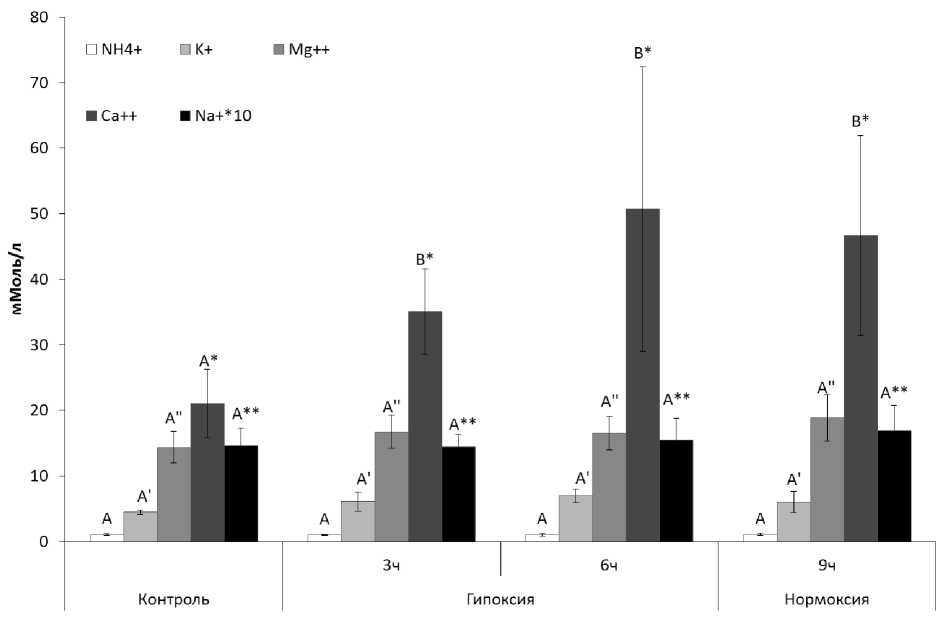
Рисунок 3. Изменение содержания катионов в мантийной жидкости L. stagnalis при экспозиции в условиях гипоксии и нормоксии
ACKNOWLEDGEMENT
Работа выполнена при частичной финансовой поддержке грантов РФФИ № 14-04-00501_а (приобретение расходных материалов), 14-3450051 (стажировка А.С.Хомича), РНФ 14-14-00400 (приобретение расходных материалов), базовой (1354) и проектной (6.382.2014) частей государственного задания, программы стратегического развития ФГБОУ ВПО «ИГУ» (приобретение расходных материалов), гранта молодых ученых ФГБОУ ВПО «ИГУ» и совместной программы академических обменов DAAD -Минобрнауки РФ М. Ломоносов 2014-2015.
References The impact of experimental hypoxia and subsequent normoxia on the content of some ions and markers of physiological stress-adaptation in gastropod species Lymnaea stagnalis
- Axenov-Gribanov D.V. (2013) Comparative evaluation of the impact of changes in the ambient temperature at the Baikal and Palaearctic organisms with focus on energy metabolism and nonspecific stress response: Dis. thesis. ISU, Irkutsk, 169 p
- Coutellec M.A., Lagadic L. (2006) Effects of self-fertilization, environmental stress and exposure to xenobiotics on fitness-related traits of the freshwater snail Lymnaea stagnalis. Ecotoxicology. 15(2) 199-213
- Ekkekakis P., Hall E.E., Petruzzello S.J. (2004). Practical markers of the transition from aerobic to anaerobic metabolism during exercise: rationale and a case for affect-based exercise prescription. Preventive medicine. 38(2) 149-159
- Frederich M. et al. (2000) Haemolymph Mg (2+) regulation in decapod crustaceans: physiological correlates and ecological consequences in polar areas. Journal of experimental biology. 203(8) 1383-1393
- Gnatishina L.L., Fal’fushinskaya G.I., Golubev O.P., Dallinger R., Stoliar O.B. (2011) Role of metallothioneins in adaptation of Lymnaea stagnalis (Mollusca: Pulmonata) to environment pollution. Hydrobiological Journal. 47(5) 56 -66
- Grieshaber M.K., Hardewig I., Kreutzer U. (1994) Physiological and metabolic responces to hypoxia in invertebrates. Rev. physiol. Biochem. Pharmocol, 125. 44 -147
- Konev O.J., Afonin V., Dromashko S. (2006) Age-related changes in the cellular composition of the mantle and the liquid spectrum cytomorphological signs of genetic damage gemotsitov Lymnaea stagnalis. Cytology and Genetics. 40(6) 21-27
- Konstantinov, A.S. (1986). General hydrobiology. MM: VSh. 4 ed. 483 p
- Lorenzon S. et al. (2008) Stress effect of two different transport systems on the physiological profiles of the crab Cancer pagurus. Aquaculture. 278(1) 156-163
- Martem'yanov V.I. (2011) Effect of the mineral composition of the external environment on the performance of water-salt metabolism instill in Rybinsk reservoir dreysseny Dreissena polymorpha Pallas. Russian journal. biol. Invasions, 2. 120-134
- Morris S., Taylor A. C., Bridges C. R. (1988). Response of haemolymph oxygen affinity to simultaneous salinity and oxygen stress in the intertidal prawn Palaemonelegans (Rathke). Comparative Biochemistry and Physiology Part A: Physiology. 90(1) 31-39
- Stadnichenko A.P., Slastenko N.N., Gouzenko O.V., Svitelsky N.N., Sychevsky A.S. (1996) The effect of trematode infestations and exposure to nitrate lead to lung and skin breathing Lymnaea stagnalis (Mollusca: Lymnaeidae). Parazitologiya. 30(1) 76 -80
- Stadnichenko A.P., Ivanenko L.D. et al. (1999) The influence of trematode invasion on soderzhaniegemotsianina in the hemolymph of the pond snail (Gastropoda, Pulmonata, Lymnaeidae). Parasitology. 33(2). 125 -128
- Timofeev MA (2010) Ecological and physiological aspects of adaptation to environmental factors of endemic Baikal and palearctic amphipods: Dis. thesis., TSU, Tomsk, 384 p
- Wijsman T. et al. (1988) Hyperglycaemia in relation to anoxia and stress in the freshwater snail Lymnaea stagnalis. Comparative Biochemistry and Physiology Part B: Comparative Biochemistry. 89(1), 55-63
